
Формирование пыльцевых зерен и зародышевого мешка кратко
Обновлено: 05.07.2024
Timeweb - компания, которая размещает проекты клиентов в Интернете, регистрирует адреса сайтов и предоставляет аренду виртуальных и физических серверов. Разместите свой сайт в Сети - расскажите миру о себе!
Виртуальный хостинг
Быстрая загрузка вашего сайта, бесплатное доменное имя, SSL-сертификат и почта. Первоклассная круглосуточная поддержка.
Производительность и масштабируемые ресурсы для вашего проекта. Персональный сервер по цене виртуального хостинга.
Выделенные серверы
Быстрая загрузка вашего сайта, бесплатное доменное имя, SSL-сертификат и почта. Первоклассная круглосуточная поддержка.
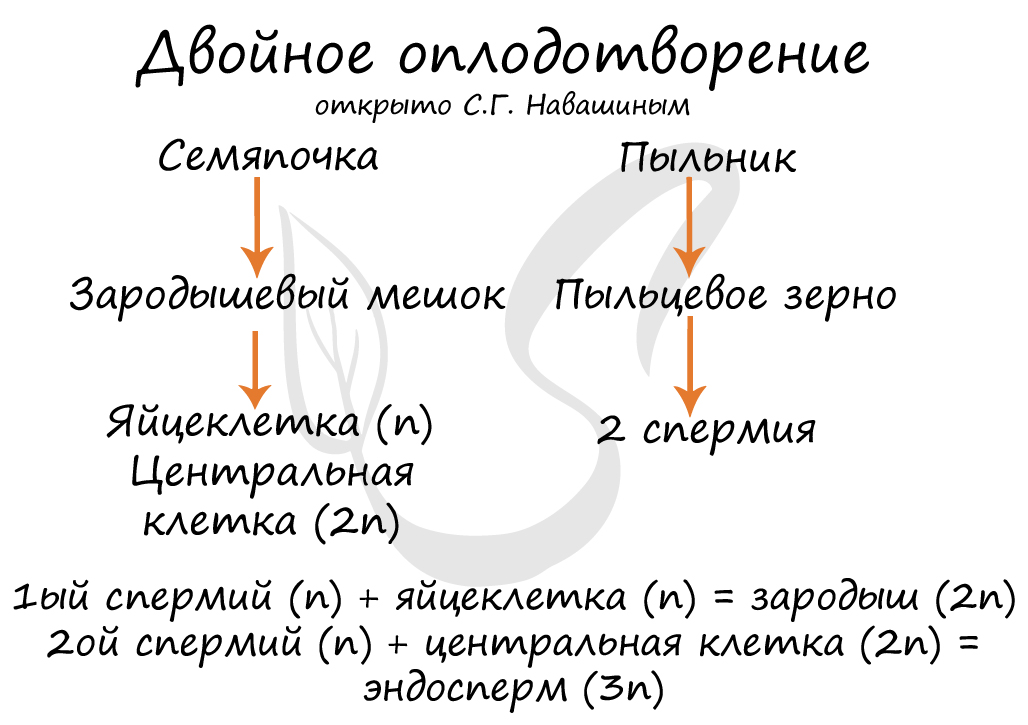
Суть двойного оплодотворения у цветковых растений заключается в том, что в нём участвуют два спермия. Один из них оплодотворяет яйцеклетку, и образуется зигота. Второй спермий сливается с центральной клеткой, из которой развивается запасающая ткань (эндосперм).

Мужской гаметофит (пыльцевое зерно) образуется в пыльцевых камерах пыльников тычинки из микроспоры. Пыльцевое зерно состоит из двух гаплоидных клеток: вегетативной и генеративной, покрытых оболочкой.
Образование женского гаметофита (зародышевого мешка) происходит в завязи пестика в семязачатке из мегаспоры. В состав зародышевого мешка входит семь клеток: гаплоидная яйцеклетка, центральная диплоидная клетка и пять вспомогательных гаплоидных клеток.
При попадании пыльцевого зерна на рыльце пестика начинается деление вегетативной клетки и образуется пыльцевая трубка. Пыльцевая трубка прорастает через столбик пестика и проникает в семязачаток через пыльцевход.
Генеративная клетка пыльцевого зерна делится и образует два спермия. По пыльцевой трубке спермии проникают в семязачаток. Один спермий сливается с яйцеклеткой и образует диплоидную зиготу. Второй спермий сливается с центральной клеткой и образует триплоидную клетку.
Зигота делится, и развивается в зародыш нового растения. Из триплоидной клетки формируется эндосперм. Стенки семязачатка становятся семенной кожурой. Таким образом, семязачаток становится семенем.
Отдел покрытосеменные (цветковые) самый многочисленный, он включает 235-250 тысяч видов. Его представители обитают по всему миру: от холодной тундры до жарких тропиков, отдельные виды освоили пресные и морские водоемы.

Покрытосеменные составляют большую часть массы растительного сообщества, являются звеном в цепи питания (продуцентами) - важнейшими производителями органических веществ на суше, как водоросли - в морях и океанах.
Цветок - генеративный орган покрытосеменных (цветковых), высшая ступень полового размножения. Цветок характерен только для покрытосеменных растений, ни один из других отделов подобным генеративным органом не обладает. По своему строению цветок это видоизмененный обоеполый стробил, гомологичный стробилам голосеменных.
В отличие от голосеменных, у которых семязачатки лежат открыто на семенных чешуях, у цветковых семязачаток находится в замкнутом вместилище - завязи, сформированной из плодолистика (-ов).
Двойное оплодотворение, открытое Навашиным Сергеем Гавриловичем, уникальное явление, характерное только для цветковых. Оно связано с тем, что в зародышевый мешок попадают два спермия, один из которых (n) сливается с центральной клеткой (2n), с образованием запасного питательного вещества - эндосперма (3n). Другой спермий (n) сливается с яйцеклеткой (n) с образованием зиготы (2n), из которой развивается зародыш.
У цветковых появляется плод - генеративный орган, служащий для защиты и распространения семян.
Ксилема - проводящая ткань, обеспечивающая восходящий ток воды и растворенных в ней минеральных солей, представлена не трахеидами, а сосудами. Во флоэме ситовидные элементы окружены клетками-спутницами.
У покрытосеменных мы не найдем антеридиев и архегониев: гаметофиты максимально редуцированы.
В процессе опыления покрытосеменных участвуют насекомые, летучие мыши, птицы. Также опыление может происходить с помощью воды или ветра.

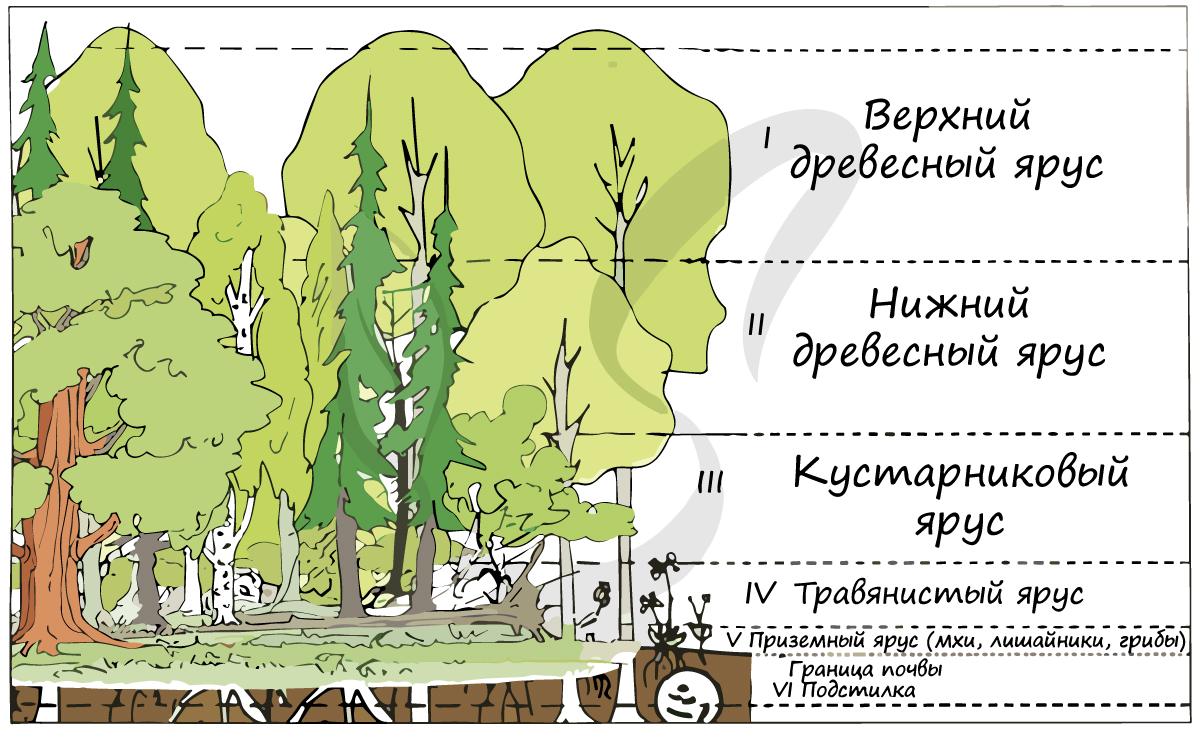
Особенностью цветковых является способность образовывать многоярусные сообщества, более устойчивые и продуктивные.
Многоярусность растительного сообщества служит приспособлением к равномерному распределению света: светолюбивые растения занимают верхний ярус, а теневыносливые растения отлично чувствуют себя в тени светолюбивых :)
Классы покрытосеменных
Отдел покрытосеменные состоит из двух классов: однодольные и двудольные. К классу двудольных относятся семейства: крестоцветные, сложноцветные, розоцветные, бобовые (мотыльковые), пасленовые. Класс однодольные включает в себя семейства: злаковые, лилейные. Для каждого класса имеются характерные признаки.
-
Двудольные - семейства: крестоцветные, сложноцветные, бобовые, розоцветные, пасленовые
-
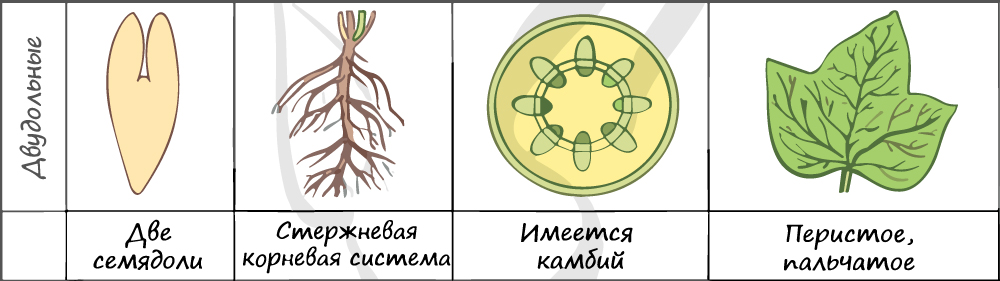
В составе зародыша обычно имеется две семядоли
В семядолях содержится запас питательных веществ. При надземном прорастании семядоли (зародышевые листья) могут выполнять функцию фотосинтеза.

Листья двудольных простые и сложные, для двудольных характерно перистое и пальчатое жилкование.
За счет камбия растения растут в толщину, возможен вторичный рост осевых органов (стебля и корня).
Корневая система чаще всего стержневого типа, с хорошо выраженным главным корнем, от которого отходят боковые корни. Главный корень развивается из зародышевого корешка.
Цветки пятичленные, реже встречаются четырехчленные. Хорошо обособлены чашечка и венчик.

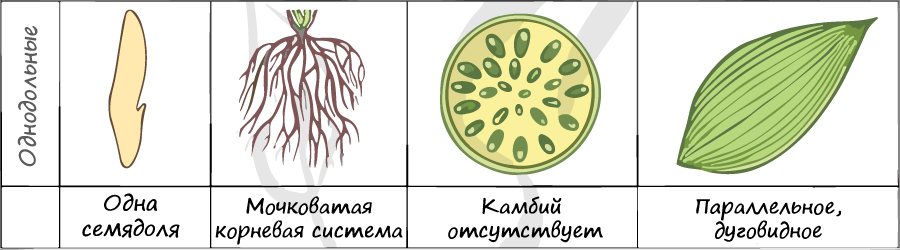
Цветок с простым околоцветником. Цветки чаще трехчленные, четырехчленные. Никогда не бывают пятичленными.
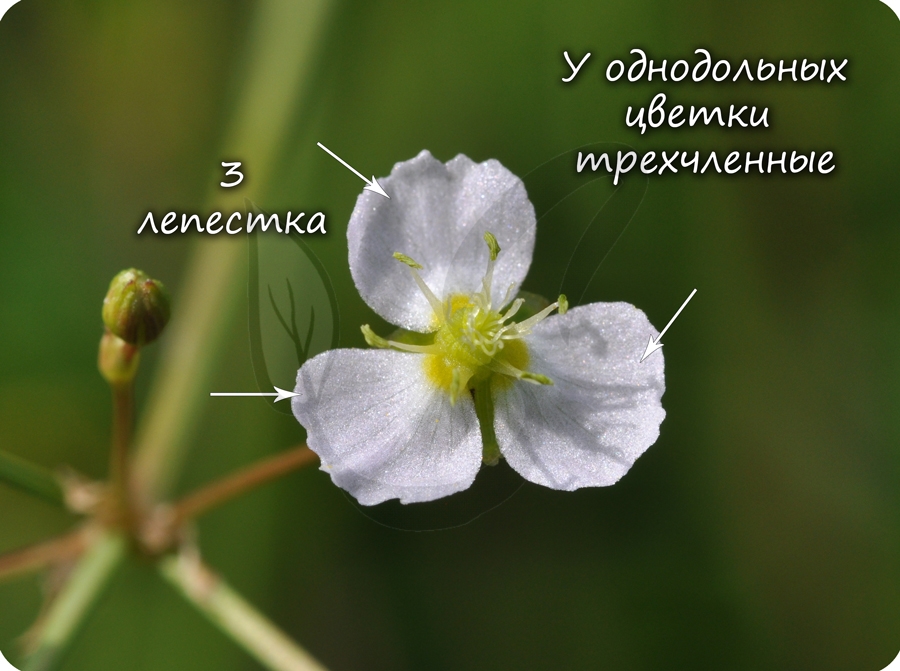
Эндосперм семени
Эндосперм (от греч. endon - внутри + греч. sperma - семя) - запасное питательное вещество, у покрытосеменных триплоидный (3n).
Эндосперм в семени есть у подавляющего большинства однодольных (лука, ландыша, пшеницы) и двудольных (тмина, хурмы, фиалки). Отсутствует эндосперм в семенах тыквенных, крестоцветных (капусты), сложноцветных (подсолнечника), бобовых (гороха, фасоли), также у - березы, липы, дуба, клена, так как на ранней стадии развития растущий зародыш поглощает эндосперм.
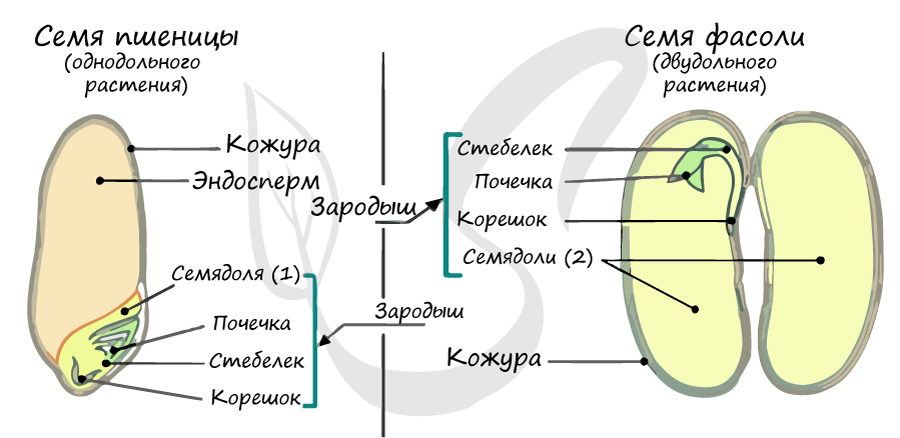
Жизненный цикл
Из генеративных почек спорофита развиваются цветки. У взрослого растения спорофита (2n) в цветке в гнездах пыльников тычинок в ходе микроспорогенеза образуется пыльцевое зерно (n) - мужской гаметофит. В завязи пестика в семязачатке формируется женский гаметофит - зародышевый мешок, внутри которого находятся центральная клетка (2n) и яйцеклетка (n).
В результате опыления (насекомым, ветром, человеком) пыльца с тычинок переносится на рыльце пестика. Пыльцевое зерно состоит из вегетативной и генеративной клеток. Вегетативная клетка начинает растворять ткани пестика, образует пыльцевую трубку и прорастает до зародышевого мешка. Генеративная клетка делится, образуя два спермия (n), из которых один сливается с центральной клеткой (2n) с образование эндосперма (3n) - запасного питательного вещества. Другой спермий (n) сливается с яйцеклеткой (n), образуя зиготу (2n).
В дальнейшем из семязачатка формируется семя, а завязь превращается в околоплодник - образуется плод. Своим внешним видом плоды привлекают животных, и те их охотно поедают) Благодаря семенной кожуре семена не подвергаются расщеплению в желудочно-кишечном тракте человека и животных. Они выходят из ЖКТ в неизменном виде и остаются способны к прорастанию: так происходит расселение растений. Попав в благоприятные условия, они прорастают в спорофит (2n). Цикл замыкается.
Значение покрытосеменных
Покрытосеменным в жизни человека отведено важное место. Только подумайте - почти все культурные растения принадлежат к этому отделу! Цветковые имеют медицинское значение, из многих растений изготавливаются лекарства. Их древесина используется для изготовления бумаги, мебели, применяются в промышленности.

Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Органом полового размножения покрытосеменных растений является цветок. Цветок — видоизмененный, укороченный, неразветвленный побег, предназначенный для образования спор и гамет и полового процесса, завершающегося образованием семян и плода.
Строение цветка
У цветка различают цветоножку, цветоложе, околоцветник, тычинки и пестики. У некоторых цветков отдельные части могут отсутствовать.
Цветки большинства видов растений имеют и тычинки, и пестики. Такие цветки называют обоеполыми (вишня, горох). Цветки, которые имеют только пестики, называют пестичными (женскими). Цветки, которые имеют только тычинки, называют тычиночными (мужскими). В зависимости от распределения однополых цветков на растениях различают: однодомные растения — растения, у которых на одних и тех же экземплярах располагаются и женские, и мужские цветки (огурец, кукуруза, дуб); двудомные растения — растения, у которых на одних экземплярах располагаются женские, а на других — мужские цветки (крапива двудомная, конопля, облепиха); многодомные растения — растения, у которых на одних и тех же экземплярах встречаются как обоеполые, так и однополые цветки в различных количественных соотношениях (гречиха, некоторые виды ясеня, клена).
Цветоножка — междоузлие под цветком. Цветки, лишенные цветоножки, называются сидячими (цветки в соцветии корзинка у подсолнечника, астры, одуванчика).
Цветоложе — укороченная стеблевая часть цветка. На ней располагаются все остальные части цветка.
Околоцветник — стерильная часть цветка, его покров. Околоцветник может быть простым (не дифференцированным на чашечку и венчик, образованным совокупностью однородных листочков, имеющих одинаковые размеры и окраску) и двойным (дифференцированным на чашечку и венчик, отличающиеся друг от друга размерами и окраской. Простой околоцветник может быть венчиковидным (образованным ярко окрашенными листочками) или чашечковидным (образованным зелеными листочками). Цветки, лишенные околоцветника (ива, тополь), называются голыми.
Чашечка — наружная часть двойного околоцветника, представляет собой совокупность чашелистиков — видоизмененных прицветных листьев. Обычно чашелистики имеют небольшие размеры и зеленую окраску. Они сходны с обычными листьями, но устроены проще.
Различают: раздельнолистную чашечку — чашечку, образованную свободными (несросшимися) чашелистиками (капуста, лютик); сростнолистную чашечку — чашечку, образованную частично или полностью сросшимися чашелистиками (картофель, табак, горох).
Венчик — внутренняя, обычно окрашенная часть двойного околоцветника. Представляет собой совокупность лепестков, часто имеющих яркую окраску. Количество лепестков венчика может быть различным. Лепестки могут быть более или менее одинаковыми (лютик,яблоня) либо отличаться размерами и формой (фиалка, горох). В результате венчик может быть правильным, неправильным или асимметричным. Венчик, как и чашечка, может быть раздельнолепестным и сростнолепестным. Раздельнолепестной венчик состоит из свободных, несросшихся лепестков. Сростнолепестной венчик состоит из сросшихся в той или иной степени лепестков. Главная функция венчика — привлечение опылителей.
Андроцей
Андроцей — совокупность тычинок одного цветка. Количество тычинок в цветке — от одной (орхидные) до нескольких сотен (некоторые кактусы). У большинства растений тычинка состоит из тычиночной нити и пыльника. Тычиночная нить — нижняя, как правило, суженная стерильная часть тычинки. Нижний конец тычиночной нити отходит от цветоложа, а верхний конец несет пыльник. Обычно тычиночные нити тонкие, длинные, округлые в сечении. Пыльник — верхняя расширенная фертильная часть тычинки. Пыльник состоит из двух половинок, соединенных связником. Каждая половинка имеет два пыльцевых гнезда (микроспорангия), в которых происходит образование микроспор, а впоследствии пылинок. Связник является продолжением тычиночной нити, через него в пыльник поступают питательные вещества.
Микроспорогенез
Микроспорогенез — процесс образования микроспор в микроспорангиях (гнездах пыльника). Микроспоры формируются из материнских клеток — микроспороцитов, имеющих диплоидный набор хромосом. В результате мейоза каждая материнская клетка образует четыре гаплоидных микроспоры. Микроспоры быстро обособляются друг от друга.
Микрогаметогенез
Микрогаметогенез — процесс образования мужских половых клеток (спермиев), происходит в пыльцевом зерне, которое является мужским гаметофитом покрытосеменных растений. Развитие мужского гаметофита происходит также в гнездах пыльников тычинок и сводится к одному митотическому делению микроспоры и формированию оболочек пыльцевого зерна. Оболочка пыльцевого зерна состоит из двух слоев: интины (внутренней, тонкой) и экзины (наружной, толстой). Каждое пыльцевое зерно содержит две гаплоидные клетки: вегетативную и генеративную. Из генеративной (спермагенной) далее образуются два спермия. Из вегетативной (сифоногенной) впоследствии образуется пыльцевая трубка.
Гинецей
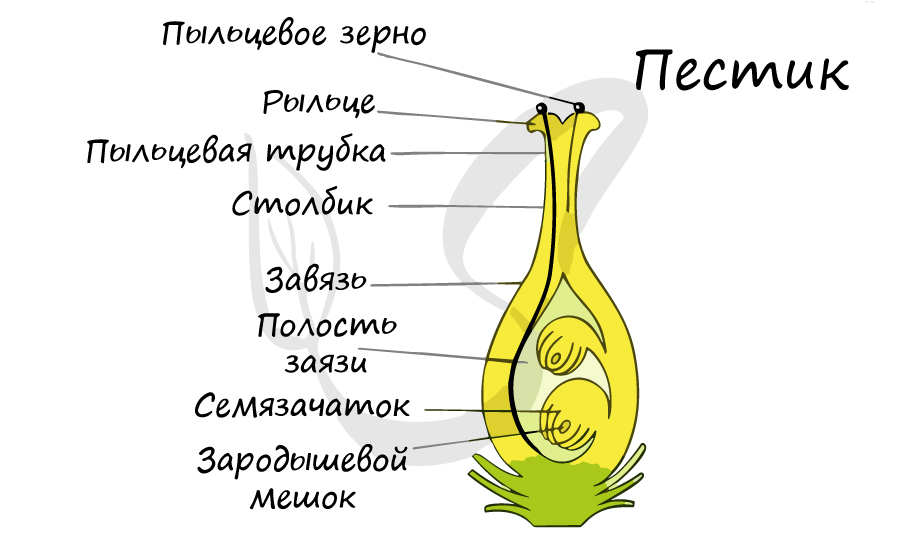
Гинецей — совокупность пестиков одного цветка. Обычно в пестике выделяют три части: завязь, столбик и рыльце.
Завязь — замкнутая, нижняя, полая часть пестика, несущая и защищающая семязачатки. Завязь бывает: верхняя, нижняя, полунижняя. В завязи может располагаться от одного (пшеница, вишня) до нескольких тысяч (мак) семязачатков. Стенки завязи выполняют функцию защиты семязачатков от неблагоприятных факторов среды (высыхание, колебание температур, поедание насекомыми и т.д.). Внутри завязи (в семязачатках) происходит мегаспорогенез и мегагаметогенез, они принимают участие в образовании околоплодника.
Столбик — средняя более или менее удлиненная стерильная часть пестика, отходящая обычно от верхушки завязи, соединяет завязь и рыльце.
Рыльце — верхняя расширенная часть пестика, предназначено для восприятия пыльцы. Рыльце может быть разнообразной формы (двухлопастное, звездчатое, перистое и т.д.) и размера в зависимости от особенностей опыления. При отсутствии столбика рыльце называют сидячим.


Семязачаток состоит из нуцеллуса (ядра) — центральной части, являющейся мегаспорангием, двух покровов — интегументов, которые при смыкании образуют узкий канал — микропиле, или пыльцевход, через который пыльцевая трубка проникает к зародышевому мешку. С помощью семяножки семязачаток прикрепляется к плаценте. Место прикрепления семязачатка к семяножке называют рубчиком. Противоположную микропиле часть семязачатка, где сливаются нуцеллус и интегументы, называют халазой.
В семязачатке происходят мегаспорогенез, мегагаметогенез и процесс оплодотворения. После оплодотворения (реже без него) из семязачатка формируется семя.

Мегаспорогенез
Процесс формирования мегаспор называется мегаспорогенезом. Он происходит в нуцеллусе семязачатка. После заложения семязачатка и формирования нуцеллуса в области микропиле начинает разрастаться одна археспориальная (спорогенная) клетка — мегаспороцит, или материнская клетка мегаспор.
Материнская клетка мегаспор имеет диплоидный набор хромосом. У большинства покрытосеменных из нее путем мейоза образуются четыре гаплоидные мегаспоры. Из них лишь одна (обычно нижняя, обращенная к халазе, реже верхняя, обращенная к микропиле) дает начало женскому гаметофиту — зародышевому мешку. Остальные мегаспоры отмирают.
Мегагаметогенез
Процесс формирования женских половых клеток происходит в зародышевом мешке. Формирование женского гаметофита начинается с разрастания мегаспоры, которая далее три раза делится митозом. В результате этого образуются восемь клеток, которые располагаются следующим образом: три — на одном полюсе зародышевого мешка (микропилярном), три — на другом (хадазальном), две — в центре. Две оставшиеся сливаются в центре клетки, образуя диплоидную центральную клетку зародышевого мешка. Одна из трех клеток, расположенных на микропилярном полюсе, отличается большими размерами и является яйцеклеткой. Две рядом расположенные клетки являются вспомогательными и называются синергидами. Группа из трех клеток, находящихся на противоположном, халазальном полюсе, называется антиподом. Таким образом, сформированный женский гаметофит включает шесть гаплоидных клеток (яйцеклетка, две клетки-синергиды, три клетки-антипода) и одну диплоидную клетку.
Оплодотворение. Образование семян и плодов
Купить проверочные работы
и тесты по биологии




Процессу оплодотворения предшествует опыление — перенос пыльцы от пыльцевых мешков тычинок к рыльцам пестиков. Попав на рыльце пестика, под воздействием веществ, выделяемых пестиком, пыльца начинает прорастать: образуется пыльцевая трубка, внедряющаяся в ткань рыльца. Кончик пыльцевой трубки выделяет вещества, размягчающие ткань рыльца и столбика. В процессе формирования пыльцевой трубки принимает участие сифоногенная клетка. По мере роста пыльцевой трубки в нее переходит спермагенная клетка, которая делится митозом с образованием двух спермиев (у некоторых растений спермагенная клетка дает начало двум спермиям еще до прорастания пыльцы). Пыльцевая трубка продвигается по столбику пестика и врастает в зародышевый мешок, как правило, через микропиле. После проникновения в зародышевый мешок кончик пыльцевой трубки разрывается, и спермии попадают внутрь. Один из спермиев сливается с яйцеклеткой, образуя диплоидную зиготу. Второй спермий сливается с центральной клеткой зародышевого мешка, образуя триплоидную клетку, из которой далее формируется эндосперм (питательная ткань) семени, обеспечивающий питание зародыша. Синергиды и антиподы дегенерируют. Вышеописанный процесс получил название двойного оплодотворения. Двойное оплодотворение у цветковых растений было открыто в 1898 году русским ботаником С.Г. Навашиным.
После двойного оплодотворения из яйцеклетки формируется зародыш семени, из центрального ядра зародышевого мешка — эндосперм, из интегументов — семенная кожура, из всего семязачатка — семя, а из стенок завязи — околоплодник. В целом из завязи пестика формируется плод с семенами.
Процесс формирования половых клеток у растений подразделяется на два этапа: 1-й этап — спорогенез — завершается образованием гаплоидных клеток — спор, в ходе 2-го этапа — гаметогенеза — происходит ряд делений гаплоидных клеток, прежде чем образуются зрелые гаметы.
Процесс образования микроспор, или пыльцевых зерен, у растений называют микроспорогенезом, а процесс образования мегаспор (или макроспор) — мега- или макроспорогенезом. Микроспорогенез протекает аналогично делению созревания у животных мужских половых клеток до стадии сперматиды, а мегаспорогенез — соответственно до стадии незрелой яйцеклетки — ооцита II.
Процесс гаметогенеза у растений в принципе сходен с таковым у животных, но протекает несколько отличным путем. У животных после двух мейотических делений формируются гаметы, и никаких дополнительных клеточных делений не происходит. У растений в результате двух мейотических делений возникает гаплоидная спора, из которой развивается гаметофит, представляющий собой у низших растений (грибов, печеночников, мхов, ряда водорослей) целый организм и наиболее продолжительную стадию цикла существования. У высших растений гаплоидная фаза редуцирована, однако ядра мужской и женской спор претерпевают ряд митотических делений, прежде чем образуются гаметы.
Микроспорогенез и микрогаметогенез
Мы рассмотрим микроспорогенез и микрогаметогенез на примере покрытосеменных растений как наиболее общем. В субэпидермальной ткани молодого пыльника обособляется специальная спорогенная ткань, называемая археспорием. Каждая первичная археспориальная клетка после ряда делении становится материнской клеткой пыльцы (микроспороцитом), которая проходит все фазы мейоза.
Схема микроспорогенеза (1-6) и микрогаметогенеза (5-10) у растений
В результате двух мейотических делений возникают четыре гаплоидные микроспоры. Последние лежат четвёрками и называются клеточными тетрадами.
У однодольных растений каждое деление ядра в мейозе, как правило, сопровождается цитокинезом; у двудольных оба деления клетки наступают одновременно по окончании мейоза.
При созревании клеточные тетрады распадаются на отдельные микроспоры с образованием внутренней (интина) и наружной (экзина) оболочек. Наружная оболочка, как правило, грубая, кутинизированная, поверхность ее либо гладкая, либо шероховатая; приспособленная для переноса пыльцы и прилипания ее к рыльцу пестика. Этим заканчивается микроспорогенез вслед за образованием одноядерной микроспоры начинается микрогаметогенез. Первое митотическое деление микроспоры приводит к образованию вегетативной и генеративной клеток. В дальнейшем вегетативная клетка и ее ядро не делятся. В ней накапливаются запасные питательные вещества, который в последующем обеспечивают деление генеративной клетки и рост пыльцевой трубки в столбике пестика.
Генеративная клетка, содержащая меньшее количество цитоплазмы, вновь делится. Это деление может осуществляться еще в пыльцевом зерне или в процессе его прорастания в пыльцевой трубке. В результате образуются две мужские половые клетки, которые в отличие от сперматозоидов животных называются спермиоклетками, или спермиями.
Таким образом, из одной споры с гаплоидным набором хромосом в результате двух митотических делений образуются три ядра: Два из них — спермии и одно — вегетативное. При образовании пыльцевой трубки это вегетативное ядро в полужидком диффузном состоянии переходит в пыльцевую трубку.
Процесс деления генеративной клетки и образование спермиев в пыльцевой трубке были впервые подробно изучены С. Г. Навашиным в 1910 г. на лилейных растениях.
Мегаспорогенез и мегагаметогенез
У покрытосеменных растений женский гаметофит — это зародышевый мешок, который закладывается и развивается внутри семяпочки.
Схема мегаспорогенеза (1-5) и мегагаметогенеза (6-12) у растений
Развитию женского гаметофита у высших покрытосеменных растений предшествует мегаспорогенез. В субэпидермальном слое молодой семяпочки обособляется археспориальная клетка, чаще она только одна. Клетка археспория растет, превращаясь в материнскую клетку мегаспоры. В результате двух делений мейоза материнской клетки мегаспоры образуется тетрада мегаспор. Каждая из клеток тетрады по числу хромосом является гаплоидной. Однако только одна из них продолжает развиваться, остальные три дегенерируют (моноспорический тип развития), судьба этих клеток напоминает судьбу редукционных телец при созревании яйцеклеток у животных.
На следующем этапе осуществляется мегагаметогенез. Оставшаяся функционировать мегаспора продолжает расти и затем ее ядро претерпевает ряд эквационных делений. При этом сама клетка не делится, делится только ядро.
У разных систематических групп растений число эквационных делений ядра мегаспоры может варьировать от одного до трех. У большинства растений (70% видов покрытосеменных) этих делений, как правило, в результате возникает восемь наследственно одинаковых ядер, вовремя этих делений ядра занимают полярное положение, четыре из них оказываются лежащими ближе к микропиле (место проникновения спермиев), а четыре других — в противоположном конце зародышевого мешка, называемого халазальным. Дальше эти ядра обособляются в самостоятельные клетки, имеющие значительные количества цитоплазмы.
Из четырех клеток, располагающихся у микропиле, три клетки — яйцеклетка, и две так называемые синергиды образуют яйцевой аппарат. Однако из этих трех клеток после оплодотворения развивается только одна, а две другие разрушаются. Четвертое ядро отходит к центру зародышевого мешка, где сливается с одним из ядер, отошедшим от халазального конца. Слившиеся в центральной части два гаплоидных ядра образуют одно диплоидное — вторичное или центральное, ядро зародышевого мешка. Это ядро с цитоплазмой зародышевого мешка называют обычно центральной клеткой зародышевого мешка. Однако часто полярные ядра, передвинувшиеся к центру, не сливаются до оплодотворения. Оставшиеся у халазального конца зародышевого мешка три ядра также обособляются в клетки; они называются антиподами.
Таким образом, в результате трех митотических делений в зародышевом мешке образуется 8 наследственно одинаковых гаплоидных ядер, из которых только одно дает яйцеклетку.
Рассмотренная схема образования восьмиядерного зародышевого мешка из одной мегаспоры является наиболее типичной. Однако у различных групп растений этот процесс протекает весьма разнообразно. В одних случаях, как мы только что рассмотрели, развитие зародышевого мешка начинается из одной гаплоидной споры (моноспорический тип развития), в других — из двух (биспорический тип) и четырех спор (тетраспорический тип).
Типы развития зародышевых мешков
Как мы указывали, при моноспорическом типе развивается лишь одна мегаспора из четырех, а остальные три разрушаются подобно тому, что имеет место с редукционными тельцами у животных. При других типах развития зародышевого мешка сохраняется разное количество мегаспор, возникших в результате мейоза и готовых к дальнейшим митотическим делениям.
Образование пыльцевых зерен и зародышевых мешков у цветковых растений
Изучая гаметогенез, нельзя не поражаться тому параллелизму, который наблюдается при созревании половых клеток у животных и растений, несмотря на то, что их расхождение (дивергенция) в филогенезе произошло на очень раннем этапе возникновения клеточной организации. Это указывает на однотипность принципов построения ряда приспособительных механизмов как в растительном, так и животном мире.
Итак, изучение развития половых клеток у животных и у растений показало, что формирование гамет является сложным процессом. Прежде чем яйцеклетка и спермий объединятся в процессе оплодотворения, они претерпевают ряд превращений. Однако половые клетки так же, как и клетки любой другой ткани, происходят из соматических. Поэтому их нельзя рассматривать как нечто обособленное от тела организма. Вместе с тем половые клетки имеют и свои особенности. Основными характерными моментами, отличающими их от соматических клеток, являются следующие:
1. У разных животных и растений на разных стадиях дифференциации тканей зародыша происходит обособление половых клеток. Процесс закладки и дифференциации, половых клеток у животных называется зачатковым путем.
2. В процессе развития половых клеток особое значение имеет мейоз с характерными для него стадиями деления ядра, а именно профазой I, во время которой конъюгируют гомологичные хромосомы, метафазой I и анафазой I, когда осуществляется редукция числа хромосом и расхождение гомологичных хромосом к различным полюсам.
3. Главным свойством половых клеток является способность их при оплодотворении сливаться в одну с образованием зиготы, которая претерпевает затем дробление и развитие. Соматические клетки этой способностью, как правило, не обладают.
Читайте также: