
Экологические основы эволюции кратко
Обновлено: 04.07.2024
Свидетельство и скидка на обучение каждому участнику
Зарегистрироваться 15–17 марта 2022 г.
Министерство образования и науки Российской Федерации
федеральное государственное бюджетное образовательное учреждение высшего профессионального образования
Институт Естествознания и Экономики
Кафедра общей биологии, экологии и методики обучения биологии
ПО ДИСЦИПЛИНЕ
ГЕНЕТИКО-ЭКОЛОГИЧЕСКИЕ ОСНОВЫ ЭВОЛЮЦИОННОГО ПРОЦЕССА
Уровни естественного отбора
Все организмы различаются между собой по тем или иным анатомо-физиологическим критериям (физическим возможностям, активности, обучаемости и т. д.). Это обстоятельство очень важно для эволюционного процесса, поскольку делает вид и популяции гетерогенными. Посредством различных способов борьбы за выживание проявляется реальный уровень приспособленности организма. Задачей естественного отбора - второй движущей силы эволюции - является удаление слабых и выживание самых сильных особей.
Отбор осуществляется на всех этапах индивидуального развития и даже до образования зиготы, потому что в размножении участвуют только особи, доказавшие свою жизнеспособность. Менее подготовленные особи либо погибают на ранних этапах, а если выживают, то не выдерживают конкуренции за полового партнера с более сильными представителями вида. Это не относится лишь к человеку. Отбор на ранних стадиях развития проявляется в избирательной смертности эмбрионов с отклонениями развития. При этом отбраковываются наименее удачные генотипы. То же самое происходит на всем протяжении дорепродуктивного периода особи. В итоге отбор обеспечивает избирательное размножение особей, обладающих наиболее ценными генотипами.
В природных условиях естественный отбор осуществляется не по генотипу, а только по фенотипу. Напомним, что в гетерозиготном состоянии генотип содержит два разных аллеля, один из которых (доминантный) выражается в фенотипе. Поэтому полезное изменение, находясь в рецессивном состоянии, не может воздействовать на отбор. Но возможен вторичный отбор генотипов, который происходит через отбор фенотипов, поскольку гетерозиготная особь продуцирует гаметы двух типов, один из которых будет содержать изменившийся аллель. Если значение измененного аллеля для повышения адаптивности организма велико, то число его носителей с каждым сезоном размножения будет расти. Таким образом, отбор наиболее эффективно действует по отношению к доминантным аллелям. Естественный отбор, который направлен на сохранение определенных фенотипов, называется положительным. Отбор, удаляющий из популяции фенотипы, называется отрицательныьм.
Естественный отбор характеризуется четырьмя основными особенностями: вероятностью, накапливающим и интегрирующим действием, адаптивной направленностью.
ЕСТЕСТВЕННЫЙ ОТБОР - результат борьбы за существование; он основывается на преимущественном выживании и оставлении потомства наиболее приспособленными особями каждого вида и гибели менее приспособленных организмов
В условиях постоянного изменения среды естественный отбор устраняет неприспособленные формы и сохраняет наследственные уклонения, совпадающие с направлением изменившихся условий существования. Происходит либо смена нормы реакции, либо ее расширение (нормой реакции называется способность организма реагировать приспособительными изменениями на действие факторов среды; норма реакции - это пределы модификационной изменчивости, контролируемой генотипом данного организма.
Уровни естественного отбора
Естественный отбор может быть не только на индивидуальном уровне, но и на других, если биологические единицы этого уровня соответствуют следующим критериям:
- способны производить избыток единиц, по сравнению с количеством, нужным для замещения родителей;
- выживаемость единиц коррелирует с их признаками;
- существуют механизмы, обеспечивающие передачу этих признаков потомству.
Таким образом, можно выделить следующие уровни естественного отбора:
Групповой отбор (популяционный отбор)
1. Гаметный отбор – этот отбор начинается собственно еще до онтогенеза и определяется отличиями в селективной ценности половых клеток. При этом отбору подвержены следующие признаки:
- жизнеспособность разных генотипов;
- способность соединяться с другой гаметой.
Отбору подвергаются все гены, а так как гаметы диплоидны, этот отбор очень эффективен. Все негативные мутации элиминируют, все положительные – быстро распространяются в популяции.
Например, у Drosophila melanogaster одна из хромосом гомологичной пары включается в более эффективно функционирующие сперматозоиды. Нарушенное расщепление хромосом в мейозе ведет к повышению частоты хромосомы одного типа в гаметном фонде по сравнению с частотой ее гомолога (мейотический драйв).
У растений пыльцевые зерна одного типа могут дать начало пыльцевым трубкам, которые растут быстрее, чем пыльцевые трубки других зерен, то пыльцевые зерна такого типа смогут оплодотворить большее число яйцеклеток. Поскольку гаметы гаплоидны, отбору подвергаются все их гены, и, таким образом, гаметный отбор очень эффективен. Следовательно, можно ожидать, что любая вредная мутация будет элиминироваться очень быстро, а любая благоприятная мутация так же быстро распространится. Если, однако, какой-либо аллель обладает плейотропным действием и, давая преимущество гамете, в то же время снижает приспособленность диплоидного организма, то частота такого аллеля будет поддерживаться на некотором промежуточном уровне.
2. Отбор родичей. Основные положения теории отбора родичей разработал У.Гамильтон (1964). Позже Мэйнард Смит (1964) ввел термин кин-отбор, определив его как отбор, направленный на сохранение признаков, благоприятствующих выживанию близких родичей данной особи. Такими родичами могут быть либо потомки данной особи, либо её сибсы и другие родичи. В качестве примеров признаков, возникших в результате кин-отбора, он привел заботу о потомстве, имитацию ранения, а также касты стерильных рабочих у общественных насекомых.
Мэйнард Смит и более позние исследователи считают кин-отбор процессом, близким к социально-групповому отбору, и, возможно, промежуточным между индивидуальным и групповым отбором.
Набор признаков — предупреждающая окраска у несъедобных насекомых — в течение некоторого времени считался результатом кин-отбора.
Этот отбор происходит в колониях, семьях и сверхмалых популяциях и приводит к накоплению альтруистических признаков.
Альтруизм – это особая форма социального поведения, которая выражается в склонности некоторых особей жертвовать индивидуальными интересами в пользу интересов других особей.
Альтруистическое поведение животных слагается из разнообразных специфических особенностей поведения. В общем и целом его можно определить как поведение, приносящее пользу другим особям. Его диапазон очень широк — от поведения, благоприятствующего другим особям в умеренной степени, до подлинного самопожертвования, вплоть до самоубийства. Альтруистическое поведение может принести пользу потомкам, другим родичам или социальным группам.
Существование различных типов альтруистического поведения позволяет считать, что в их создании могут участвовать разные типы отбора. Рассмотрим три случая.
Первый случай — альтруистическое поведение родительских особей по отношению к своим потомкам. К этому типу альтруистического поведения можно отнести общее явление заботы о потомстве у птиц и млекопитающих или особые виды поведения, характерные для чибисов, крикливых зуйков и других землегнездников, которые притворяются ранеными, отвлекая внимание хищника, или отводят его от гнезда. Забота о потомстве — явно результат индивидуального отбора, поскольку индивидуальный отбор благоприятствует сохранению генов тех родительских особей, которые оставляют наибольшее число выживающих потомков.
Рассмотрим теперь связанное с самопожертвованием оборонительное поведение рабочих особей у общественных пчел, таких, как Apis mellifera . Когда рабочая пчела использует жало, это равносильно для нее самоубийству, но полезно для колонии, так как предотвращает вторжение врага. Колонии, обороняющиеся лучше других, выживают и размножаются более успешно, чем колонии с менее эффективной защитой. Самопожертвование рабочих пчел, наряду с другими характеристиками касты рабочих, можно адекватно объяснить как результат социально-группового отбора, поскольку оно выгодно колонии пчел в целом.
Третий случай — группы примитивных людей, находящихся на стадии собирательства и охоты, примером которых могут служить бушмены юго-западной Африки (Lee, Devore, 1976*), Эти сообщества представляют собой организованные группы, в которые входят члены семьи, другие родственники, свойственники, а иногда случайные гости из других групп. В них глубоко укоренен обычай разделения пищи. Если убито крупное животное, его мясо раздается всем членам группы независимо от того, родственники это или случайные посетители. В таких группах развиваются также другие типы кооперативного поведения.
Допустим теперь в порядке обсуждения, что распределение корма и другие аналогичные типы социального поведения имеют какую-то генетическую основу; это позволит нам попытаться изучить типы отбора, которые, возможно, участвуют в развитии такого поведения.
Этот отбор определяется соотношением популяции и ресурса. Если в условиях ограниченных ресурсов в популяции преобладают особи, способные полностью исчерпать ресурсы – популяция может погибнуть. Если в популяции из-за соответствующих мутаций не возникли подобные фенотипы, способные исчерпать ресурс – популяция доживет до его возобновления и даже расселится.
Адаптивные признаки бесполых особей — касты рабочих у общественных насекомых — служат прекрасным примером социально-группового отбора в действии. Так, колония медоносной пчелы ( Apis mellifera ) представляет собой популяцию-сообщество, состоящую из самок (матки), самцов (трутней) и бесполых особей (рабочие). Матки и трутни фертильны, но бесполезны для экономики социальной группы.
Важные функции добывания пищи, защиты и поддержания колонии и выращивания молоди выполняют рабочие особи, обладающие соматическими и поведенческими адаптациями для выполнения этих функций. Носители главных адаптации, благодаря которым колония существует, — это рабочие особи. Между тем рабочие пчелы, будучи бесполыми, не размножаются как таковые, а поэтому не имеют возможности передавать гены, определяющие их адаптивные черты, следующим поколениям. Эту задачу выполняют матки и трутни. Следовательно, если бы матки и трутни не несли генов, которые обеспечивают развитие умелых и прилежных рабочих пчел, то улей не мог бы процветать, а оказался бы элиминированным в результате конкуренции с другими ульями, содержащими лучше адаптированных рабочих особей. Подобный процесс замещения одной линии пчелиных семей другой действительно наблюдался за последние годы в разных частях земного шара (см. Michener, 1975*). Основной единицей отбора в этом случае, определённо, служит колония.
Социально-групповой отбор у общественных перепончатокрылых облегчается характерным для этого отряда насекомых гаплодиплоидным способом определения пола, при котором самцы развиваются из неоплодотворенных яиц, а генетические самки —матки и рабочие особи — из оплодотворенных яиц. В результате матки и рабочие особи данной колонии, принадлежащие к одному и тому же поколению, приходятся друг другу сёстрами. Более того, в колонии перепончатокрылых родство между сёстрами теснее, чем между родительскими особями и потомками, в отличие от симметричных родственных отношений в пределах семьи у обычных раздельнополых видов. Ввиду такого особенно тесного родства между рабочими особями и матками у общественных перепончатокрылых, давление отбора на рабочих, принадлежащих к различным колониям, легко может перейти в отбор между их матками (Hamilton, 1964; 1972; Trivers, Hare, 1976*).
Общественные перепончатокрылые представляют собой один из примеров крайне высокой интегрированности сообществ в мире животных, и у них очень ярко выражены эффекты социально-группового отбора. Отбор этого типа, вероятно, действует также в группах с не столь высокоинтегрированной структурой сообщества, например у птиц и млекопитающих. В части VIII мы попытаемся показать, что некоторые особенности вида Homo sapiens , возможно, возникли в результате социально-группового отбора.
4. Половой отбор , форма естественного отбора, основанная на соперничестве особей одного пола (чаще мужского) за спаривание с особями другого пола. В результате полового отбора или при его участии у многих видов животных в процессе эволюции возникли и развились вторичные половые признаки (например рога у оленей).
Концепция полового отбора была выдвинута Ч. Дарвином в 1871 г. в связи с интерпретацией фактов полового диморфизма. Дарвин обнаружил, что существуют два пути, с помощью которых самец может получить преимущество перед другими самцами:
1.конкуренция между самцами в поединках или иных формах ритуального соперничества (внутриполовой отбор);
2.опосредованное соперничество самцов, привлечение самок особыми демонстрациями и украшениями (межполовой отбор).
Эти два типа полового отбора могут действовать одновременно. Прямое соперничество можно наблюдать во время гона у самцов благородного оленя. Самцы соревнуются в рёве, вероятно, оценивая друг друга на этой стадии соперничества. Иногда они переходят к поединку, сцепив рога, толкают друг друга. Побеждает, как правило, более старший и сильный. Примерно четвёртая часть схваток оканчивается серьёзными ранениями одного из соперников. Ежегодная смена рогов, их отрастание к брачному сезону ложится тяжким бременем на обмен веществ оленя.
Таким образом, представляется вероятным, что в отношении рогов преимущества во внутриполовом отборе превышают негативное давление естественного отбора, который, в свою очередь, привёл к эволюции способов оценки боевого потенциала соперников.
Животные, избегающие прямых драк, будут иметь эволюционное преимущество. При межполовом отборе самки, спаривающиеся с привлекательными самцами, будут иметь и привлекательных сыновей (с большей вероятностью) при условии, что привлекающие признаки наследуются. Эти сыновья, в свою очередь, будут успешно привлекать самок и размножаться.
Таким образом, у самки, выбравшей полового партнёра из-за его сексуальной привлекательности, будет больше потомков.
5. Культурная эволюция - это эволюция поведения и способностей человека. Чтобы понять специфику развития человечества, оценить его возможности, необходимо понять особенности как биологической, так и культурной эволюции.
Биологическая эволюция, будучи саморазвитием органических форм - что признается в настоящее время большинством ученых, основывается на мутационной изменчивости, на борьбе за существование и на естественном отборе. Биологическая преемственность при этом обеспечивается почти полной неизменностью наследственных признаков (генетической памятью) биологических видов. Только мутация приносит изменения в генетическую информацию, поскольку является единственным источником новых организмов в биосфере. Естественный отбор закрепляет лишь те изменения, которые увеличивают способность живого существа к добыванию пищи, самозащиты, размножение, т.е. направленные на повышение ее шансов на выживание, приспособления к жизни.
Биологическая эволюция человека - также результат мутаций, которые оказались в смене деятельности центральной нервной системы, развития мозга. В основе биологической эволюции лежит изменение информации, а не организмов как таковых.
Культурная эволюция во многом похожа на биологическую - информация передается последующим поколениям, но не через размножение с помощью ДНК, а путем подражания, обучения и овладения знаниями и навыками.
Культурная эволюция связана со способностью сохранять всю накопленную информацию и передавать ее следующему поколению.
Культурная эволюция не зависит полностью от случайности - человек способен активно искать новую информацию, сознательно выбирая направление своих поисков. Она может также получить ее путем размышлений, открывая новые факты, взаимосвязи, формы поведения методом дедукции. В культурной эволюции для устранения вредного информации не нужна гибель индивидов. Человек способен сознательно от нее отказаться. Например, мутация, благодаря которой у птицы появляется клюв, может лишить его способности раскалывать зерна или улавливать насекомых. Наоборот, приобретения новой информации в ходе культурной эволюции обычно не сказывается негативно на прежних способностей индивида. Тем-то культурная эволюция может происходить значительно быстрее: ведь новые свойства не вытесняют существующие, а лишь дополняют их. Она имеет кумулятивный характер, и ее темпы с течением времени растут.
С помощью культурной эволюции человек произвела такое поведение, которое постоянно повышает приспособленность к жизни. За очень короткий срок человечество освоило большинство специфических навыков, которыми другие организмы овладевали в течение всей биологической эволюции: мы летаем выше птиц и передвигаемся по воде быстрее любой рыбы.
Человек - единственное живое существо, специфической особенностью которой является культурная эволюция как средство выживания и приспособления к окружающей среде. С помощью культурной эволюции человек постоянно повышает свою приспособленность к жизни путем усовершенствования своего поведения, своего отношения к окружающей действительности.
Таким образом, особенности культурной эволюции и ее отличие от биологической заключаются, во-первых, в способах передачи информации - в подражании (показе, примере), учебе, овладении знаниями и навыками в процессе общения с помощью языка и других знаковых систем. Во-вторых, в характере, содержании и оформлении, закреплении информации - накоплении знаний, оформлении их в науку: искусство, моральные нормы, материальные объекты и т.д. Знание стало первым и неисчерпаемым источником средств выживания человечества. Человек постоянно замечала, открывала и усваивал с пользой для себя все новые и новые взаимосвязи и закономерности типа "если-то", что дало ей возможность преобразовывать окружающий мир в соответствии со своими потребностями. В-третьих, в результатах ее - особенностях организации человеческой жизнедеятельности, становлении социальных взаимоотношений, общества. Когда возник Homo sapiens, все экологические ниши на Земле были заняты другими биологическими видами. И только благодаря возникновению новой формы жизнедеятельности - общества, человечество смогло выжить, расселиться по всей планете и стать доминирующим биологическим видом.
Сущность естественного отбора в теории эволюции заключается в дифференцированном (неслучайном) сохранении в популяции определенных генотипов и избирательном их участии в передаче генов следующему поколению. При этом он влияет не на отдельный признак (или ген), а на весь фенотип, который формируется вследствие взаимодействия генотипа с факторами окружающей среды. Естественный отбор в разных условиях среды будет носить неодинаковый характер.
Следствия естественного отбора:
-Эволюция (изменение, усложнение организмов);
-Возникновение новых видов (увеличение количества [многообразия] видов);
-Приспособленность организмов к условиям окружающей среды. Любая приспособленность относительна, т.е. приспосабливает организм только к одним определенным условиям.

ЭКОЛОГИЧЕСКИЕ ОСНОВЫ ЭВОЛЮЦИИ

Колебания численности (“волны жизни” по С. С. Четверикову, 1905) колебания численности обусловленные элиминацией части особей. ЭЛИМИНАЦИЯ • Периодическая (сезонные и др. циклические процессы). • Непериодическая. • Селективная. • Групповая. • Неизбирательная элиминация (может быть избирательна). ЗНАЧЕНИЕ “ВОЛН ЖИЗНИ”: 1. Случайно меняют концентрации генотипов. 2. Характеризуются повышением и понижением относительной смертности потомства; 3. Выводят ряд генотипов на эволюционную арену.
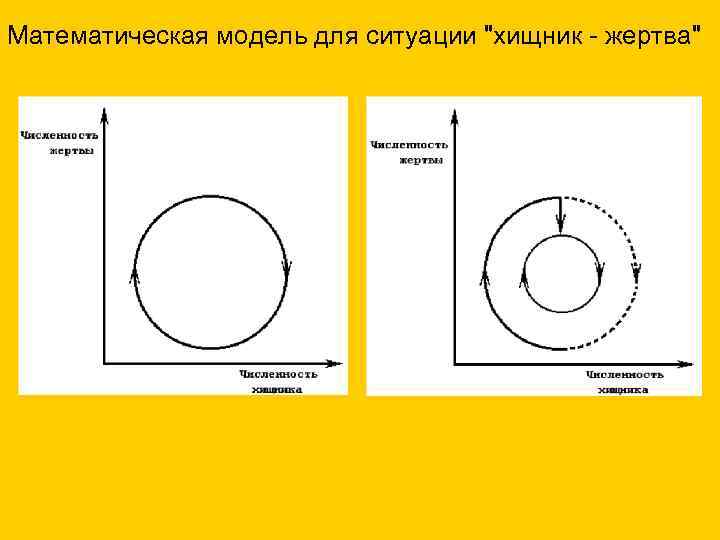
Математическая модель для ситуации "хищник - жертва"

Изоляция 1. Территориально-механическая. 2. Биологическая: • этолого-экологическая, • морфофизиологическая, • генетическая. • прекопуляционные механизмы (предотвращают потерю гамет), • посткопуляционные механизмы (потеря гамет и зигот).
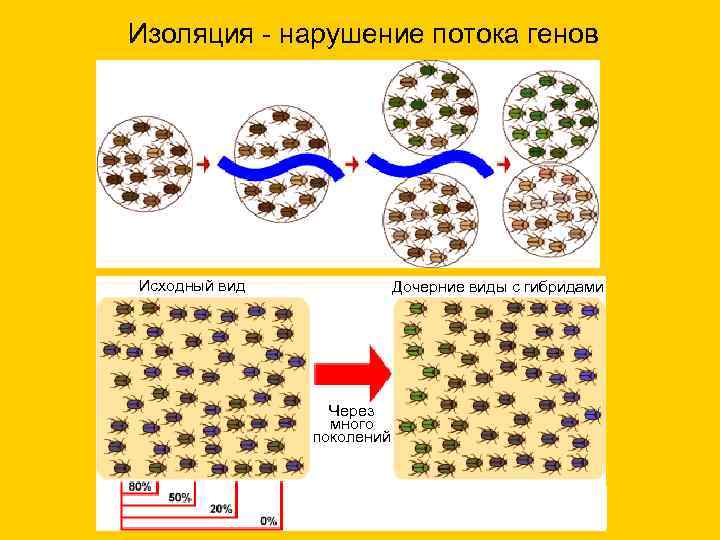
Изоляция - нарушение потока генов Исходный вид Дочерние виды с гибридами Через много поколений

“Принцип основателя” (Э. Майр – founder principle) Особенности новой популяции: · низкое (неполное) генетическое разнообразие в сравнении с исходной популяцией; · неслучайный (лимитированный) набор генов в сформировавшейся популяции. Дальнейшая эволюция будет происходить на основе этого лимитированного изначального фонда
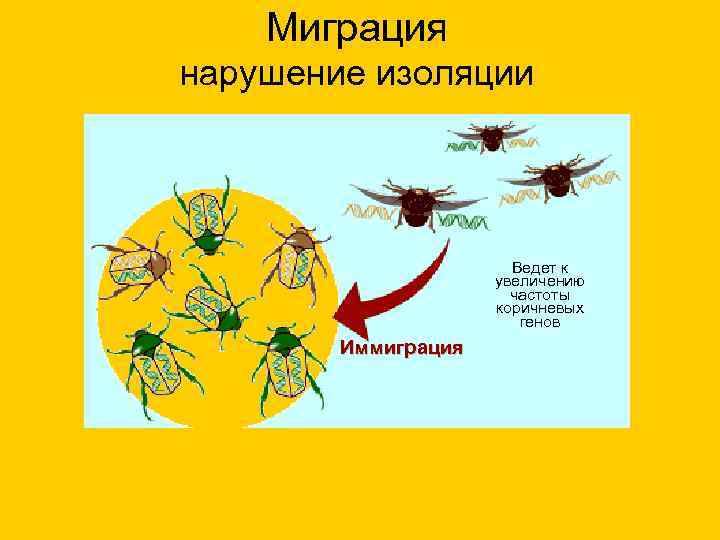
Миграция нарушение изоляции Ведет к увеличению частоты коричневых генов Иммиграция
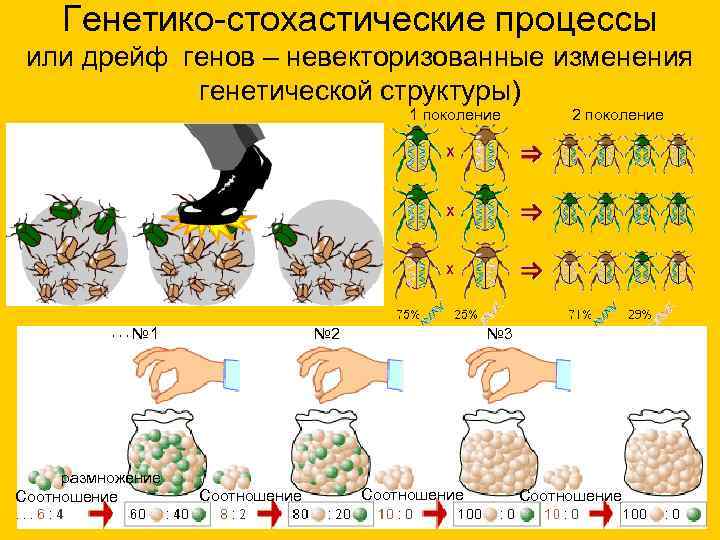
Генетико-стохастические процессы или дрейф генов – невекторизованные изменения генетической структуры) 1 поколение 2 поколение № 1 № 2 № 3 размножение Соотношение

Борьба за существование: 1 - со средой, 2 - внутривидовая, 3 - межвидовая. Принцип конкурентного исключения (competitive-exclusion principle) Г. Ф. Гаузе Два вида не могут сосуществовать неограниченно долго в одной и той же местности, занимая одинаковые экологические ниши. Результат – расхождение по разным экологическим нишам, или – элиминация. книга “Борьба за существование”, 1934 г. , Балтимор

Коэволюция Центральноамериканская Acacia и муравьи, потребляющие нектар, обитающие в полых шипах и защищающие ее от фитофагов.
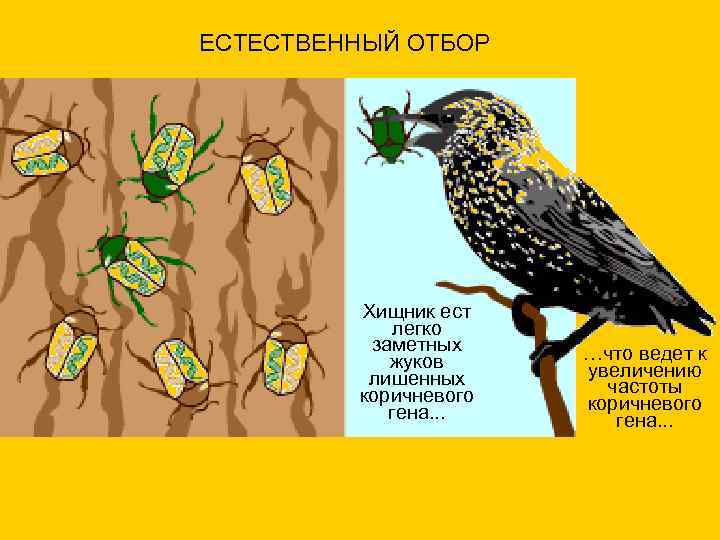
ЕСТЕСТВЕННЫЙ ОТБОР Хищник ест легко заметных …что ведет к жуков увеличению лишенных частоты коричневого гена. . .
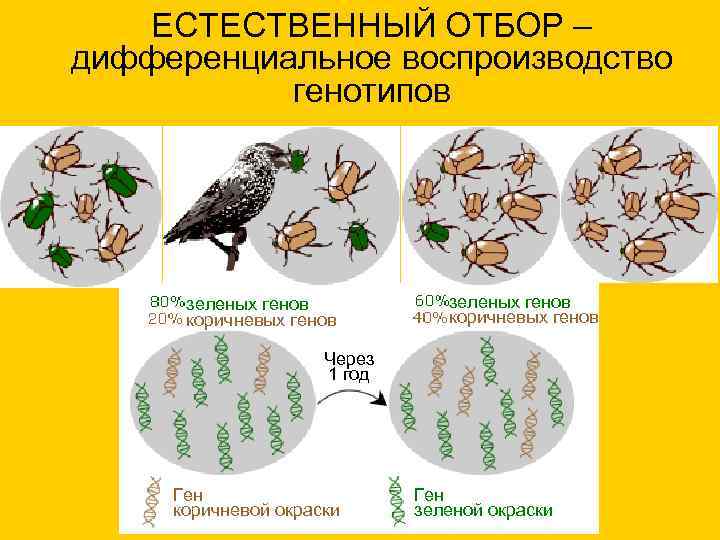
ЕСТЕСТВЕННЫЙ ОТБОР – дифференциальное воспроизводство генотипов зеленых генов коричневых генов Через 1 год Ген коричневой окраски зеленой окраски
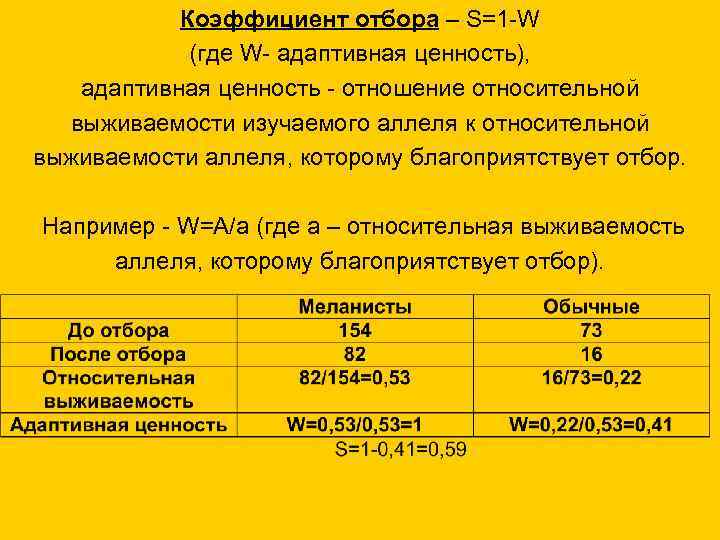
Коэффициент отбора – S=1 -W (где W- адаптивная ценность), адаптивная ценность - отношение относительной выживаемости изучаемого аллеля к относительной выживаемости аллеля, которому благоприятствует отбор. Например - W=A/a (где a – относительная выживаемость аллеля, которому благоприятствует отбор).

Групповой отбор – селекция на признаки, неблагоприятные для особи и благоприятные для популяции в целом. Половой отбор – форма естественного отбора, касающаяся признаков особей одного пола. Различные модели размножения – полиандрия, полигиния, моногамия. Принципы дифференциального спаривания: преимущественное спаривание гетерозигот, уделение предпочтения редким фенотипам и т. п.

Половой отбор 20 лет 40 лет 65 лет Джо размножается, Дети Джо размножаются, имеет детей имеют детей Джим не передал своих размножается генов ·Соревнование самцов ·Выбор самок

ПОЛОВОЕ РАЗМНОЖЕНИЕ КАК СРЕДСТВО УВЕЛИЧЕНИЯ ИЗМЕНЧИВОСТИ Организмы, способные к бесполому размножению, в критической ситуации резко переходят к размножению половому. Бесполое размножение дает клоны. Исходно половой процесс использовался именно как средство экстренного повышения изменчивости. Потомство, возникшее в результате полового размножения: во-первых разнообразнее, во-вторых не является точной копией родителей
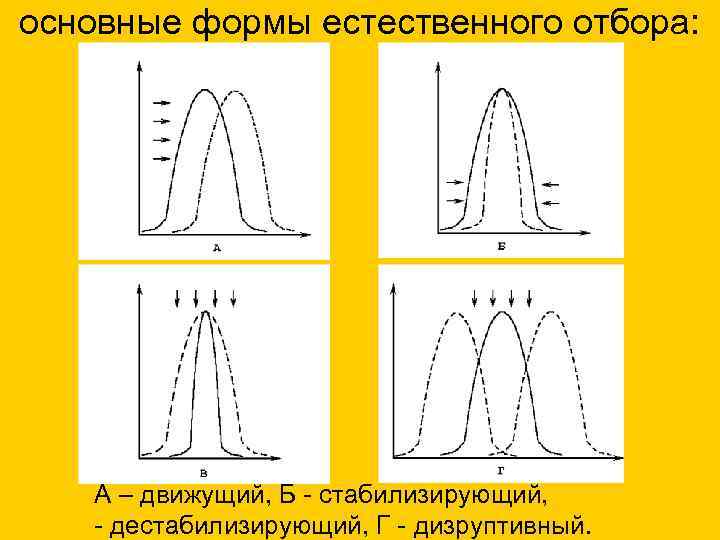
основные формы естественного отбора: А – движущий, Б - стабилизирующий, - дестабилизирующий, Г - дизруптивный.
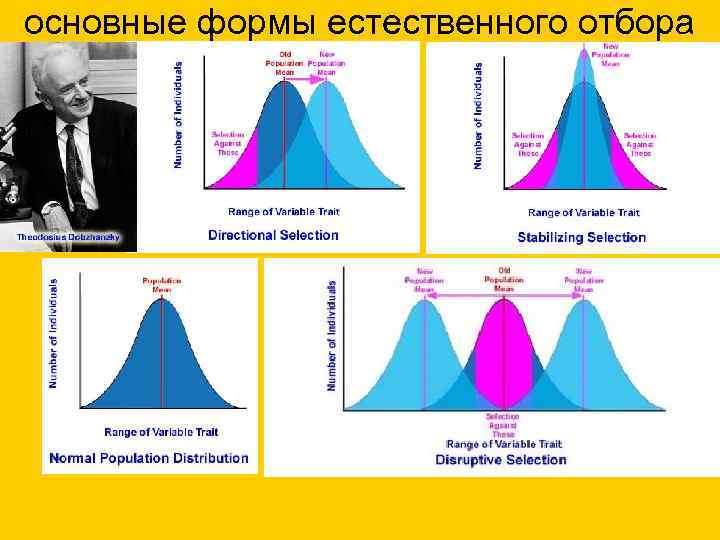
основные формы естественного отбора
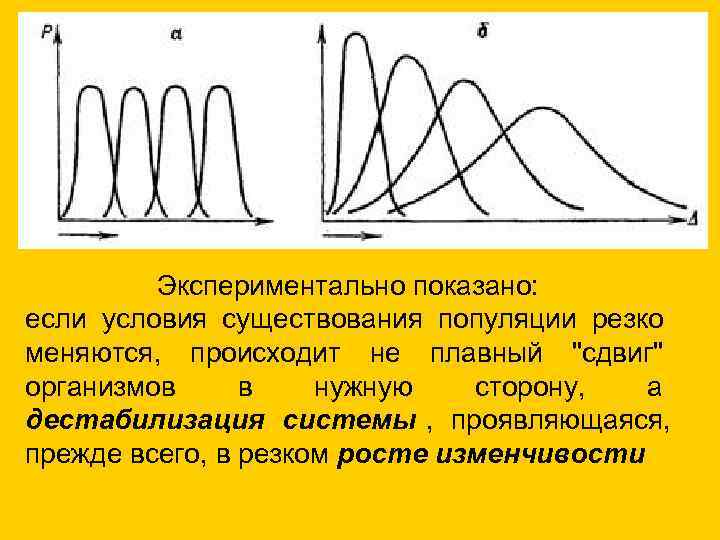
Экспериментально показано: если условия существования популяции резко меняются, происходит не плавный "сдвиг" организмов в нужную сторону, а дестабилизация системы , проявляющаяся, прежде всего, в резком росте изменчивости
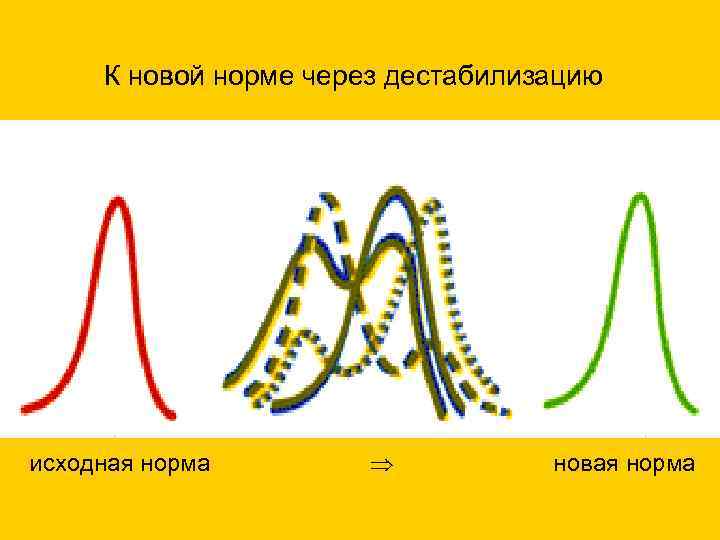
К новой норме через дестабилизацию исходная норма новая норма
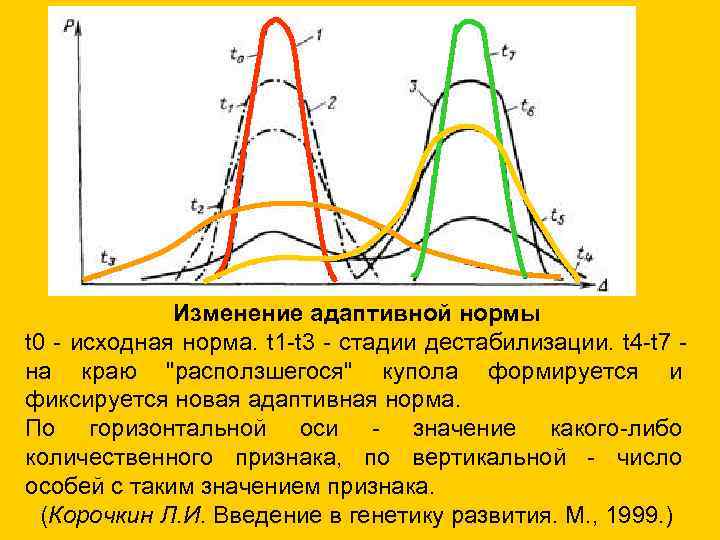
Изменение адаптивной нормы t 0 - исходная норма. t 1 -t 3 - стадии дестабилизации. t 4 -t 7 - на краю "расползшегося" купола формируется и фиксируется новая адаптивная норма. По горизонтальной оси - значение какого-либо количественного признака, по вертикальной - число особей с таким значением признака. (Корочкин Л. И. Введение в генетику развития. М. , 1999. )

Искуственный отбор в лаборатории Эксперименты Джона Эндлера. Характером пятнистости гуппий можно манипулировать с помощью искусственного отбора. крупный гравий+хищник мелкий гравий+хищник крупный гравий без хищника мелкий гравий без хищника менее 15 поколений отбора отбора
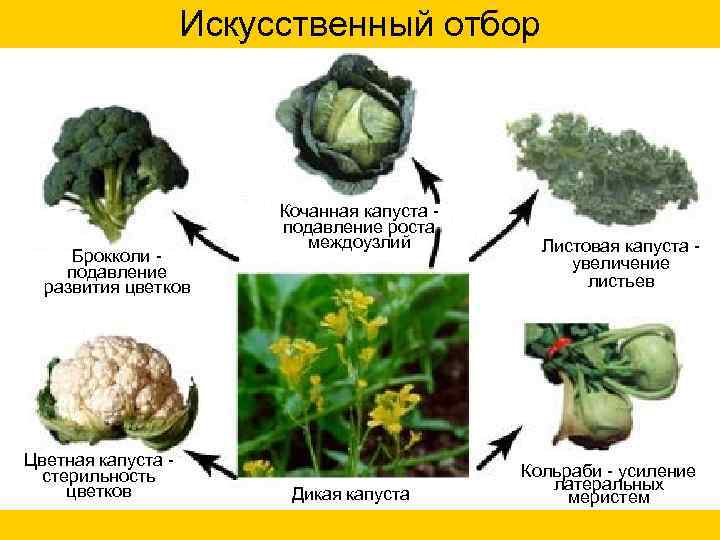
Искусственный отбор Кочанная капуста - подавление роста междоузлий Листовая капуста - Брокколи - увеличение подавление листьев развития цветков Цветная капуста - стерильность Кольраби - усиление цветков латеральных Дикая капуста меристем

Искусственный отбор

Искусственный отбор

Искусственный отбор

Особенности пород домашних животных и сортов культурных растений • 1) они представлены большим многообразием; • 2) породы и сорта одной группы отличаются как между собой, так и от дикого предка; • 3) они отвечают нуждам человека. Ч. Дарвин Изменение животных и растений в одомашненном состоянии (1868).

Columbia livia

Монофилия пород Columbia livia • все они имеют сходные признаки в строении; • все породы скрещиваются между собой и с диким голубем и дают плодовитое потомство; • в потомстве от таких скрещиваний часто наблюдаются уклонения в сторону дикого голубя; • все породы имеют одинаковые повадки (строительный инстинкт, вынашивание и кормление птенцов и т. д. ); • все породы способны к одичанию, и при этом наблюдается утрата породных признаков

Условия искусственного отбора: • 1) усиление изменчивости (большое число особей); • 2) устранение нежелательных скрещиваний; • 3) строгая методичность ведения селекции; • 4) наблюдательность и опыт действие искусственного отбора ограничено во времени и численностью особей
Содержание работы
Введение…………………………………………………………………. 3
1. Основная часть…..………………………………………………..……… 5
1.1. Популяция, как элементарная единица эволюции. …………………….6
1.2. Борьба за существование………………. ……………………………. 13
1.3. Биогеоценоз как арена борьбы за существование. Коэволюция………………………………………. ………………. 19
1.4. Динамика численности……………………… ……………. ……. 21
Заключение………………………………………………………….………. 23
Список использованной литературы………………………………………. 25
Файлы: 1 файл
моя курсовая.docx
Федеральное государственное бюджетное образовательное учреждение высшего профессионального образования
Факультет ___________________ПИЩЕВЫХ ПРОИЗВОДСТВ___________________ ____
Кафедра ___________ТЕХНОЛОГИИ ПИЩЕВЫХ ПРОИЗВОДСТВ _
И ПАРФЮМЕРНО-КОСМЕТИЧЕСКИХ ПРОДУКТОВ_____________
На тему Экологические основы эволюции
Дисциплина Общая биология и микробиология
Принял к исполнению Научный руководитель
Студент __III – ФПП – 5 ______________________
__Пономарева Ю.В ______Руденко Е.Ю.____
1.1. Популяция, как элементарная единица эволюции. …………………….6
1.2. Борьба за существование………………. ……………………………. 13
1.3. Биогеоценоз как арена борьбы за существование. Коэволюция………………………………………. ………………. 19
1.4. Динамика численности……………………… ……………. ……. 21
Список использованной литературы………………………………………. 25
Чарльз Дарвин – выдающийся английский естествоиспытатель – создал эволюционную теорию, основные положения которой не утратили своего значения и в наше время.
Независимая эволюция изолированных видов приводит к дивергенции – усилению различий между организмами разных видов – и к повышению видового разнообразия. Смена менее приспособленных видов более приспособленными (вследствие принципа конкурентного исключения) приводит к общей прогрессивной эволюции органического мира Земли.
1. Основная часть
В 1960 г. академик И. И. Шмальгаузен сформулировал представление об эволюции, как об авторегуляторном процессе, основанном на обратной связи. Эта модель дает наиболее наглядное представление о механизме эволюции и ее факторах - движущих силах. Поэтому с нее удобно начать рассмотрение микроэволюции. Микроэволюция - это условно выделяемый раздел теории эволюции, рассматривающий процессы, протекающие на уровне популяций, начиная с механизмов формирования изменчивости, в первую очередь наследственной, до возникновения нового вида.
Экологические основы эволюции включают два основных вопроса: 1) характеристику популяции, как элементарной эволюционирующей единицы; 2) характеристику взаимодействий популяции как целого с ее экологическим окружением и характеристику взаимодействий особей, составляющих популяцию, как между собой, так и с внешними по отношению к популяции факторами среды.
Эволюционной экологии, как на популяционном, так и на экосистемном уровнях посвящено множество публикаций. Потому, сделана попытка сконцентрироваться лишь на тех аспектах этой науки, которые важны для понимания эволюции [1].
1.1. Популяция, как элементарная единица эволюции
Элементарная единица эволюции – биологическая система, в которой протекает эволюционный процесс. Исходя из определения эволюции, элементарная эволюционная единица должна:
- быть целостной в пространстве и во времени;
- постоянно наследственно изменяться во времени;
- реально существовать в природе.
Эти черты в комплексе впервые проявляются на популяционном уровне. Поэтому современная эволюционная биология в качестве элементарной единицы эволюции рассматривает популяцию [2].
Популяция – это совокупность особей данного вида, занимающих определенный участок видового ареала, свободно скрещивающихся между собой и частично или полностью изолированных от других популяций.
Популяция существует у всех видов живых организмов. Некоторые виды состоят из огромного числа популяций, другие - из немногих или даже из одной популяции. Популяция характеризуется пространственным распределением, определенной численностью, динамикой численности, системой размножения, возрастным и половым составом. Характеристики популяции оказывают влияние на действие эволюционных факторов [3].
Пространственное распределение. По пространственному распределению выделяют три основных типа популяций:
1) большие непрерывные популяции ( популяции злаков, растущих на равнинах и покрывающих площади шириной в десятки или сотни километров);
2) мелкие изолированные колониальные популяции (популяции, соответствующие островному типу). К такому типу популяций относятся популяции наземных животных, обитающих на архипелагах; пресноводные формы, населяющие цепь озер, обитатели горных вершин в горной местности и организмы, ограниченные определённым типом почвы или горной породы с пятнистым распределением;
3) линейные популяции. Линейные популяции возникают вдоль рек, на побережьях морей и в аналогичных местообитаниях, обладающих большой протяженностью и более или менее непрерывных в одном измерении, но ко- ротких и ограниченных в другом [4].
Кроме того, существуют разнообразные варианты, промежуточные между этими трёмя основными типами. Так большая популяция может быть непрерывной в одних частях ареала, но прерывистой или полунепрерывной в других. Подобным же образом колонии, населяющие систему островов, могут быть изолированы лишь частично, а не полностью.
Численность особей. В связи с размерами ареала и биологическими особенностями организмов численность особей в популяции значительно варьирует. Так одни популяции могут быть незначительными по численности (десятки особей), другие – огромными (миллионы особей).
Динамика численности. Размеры популяций подвержены значительным колебаниям. Причины колебаний численности довольно разнообразны, но в общей форме сводятся к влиянию абиотических и биотических факторов.
Системы размножения. Системы размножения варьируют от панмиксии до самооплодотворения. Часто встречаются промежуточные типы (например, скрещивание между сибсами у животных и др.).
Возрастной и половой состав. Популяция состоит из разных по полу и возрасту особей. Для каждого вида и популяции характерны свои соотношения возрастных групп. На эти соотношения влияют общая продолжительность жизни, время достижения половой зрелости, интенсивность размножения [5].
Каждая популяция имеет определенную генетическую структуру и генофонд.
Генофонд популяции – совокупность генотипов всех особей в популяции.
Генетическая структура популяции определяется концентрацией каждого гена (или его аллеля) в популяции, характером генотипов и частотой их распространения. Генетическая структура сохраняется в ряду поколений до тех пор, пока какой-либо фактор не выведет ее из состояния равновесия.
Для изучения генетической структуры популяции анализируется выборка особей из этой популяции. Чем больше эта выборка, тем точнее она представляет реальное соотношение частот генотипов в популяции. В качестве материала используются полевые наблюдения, данные лабораторных анализов полевых сборов или музейных экземпляров и даже архивные данные [2].
Известно несколько факторов, которые могут нарушать равновесие генотипов в популяции:
• низкая численность популяций;
Реально существующие популяции очень разнообразны по величине и форме. Структура популяции слагается из четырёх главных компонент: величины популяции, пространственной конфигурации, системы размножения и скорости расселения. Что касается систем размножения, то их диапазон очень широк — от свободного неродственного скрещивания до самооплодотворения. Часто встречаются такие промежуточные типы, как свободное скрещивание между близкими соседями; инбридинг, осуществляемый иными способами, нежели самооплодотворение (например скрещивание между сибсами у животных); сочетание неродственного скрещивания с самоопылением, как у гермафродитных, но самосовместимых цветковых растений. Варианты пространственного распространения и систем размножения встречаются во всевозможных сочетаниях, создавая в результате чрезвычайно разнообразные структуры популяций. Так, большая непрерывная популяция может состоять из свободно скрещивающихся особей, как у многих опыляемых ветром травянистых растений равнин, но она может также состоять из особей с ограниченной свободой скрещивания или из инбредных особей. Подобно этому небольшая изолированная колония может состоять или из свободно скрещивающихся или из инбредных особей.
Структура популяции оказывает влияние на характер её изменчивости.
Отдельные особи не могут эволюционировать. Каждая особь развивается на основе генотипа, унаследованного от родителей. Генотип определяет особенности ее развития, ее взаимоотношения с внешней средой, в том числе и возможность адаптивных модификаций в ответ на изменение внешних условий. Но как бы ни менялась особь, ее генотип остается неизменным. Таким образом, элементарной единицей эволюции является не особь, а популяция. Совокупность называют генофондом. В ходе эволюции меняется набор генотипов в генофонде популяций. Одни генотипы распространяются, а другие становятся редкими и постепенно исчезают [4].
Следует выделить четыре основных фактора эволюции: мутационный процесс, изоляция, популяционные волны (и дрейф генов), а также единственный направленный фактор эволюции - естественный отбор.
Мутационный процесс
Мутации (лат. mutatio — изменение) - стойкое изменение генома (наследственного материала), которое может быть унаследовано потомками организма. Процесс возникновения мутаций - мутагенез.
- Генные
- Хромосомные
- Геномные
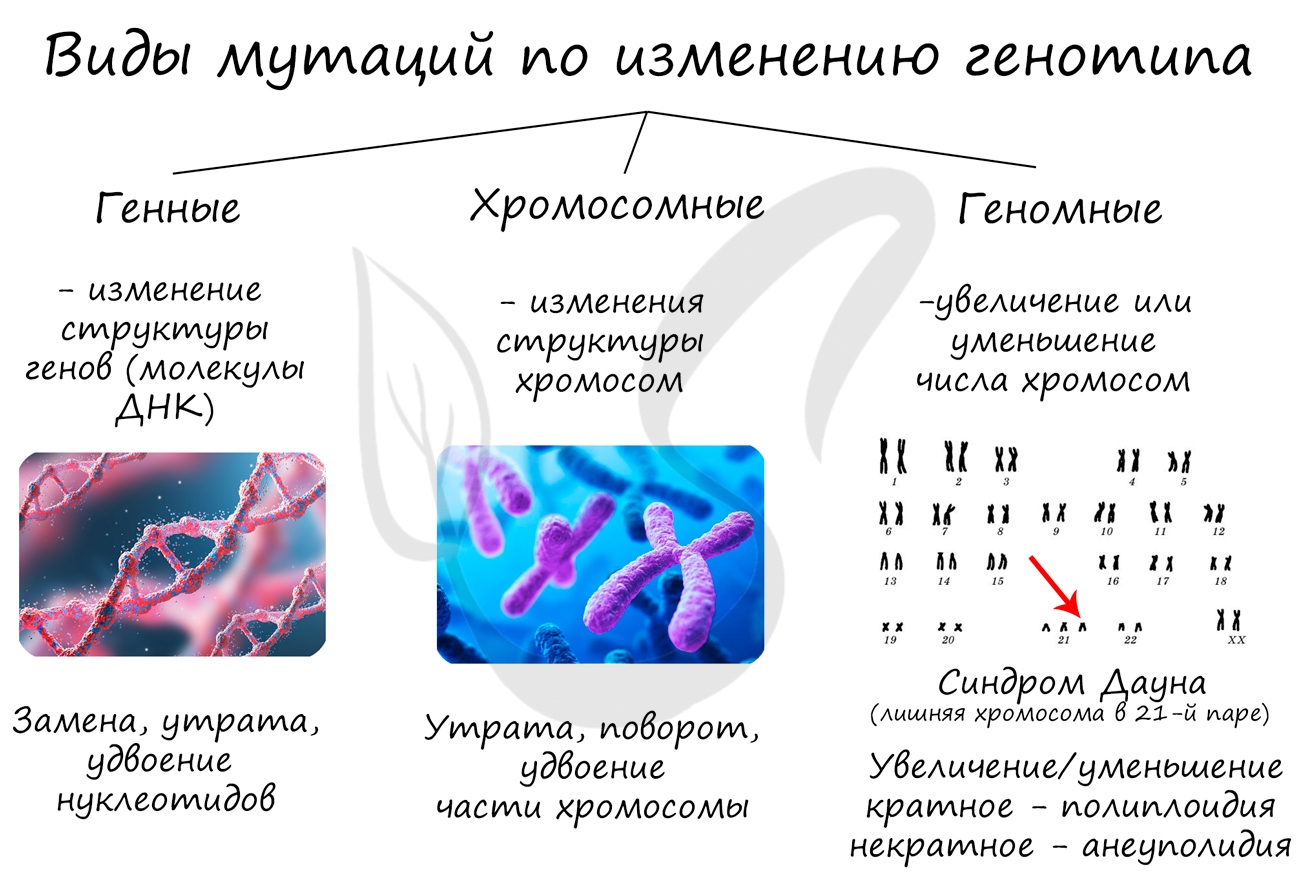
Большинство мутаций возникает спонтанно и вредит организму. Часть мутаций являются рецессивными, поэтому не проявляются и передаются многим поколениям, накапливаясь в генофонде популяции.
Мутации напоминают колоду карт: неизвестно, что выпадет - чаще всего это карты невысокого ранга, козыри - большая удача. Так и мутации, большинство из них вредные, приводят к развитию опухолей. Полезные встречаются гораздо реже. Как игра в карты, все подчиняется случайности.

Популяционные волны
- Сезонные изменения, периодические изменения какого-либо значимого фактора среды
- Непериодические изменения, например, в результате природных катастроф, изменение численности популяций хищник-жертва
- Заселение новых территорий, ярким примером которого является бурный рост численности кроликов, завезенных в Австралию
Колебание численности популяций по типу "хищник - жертва" является классическим примером популяционных волн. Представим себе популяцию зайцев (жертв), которая бурно увеличилась в численности. Зайцами питаются лисица, волк (хищники). С увеличением их кормовой базы (зайцев) наблюдается и рост численности хищников, которые поедают зайцев, вследствие чего численность зайцев снижается. С уменьшением кормовой базы, снижается и число хищников. Так в природе устанавливается баланс между хищниками и жертвами.

Особенно весомым фактором эволюции популяционные волны выступают в небольших популяциях. Их участие в эволюционном процессе основано на явлении дрейфа генов.
Форма гена - аллель, с которым вы подробнее познакомитесь в ходе изучения генетики, встречается в популяции с определенной частотой. Дрейф генов - изменение частоты встречаемости аллельных вариантов генов.
Дрейф генов обусловлен случайными причинами: у особей образуются гаметы, несущие различные формы аллельных генов. Не все из гамет принимают участие в процессе оплодотворения: здесь вновь руководит случайность. Вследствие этого одни аллельные формы генов могут встречаться в популяции часто, другие - редко.
Если представить, что часть особей, составляющих одну популяцию, погибли по тем или иным причинам, то редкие гены в оставшихся особях могут увеличить свою частоту, то есть в результате размножения оставшихся особей редкие гены начнут встречаться более часто - это и есть дрейф генов.
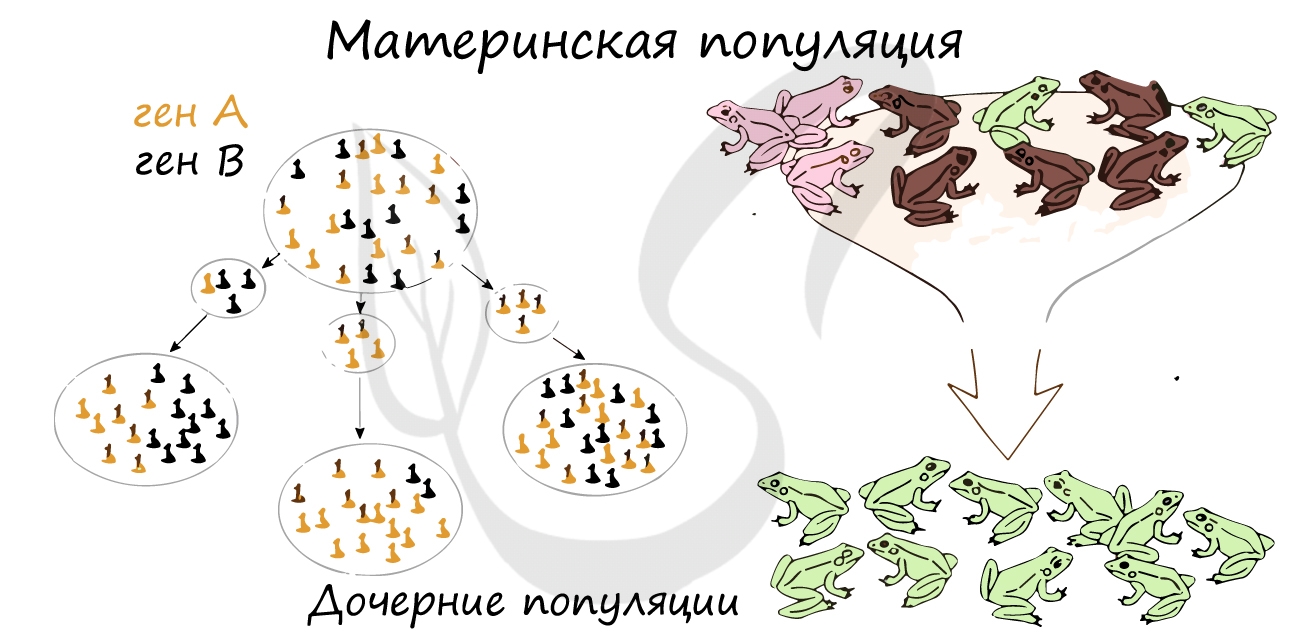
В закрытых популяциях не только животных, но и людей - в религиозных общинах, происходит возрастание гомозиготности популяции, что приводит к снижению ее жизнеспособности и проявлению редких аллелей.
Такое повышение встречаемости аллелей возникает в результате близкородственных браков: проявляются редкие гены, которые часто приводят к заболеваниям.

Изоляция
Изоляцией называют невозможность или затруднение свободного скрещивания между особями одного вида. Вследствие этого, генофонды двух популяций становятся независимыми друг от друга. Внутри каждой популяции происходит генотипическая дифференцировка из-за их разобщенности.
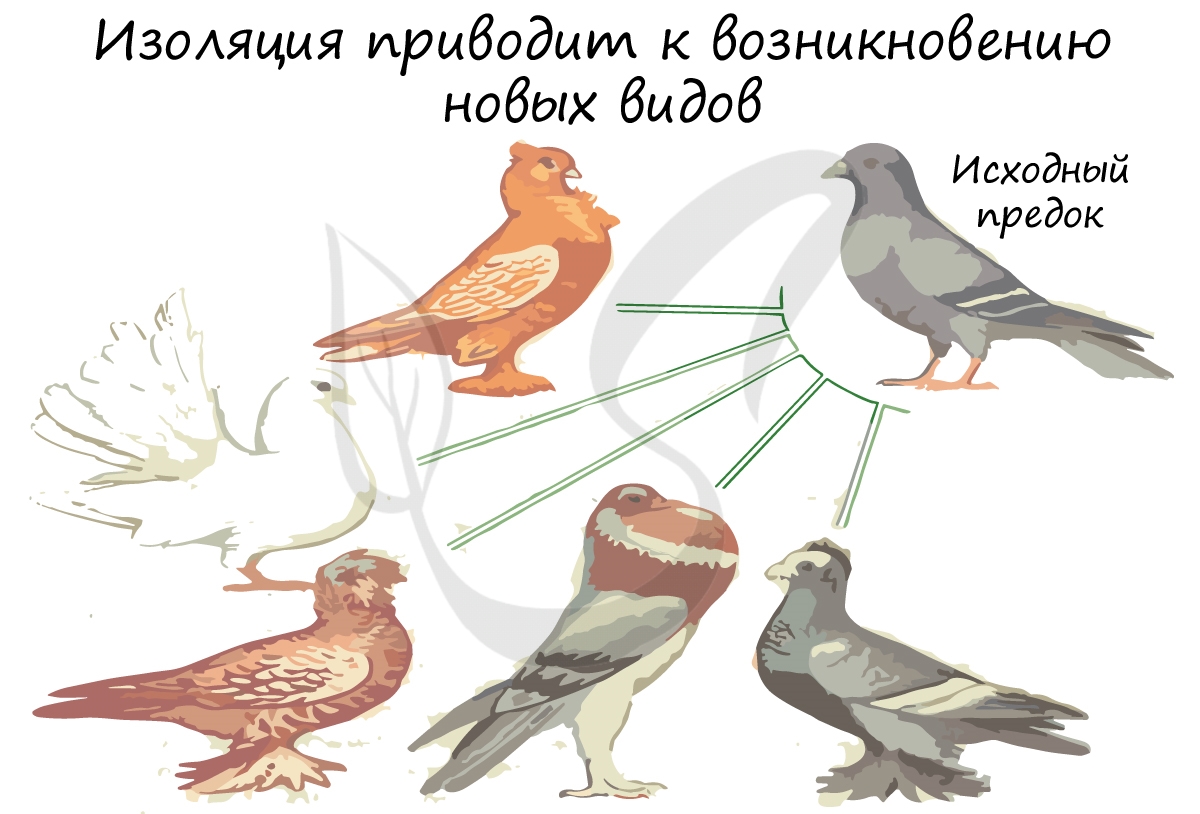
Географическая (греч. geo - земля) изоляция может возникать вследствие географических барьеров - пустыни, горы, водоемы.
- Экологическая - особи обитают на одной территории, но в различных местах обитания (к примеру, разделены друг от друга непроходимой чащей)
- Временная - изоляция вследствие разновременности половой активности, периода цветения
- Этологическая - изоляция вследствие различного брачного поведения
- Механическая - отличия в строении половых органов, невозможность спаривания

Естественный отбор
Изученные нами факторы эволюции: мутации, популяционные волны и дрейф генов, изоляция - все они носят случайный, ненаправленный характер. Они приводят к появлению различных признаков у отдельных особей, которые могут быть как полезны, нейтральны, так и вредны для особи.
Таким образом, перечисленные факторы создают основу, "базу" для действия единственного направленного фактора эволюции - естественного отбора. В ходе естественного отбора особи с полезными признаками, которые помогают им приспособиться к условиям внешней среды и способствуют выживанию, остаются и размножаются, а особи без этих признаков выживают реже и не продолжают род.

Закон естественного отбора безапелляционно провозглашает: будь приспособлен - или умри. Выживает в природе не самый сильный, а самый приспособленный. Иногда выжить животным помогает и сила, но гораздо больше других примеров. Многие животные сливаются с окружающей средой: приобретают покровительственную окраску (мимикрию), которая делает их незаметными.
Иногда безобидные животные, в результате приспособления к внешней среде, приобретают окраску тела, напоминающую окраску опасных хищных животных. Примером может послужить внешнее сходство мухи из семейства журчалок с осой.

Многие хорошо защищенные, ядовитые виды в ходе естественного отбора получили яркую, так называемую предупреждающую окраску. Эта окраска предупреждает хищников об опасности. Если хищник съест такое ядовитое животное, то рискует получить тяжелую интоксикацию и погибнуть.

Теперь вы понимаете, что признаки животных - различные формы их тела и окраска - являются приспособлениями к условиям внешней среды, это - полезные признаки, которые в ходе естественного отбора позволили животным выжить и размножиться. Таким образом, естественный отбор это отбор особей, с наиболее приспособленным к среде фенотипом.
Необходимо осознавать относительность приспособленности к окружающей среде. Она помогает выживать лишь при определенных условиях, и, если условия меняются, то окраска может оказаться вовсе не полезной, но даже и вредной. К примеру, при таянии снега заяц-беляк становится еще более заметен на голой земле.

- Генетическое разнообразие особей, на основе которого возникают различные признаки
- Способность к неограниченному размножению (избыточность потомства)
- Борьба за существование
Самая ожесточенная борьба. Происходит между особями, принадлежащими к одному виду. Благодаря внутривидовой борьбе происходит половой отбор: к размножению редко допускаются неприспособленные особи, род продолжают лучшие из лучших.
Возникает между особями, которые принадлежат к разным видам. Более приспособленный к условиям среды вид побеждает и размножается, менее приспособленный - проигрывает и вымирает. Примером могут послужить формы взаимодействий: хозяин-паразит, хищник-жертва, симбиоз.
В изменяющихся условиях внешней среды выживают наиболее приспособленные особи. Примером такой борьбы являются сезонные миграции птиц, зимняя спячка у животных.

Формы естественного отбора
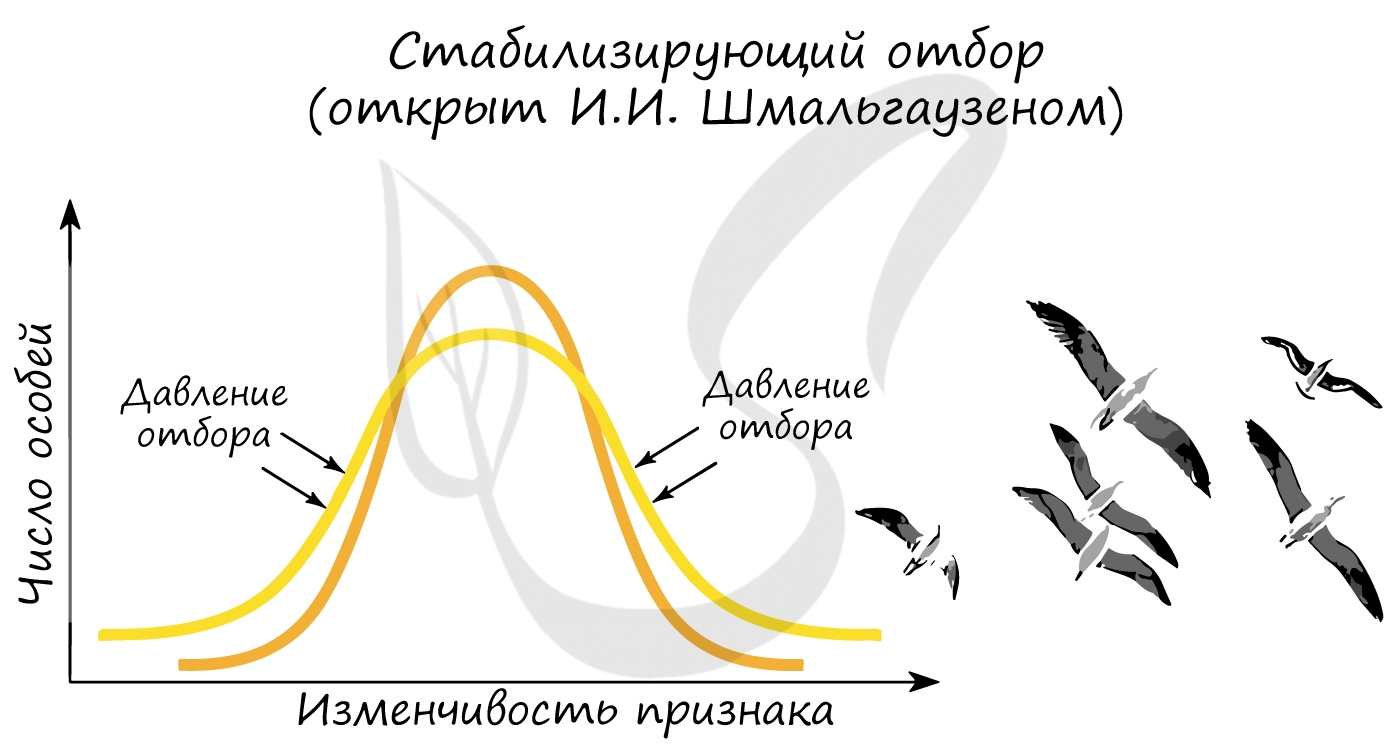
Открыт И.И. Шмальгаузеном. Стабилизирующий отбор приводит к сужению нормы реакции, устраняя отклонения от нее. В результате преимущество получают особи, обладающие средней степенью признака, который характерен для вида или популяции. Этот отбор действует при стабильных (неизменных) условиях среды.
Примером действия стабилизирующего отбора может послужить буря: во время бури чаще всего выживают птицы со средней длиной крыла, тогда как особи с слишком короткими, или слишком длинными крыльями погибают.
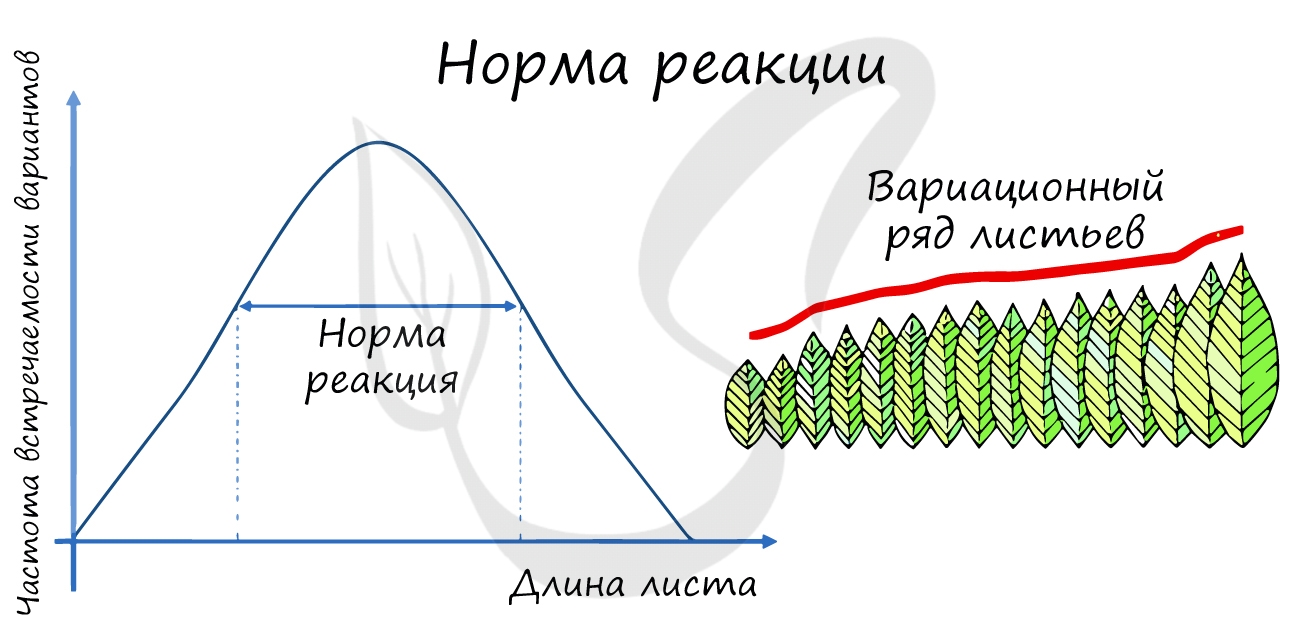
Новый термин, который вы увидели - норма реакции - подразумевает способность генотипа, в зависимости от условий среды, формировать различные фенотипы.
- C узкой нормой реакции - цвет глаз, число пальцев у человека, окраска цветов растения
- C широкой нормой реакции - рост и вес человека, размеры листьев растения
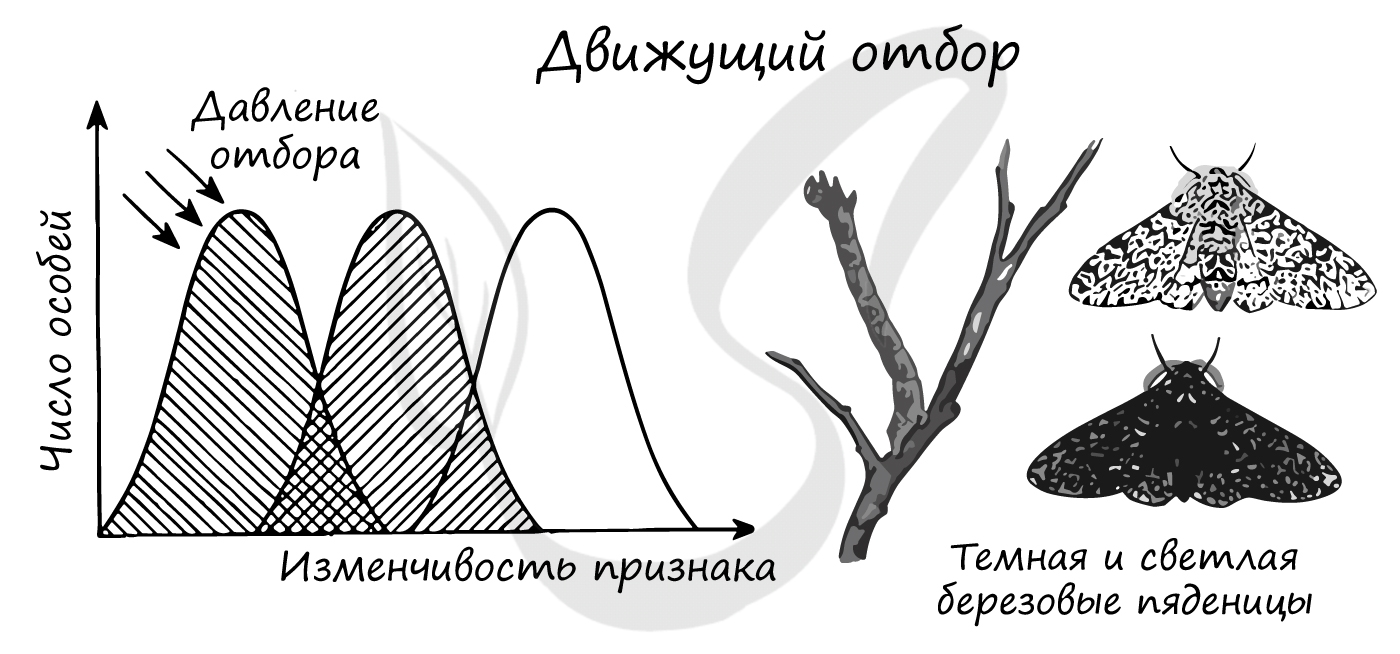
Движущий естественный отбор приводит к смещению нормы реакции, в результате чего изменяется среднее значение признака. Этот вид отбора действует при изменяющихся условиях среды.
Известным примером является индустриальный меланизм - возникновение меланистических форм животных (греч. melanos — чёрный), отличающихся темным окрасом. Это явление началось в Англии со второй половины XIX века вследствие бурного развития промышленности.
Из-за копоти, оседающей на поверхности стволов деревьев, бабочки со светлой окраской - берёзовые пяденицы - стали заметны на стволах деревьев и легко поедались птицами. В результате остались только приспособленные - бабочки с темным окрасом, которые были незаметны на стволах деревьев, вследствие чего они выживали и размножались.
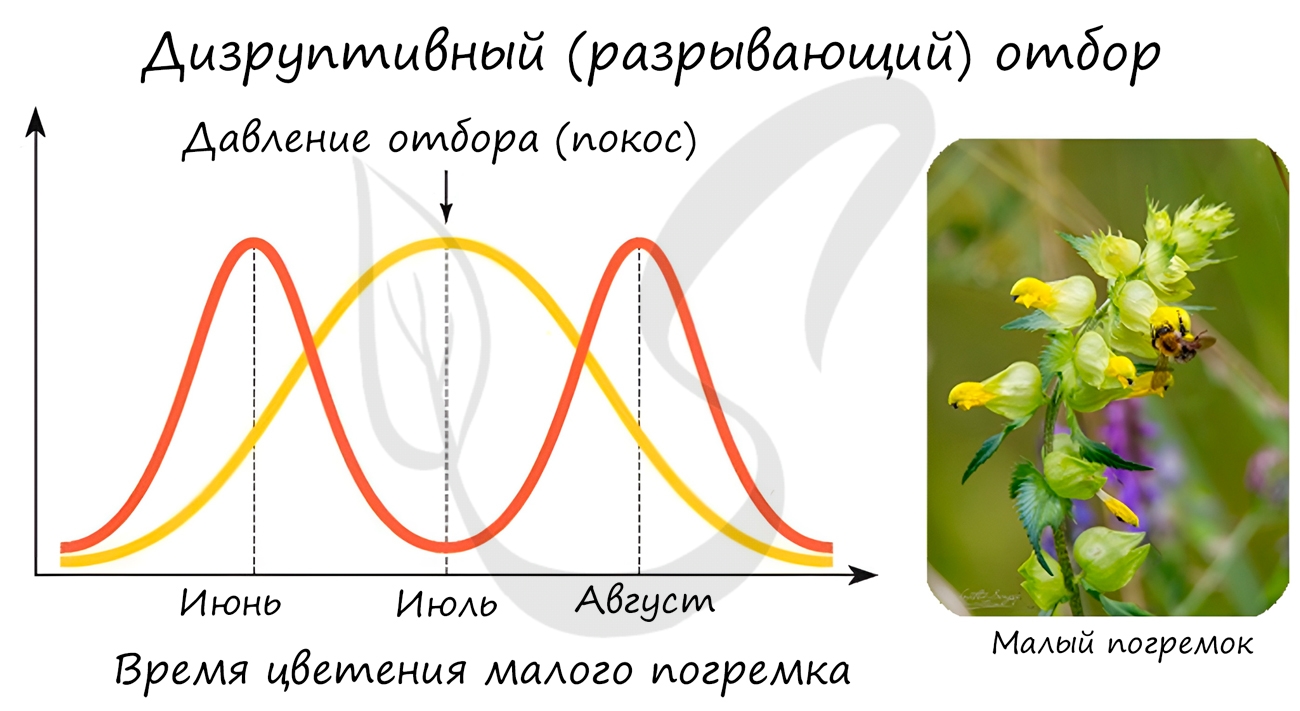
Направлен на сохранение в популяции крайних значений признаков, не благоприятствует среднему промежуточному значению признака. В результате в популяции сохраняется более чем одно значение признака.
Типичным примером является появление в луговых сообществах раноцветущих и поздноцветущих растений. В результате летних покосов, особи со средним значением признака, у которых цветение приходит на середину лета, постепенно исчезают из популяции растений. Выживают и размножаются только те растения, у которых цветение происходит до или после покосов.
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
3. Мутационный процесс
4. Колебания численности (“волны жизни” по С.С.Четверикову, 1905)
колебания численности обусловленные элиминацией части особей.
ЭЛИМИНАЦИЯ
•Периодическая (сезонные и др. циклические процессы).
•Непериодическая.
•Селективная.
•Групповая.
•Неизбирательная элиминация (может быть избирательна).
ЗНАЧЕНИЕ “ВОЛН ЖИЗНИ”:
1.Случайно меняют концентрации генотипов.
2. Характеризуются повышением и понижением
относительной смертности потомства;
3. Выводят ряд генотипов на эволюционную арену.
8. Изоляция
1. Территориально-механическая.
2. Биологическая:
• этолого-экологическая,
• морфофизиологическая,
• генетическая.
•прекопуляционные механизмы
(предотвращают потерю гамет),
•посткопуляционные механизмы
(потеря гамет и зигот).
“Принцип основателя”
(Э.Майр – founder principle)
Особенности новой популяции:
низкое (неполное) генетическое
разнообразие в сравнении с исходной
популяцией;
неслучайный (лимитированный) набор
генов в сформировавшейся популяции.
Дальнейшая эволюция будет происходить на
основе этого лимитированного изначального
фонда
13. Миграция нарушение изоляции
14. Генетико-стохастические процессы или дрейф генов – невекторизованные изменения генетической структуры)
15. Борьба за существование: 1- со средой, 2- внутривидовая, 3- межвидовая. Принцип конкурентного исключения (competitive-exclusion
principle) Г.Ф.Гаузе
Два вида не могут сосуществовать неограниченно
долго в одной и той же местности, занимая
одинаковые экологические ниши. Результат –
расхождение по разным экологическим нишам, или –
элиминация.
книга “Борьба за существование”, 1934 г., Балтимор
18. Коэволюция
Центральноамериканская Acacia и муравьи, потребляющие
нектар, обитающие в полых шипах и защищающие ее от
фитофагов.
ЕСТЕСТВЕННЫЙ ОТБОР
Хищник ест
легко
заметных
жуков
лишенных
коричневого
гена.
…что ведет к
увеличению
частоты
коричневого
гена.
20. ЕСТЕСТВЕННЫЙ ОТБОР – дифференциальное воспроизводство генотипов
зеленых генов
коричневых генов
зеленых генов
коричневых генов
Через
1 год
Ген
коричневой окраски
Ген
зеленой окраски
21. Коэффициент отбора – S=1-W (где W- адаптивная ценность), адаптивная ценность - отношение относительной выживаемости изучаемого
аллеля к относительной
выживаемости аллеля, которому благоприятствует отбор.
Например - W=A/a (где a – относительная выживаемость
аллеля, которому благоприятствует отбор).
До отбора
После отбора
Относительная
выживаемость
Адаптивная ценность
Меланисты
154
82
82/154=0,53
W=0,53/0,53=1
S=1-0,41=0,59
Обычные
73
16
16/73=0,22
W=0,22/0,53=0,41
22. Групповой отбор – селекция на признаки, неблагоприятные для особи и благоприятные для популяции в целом. Половой отбор – форма
естественного
отбора, касающаяся признаков особей
одного пола.
Различные модели размножения –
полиандрия, полигиния, моногамия.
Принципы дифференциального спаривания:
преимущественное спаривание гетерозигот,
уделение предпочтения редким фенотипам
и т.п.
Половой отбор
20 лет
Джо
Джим
40 лет
65 лет
Джо размножается,
имеет детей
Джим не
размножается
Соревнование самцов
Выбор самок
Дети Джо размножаются,
имеют детей
Джим не передал
своих генов
25. "Пока два петуха дерутся, третий топчет кур!" (Н.В.Тимофеев-Ресовский) «…пока крупные самцы каракатиц дерутся, мелкий
26. «… большинство видов вообще прекрасно обходится без полового отбора, и именно этот факт и наводит на мысль, что такого фактора
ПОЛОВОЕ РАЗМНОЖЕНИЕ КАК СРЕДСТВО
УВЕЛИЧЕНИЯ ИЗМЕНЧИВОСТИ
Организмы, способные к бесполому размножению, в
критической ситуации резко переходят к размножению
половому. Бесполое размножение дает клоны.
Исходно половой процесс использовался именно как средство
экстренного повышения изменчивости.
Потомство, возникшее в результате полового размножения:
во-первых разнообразнее,
во-вторых не является точной копией
родителей
28. основные формы естественного отбора:
29. основные формы естественного отбора
Экспериментально показано:
если условия существования популяции резко
меняются, происходит не плавный "сдвиг"
организмов
в
нужную
сторону,
а
дестабилизация системы, проявляющаяся,
прежде всего, в резком росте изменчивости
Изменение адаптивной нормы
t0 - исходная норма. t1-t3 - стадии дестабилизации. t4-t7 на краю "расползшегося" купола формируется и
фиксируется новая адаптивная норма.
По горизонтальной оси - значение какого-либо
количественного признака, по вертикальной - число
особей с таким значением признака.
(Корочкин Л.И. Введение в генетику развития. М., 1999.)
Искуственный отбор в лаборатории
Эксперименты Джона Эндлера.
Характером пятнистости гуппий можно манипулировать с помощью
искусственного отбора.
крупный гравий+хищник мелкий гравий+хищник
менее 15 поколений
отбора
менее 15 поколений
отбора
крупный гравий без хищника мелкий гравий без хищника
менее 15 поколений
отбора
менее 15 поколений
отбора
Искусственный отбор
Брокколи подавление
развития цветков
Цветная капуста стерильность
цветков
Кочанная капуста подавление роста
междоузлий
Дикая капуста
Листовая капуста увеличение
листьев
Кольраби - усиление
латеральных
меристем
38. Искусственный отбор
39. Особенности пород домашних животных и сортов культурных растений
• 1) они представлены большим
многообразием;
• 2) породы и сорта одной группы отличаются
как между собой, так и от дикого предка;
• 3) они отвечают нуждам человека.
Ч. Дарвин Изменение животных и растений в одомашненном
состоянии (1868).
40. Columbia livia
41. Монофилия пород Columbia livia
• все они имеют сходные признаки в строении;
• все породы скрещиваются между собой и с
диким голубем и дают плодовитое потомство;
• в потомстве от таких скрещиваний часто
наблюдаются уклонения в сторону дикого
голубя;
• все породы имеют одинаковые повадки
(строительный инстинкт, вынашивание и
кормление птенцов и т.д.);
• все породы способны к одичанию, и при этом
наблюдается утрата породных признаков
42. Условия искусственного отбора:
• 1) усиление изменчивости (большое число
особей);
• 2) устранение нежелательных скрещиваний;
• 3) строгая методичность ведения селекции;
• 4) наблюдательность и опыт
действие искусственного отбора ограничено
во времени и численностью особей
Читайте также: