
Цикл развития мхов и папоротников кратко
Обновлено: 04.07.2024
При половом размножении постоянно создаются новые комбинации генов, что увеличивает генетическое разнообразие потомства и, соответственно, шансы приспособиться к меняющимся условиям среды. Создание новых комбинаций генов происходит в процессе мейоза. Мейоз имеет место в ходе образования гаплоидных клеток — спор или гамет. При этом хромосомы, доставшиеся организму от матери и отца, перестают существовать как целое — в результате кроссинговера образуются новые варианты хромосом, скомбинированные из отцовских и материнских. Таким образом, хромосомы детей, как правило, не идентичны хромосомам родителей — они содержат другие комбинации аллелей (вариантов генов).
В ходе мейоза происходит редукция хромосомного набора — образование из диплоидной клетки 4 гаплоидных клеток (n), у которых каждая хромосома представлена уже не парой гомологов, а 1 хромосомой. У человека путем мейоза образуются половые клетки (гаметы), каждая из них несет 23 хромосомы.
Диплоидность клеток в жизненном цикле восстанавливается при оплодотворении — слиянии гамет. При этом объединяются два разных хромосомных набора из двух разных геномов (генома матери и генома отца).

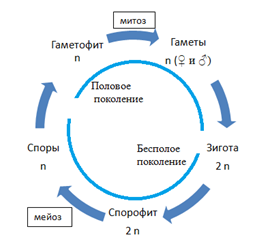
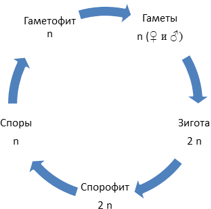
У других организмов соотношение диплоидной и гаплоидной фаз цикла может быть разным. Разные организмы демонстрируют большое разнообразие жизненных циклов, однако во всех них можно выделить диплоидную фазу, или диплофазу — от оплодотворения до мейоза — и гаплоидную фазу, или гаплофазу — от мейоза до следующего оплодотворения.

Например, у животных (слева) гаплоидны только гаметы, у растений (посередине) есть диплоидное поколение (спорофит) и гаплоидное поколение (гаметофит), а у многих одноклеточных преобладает гаплоидная фаза, а зигота после оплодотворения делится мейозом, в результате чего снова получаются гаплоидные клетки.
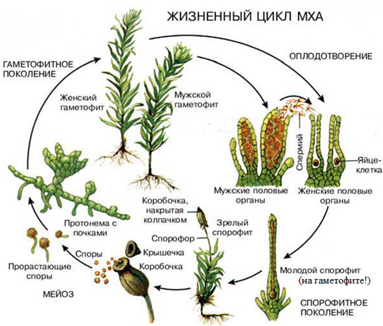
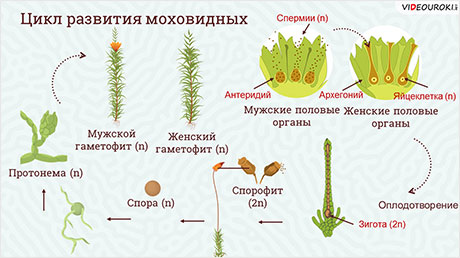
Соотношение гаплоидной и диплоидной стадии в жизненном цикле разных организмов может быть разным. Как мы уже видели, у животных единственной стадией является диплоидная, а гаплоидны только гаметы. У некоторых организмов, наоборот, единственной стадией является гаплоидная, а диплоидна только зигота, которая сразу после полового процесса вступает в мейоз. Такую ситуацию мы можем наблюдать, например, у хламидомонады. У большинства растений обе фазы представлены. У мхов привычные нам зеленые растения являются гаплоидными. На них формируются органы полового размножения: антеридии, в которых образуются сперматозоиды, и архегонии, в каждом из которых имеется одна яйцеклетка. Образующиеся гаметы после оплодотворения дают диплоидную стадию, которая развивается непосредственно на гаплоидной. Она представляет собой тонкую длинную нить с расширением на конце. Она коричневого цвета, т. к. не содержит хлорофилла, неспособна к фотосинтезу и живет за счет гаплоидной части растения. Внутри расширения, называемого коробочкой, множество клеток делится мейозом, образуя гаплоидные споры. Споры дают начало зеленым гаплоидным растениям. Таким образом, основной стадией у мхов является гаплоидная.
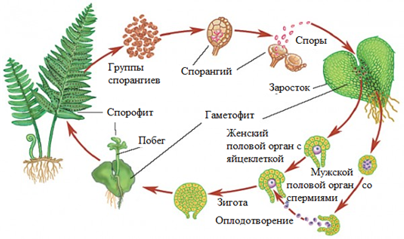
Основная жизненная стадия у папоротникообразных — диплоидная. На листьях папоротников или в специальных структурах у хвощей и плаунов образуются спорангии, в которых в результате мейоза формируются мелкие одноклеточные споры. Разлетаясь и попадая в подходящие условия, они дают начало гаплоидной стадии — заростку. На заростках образуются антеридии и архегонии, формируются гаметы, происходит оплодотворение, и из зиготы развивается диплоидное растение. У семенных растений самостоятельной гаплоидной стадии не существует, она представлена группами клеток, развивающихся в специальных органах диплоидных родительских организмов. Некоторое время отдельно существует только мужской гаметофит в виде пыльцевого зерна, но в нем в это время не происходит заметных процессов жизнедеятельности. После оплодотворения из зиготы новый диплоидный организм первые этапы развития также проходит внутри материнского организма.
Систематическое положение
Высшие споровые растения
Отдел: моховидные, или настоящие мхи
Класс: листостебельные, или настоящие, мхи
Класс: сфагновые мхи
Отдел: печеночные мхи
Отдел: антоцеротовые мхи
Бриология — наука о моховидных.
Основная характеристика отдела Моховидные
- Небольшие размеры.
- Влажные местообитания.
- Отсутствие корней, есть ризоиды.
- Отсутствие проводящей ткани.
- Для полового размножения необходимо присутствие капельножидкой влаги для передвижения сперматозоидов.
- В жизненном цикле чередование поколений с преобладанием гаплоидного гаметофита (!).
Отдел Моховидные
КЛАСС ЛИСТОСТЕБЕЛЬНЫЕ МХИ
Кукушкин лен (Рис. 1)

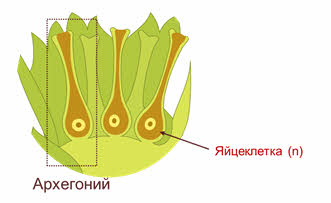
Широко распространен во влажных лесах умеренной зоны. Его коричневатый стебель, достигающий в длину 30 см, содержит мертвые вытянутые клетки, выполняющие функцию проведения и накопления воды. Простые листочки состоят из нескольких слоев зеленых клеток и не имеют жилок. В нижней части стебля молодого мха образуются одноклеточные выросты — ризоиды, выполняющие функцию поглощения воды и минеральных солей. Обычно кукушкин лен растет в виде куртин, в которых содержится несколько сотен особей. Зеленые растения мха — это гаплоидная стадия, или гаметофит. Мох кукушкин лен — раздельнополое, или двудомное, растение. На верхушке одних растений формируются женские органы, называемые архегониями (рис. 2).

Каждый архегоний содержит одну яйцеклетку. На мужских растениях развиваются антеридии — органы, представляющие собой мешочки, в которых образуются сперматозоиды. Так как растение гаплоидное, образование гамет не требует мейоза. Для оплодотворения мху нужна вода, по которой сперматозоиды могли бы переплыть на женское растение и проникнуть в архегоний. Для этого бывает достаточно дождя или обильной росы, капли которой лягут на верхушки растений. После оплодотворения из образовавшейся зиготы развивается диплоидный спорофит. Он растет на гаплоидном зеленом растении и использует образованные им вещества. Сам он не содержит хлорофилла и не фотосинтезирует. Он имеет вид длинной коричневой нити, называемой ножкой. Одним концом она прикреплена к верхушке гаметофита, а на другом образуется расширение, являющееся спорангием. В этом расширении формируются материнские клетки спор, которые делятся мейозом, образуя множество одноклеточных спор. Зрелый спорангий состоит из коробочки и крышечки. Крышечка отрывается, и споры высыпаются и разносятся ветром на большие расстояния. Из спор, попавших во влажное освещенное место, вырастает тонкая ветвящаяся нить (протонема). На ее концах формируются почки, дающие начало новым гаплоидным растениям.
КЛАСС СФАГНОВЫЕ МХИ
Мох Сфагнум
Важной группой мхов являются сфагновые мхи (рис. 3). Они широко распространены и образуют так называемые сфагновые, или верховые, болота. Верхушки сфагнума имеют светло-зеленые листочки. По мере роста мха нижняя его часть оказывается под толщей более молодых листочков, отмирает и начинает разлагаться. Ткани мха содержат карболовую кислоту, которая имеет мощное бактерицидное действие. Разложение мха происходит без доступа воздуха и в условиях почти полного отсутствия микроорганизмов. Мох практически не гниет и превращается в торф. Торф, как губка, удерживает влагу, происходит заболачивание лесов.

Сфагнум не имеет ризоидов. Он впитывает воду всей своей поверхностью. Мелкие светло-зеленые листья, покрывающие стебель и ветви, состоят из клеток двух хорошо различимых под микроскопом типов. Узкие зеленые фотосинтезирующие клетки образуют сетчатую структуру, в которой происходит движение органических веществ. Между ними находятся крупные прозрачные мертвые клетки, от которых остались только оболочки (рис. 4). Эти клетки служат резервуаром для воды. Через поры в их клеточных стенках происходит пополнение запасов воды из окружающего воздуха.

Сфагнум — однодомное растение. Архегонии (женские органы полового размножения) и антеридии (мужские органы полового размножения) располагаются на одном растении. Сперматозоиды (n) по каплям воды достигают яйцеклеток (n), происходит оплодотворение. Образуется зигота (2n), из которой вырастает спорангий. Там происходит мейоз и образуются гаплоидные (n) споры, дающие начала тонкой нити-предростку — протонеме (n). На ней образуется почка, прорастающая в новый гаметофит (n).
ОТРЯД ПЕЧЕНОЧНЫЕ МХИ
Небольшие нежные растения, тело которых представлено чаще всего слоевищем в виде пластинки неправильной формы. У некоторых слоевище несет чешуйки, соответствующие листьям. Есть многочисленные ризоиды. Протонема развита слабо.
Размножение половое (подобно другим мхам) и бесполое (с помощью особых почек).
Распространенный представитель печеночных мхов — маршанция (рис. 5).

ОТРЯД АНТОЦЕРОТОВЫЕ МХИ (РИС. 6)
Распространены большей частью в тропиках.
Спорофиты (спорогонии) рогообразной формы.
В основании ножки спорангия находится меристема, поэтому ножка постоянно растет. Образование и созревание спор происходит продолжительное время.
Большинство однодомны. Перекрестное опыление (женские и мужские гаметофиты созревают в разное время).
Значение мхов
- Образование торфа.
- Водный запас экосистем.
- Виды, образующие экосистемы.
- Содержат антибиотики.
Систематическое положение
Высшие споровые растения
Общая характеристика Папоротникообразных
- Возникли в девоне. Биологического расцвета достигли в карбоне, став основной лесообразующей группой. Остатки лесов образовали обширные залежи каменного угля.
- Наибольшее количество видов в тропической зоне.
- Предпочитают влажные местообитания, т. к. мужские гаметы подвижны, и влага нужна для передвижения сперматозоида к яйцеклетке.
- Есть ткани и органы.
- В жизненном цикле преобладает диплоидный спорофит.
- Размножение половое и бесполое (спорообразование).
- Органы полового размножения многоклеточные.
Отдел Папоротниковидные
В настоящее время отдел насчитывает примерно 12 000 видов.
Жизненные формы: травы, деревья (рис. 1) и лианы (несколько тропических видов). Есть водные формы (сальвиния плавающая (рис. 2)).


Листья могут быть дифференцированы на стерильные и фертильные (ужовник (рис. 4)), или выполнять одновременно обе функции (большинство папоротников (рис. 5)). У страусника фертильные листья не фотосинтезируют (рис. 6).



Рис. 3 Рис. 4 Рис. 5
Большинство папоротников имеют подземное корневище и хорошо развитые придаточные корни (рис. 7).


Жизненный цикл папоротников включает чередование гаплоидного гаметофита и диплоидного спорофита с преобладанием спорофита. В жизненном цикле происходит чередование полового и бесполого размножения (рис. 8).

При бесполом размножении на нижней стороне листа образуются парные выросты —сорусы (рис. 9). Сорус представляет собой ножку и покрывальце, закрывающее снизу шаровидные спорангии (рис. 10), отходящие от основания ножки. В спорангиях формируются материнские клетки спор, которые делятся мейозом с образованием гаплоидных клеток, становящихся спорами. В сухую погоду края покрывальца отгибаются, а оболочка спорангия лопается из-за неравномерного утолщения стенок образующих ее клеток.


Из спор, попавших во влажное освещенное место, развивается гаплоидный гаметофит папоротника — заросток — в виде сердцевидной пластинки с многочисленными ризоидами. На его нижней стороне образуются антеридии со сперматозоидами и архегонии с яйцеклетками. Так же как и мхам, папоротникам для оплодотворения нужна вода. По ней многожгутиковые сперматозоиды папоротника плывут к архегониям. Там сперматозоиды сливаются с яйцеклеткой, образуя диплоидную зиготу. Из нее вырастает новое диплоидное растение.
В лесах умеренной зоны наиболее часто встречаются щитовник мужской, кочедыжник женский, орляк.
Отдел плауновидные
- Побег стелющийся, дихотомически ветвящийся.
- Листочки (филлоиды) мелкие простые с одной центральной жилкой.
- Размножение половое и бесполое (спорообразование).
- В жизненном цикле преобладает диплоидный спорофит.
Представителем плаунов, часто встречающихся в нашей стране, является плаун булавовидный (рис. 11).

В жизненном цикле плаунов, как и всех папоротникообразных, происходит чередование полового и бесполого размножения (рис. 12). На концах побегов плауна образуются прямостоячие спороносные колоски — стробилы. Спороносные колоски покрыты видоизмененными чешуевидными листочками — спорофиллами — на которых образуются спорангии. В спорангиях в результате мейоза образуются гаплоидные споры. Созревшие споры высыпаются, и из них развивается гаплоидный заросток. У многих видов плаунов заросток развивается под землей в течение нескольких лет, питаясь гетеротрофно, в основном за счет симбиоза с грибом. На зрелом гаметофите образуются архегонии с яйцеклетками и антеридии со сперматозоидами. После оплодотворения из зиготы развивается диплоидный спорофит, который питается за счет гаметофита, пока не достигнет поверхности земли, где он начинает фотосинтезировать.

Отдел хвощевидные
Древняя группа сосудистых растений, представленная в настоящее время примерно 30 видами.
Стебли полые, состоящие из отдельных члеников и выполняющие функцию фотосинтеза (рис. 13). Для увеличения прочности под эпидермой проходят пучки волокон склеренхимы, образуя ребра на поверхности стебля. Кроме того, в стеблях хвощей откладываются мелкие кристаллы окиси кремния, увеличивающие их жесткость.
Под землей у хвоща образуется густая сеть корневищ, служащих для вегетативного размножения и переживания зимы.

Весной из-под земли вырастают спороносные побеги. Они имеют коричневый цвет, т. к. не содержат хлорофилла и живут за счет накопленных в прошлом году запасов питательных веществ. На их спорофиллах в результате мейоза образуются гаплоидные споры, которые имеют специальные нитевидные выросты, меняющие форму в зависимости от влажности. Это позволяет им легче покидать спорангий и шире распространяться. Они дают начало гаплоидному заростку. Жизненный цикл хвощей похож на жизненный цикл папоротников (рис. 14).
Циклы развития водорослей весьма многообразны, отличаются большой пластичностью и предопределяются многими экологическими факторами.
- Гаплофазный тип характеризуется отсутствием чередования поколений. Вся вегетативная жизнь водорослей проходит в гаплоидном состоянии, т.е. они являются гаплонтами. Диплоидна лишь зигота, прорастание которой сопровождается редукционным делением ядра (зиготическая редукция). Развивающиеся при этом водоросли оказываются гаплоидными. Примером являются многие зеленые (вольвоксовые, большинство хлорококковых, конъюгаты) и харовые водоросли.
- Диплофазный тип отличается тем, что вся вегетативная жизнь водорослей осуществляется в диплоидном состоянии, а гаплоидная фаза представлена только гаметами. Перед их образованием происходит редукционное деление ядра (гаметическая редукция). Зигота без деления ядра прорастает в диплоидный таллом. Эти водоросли являются диплонтами. Такой тип развития характерен для многих зеленых водорослей, имеющих сифоновую структуру, всех диатомовых и некоторых представителей бурых.
- Диплогаплофазный тип характеризуется тем, что в клетках диплоидных талломов (спорофитов) многих водорослей редукционное деление ядра предшествует образованию зоо- или апланоспор (спорическая редукция). Споры развиваются в гаплоидные организмы (гаметофиты), размножающиеся только половым путем. Оплодотворенная яйцеклетка — зигота — прорастает в диплоидный спорофит, несущий органы бесполого размножения. Таким образом, у этих водорослей имеет место чередование форм развития (генераций): диплоидного бесполого спорофита и гаплоидного полового гаметофита. Оба поколения по внешнему виду могут не различаться и занимать одинаковое место в цикле развития (изоморфная смена генераций) или же резко различаться по морфологическим признакам (гетероморфная смена генераций). Изоморфная смена генераций характерна для ряда зеленых (ульва, энтероморфа, кладофора), бурых и большинства красных водорослей.
Мхи по сравнению с другими высшими растениями наиболее примитивно организованы.

В отделе Моховидных развито половое поколение – гаметофит, который представляет из себя прежде всего взрослое растение мха. Бесполое же поколение (спорофит) представлен у мхов спорогоном (коробочка на ножке), который развивается на гаметофите после оплодотворения.
У низших мхов нет дифференцировки тела на вегетативные органы, и оно представляет собой плоскую листовидную пластинку – таллом, лежащую на почве или ином субстрате, прикрепляемую к ней тонкими ризоидами.
Развитие мха начинается со споры, т.е. с одноклеточного, микроскопически гаплоидного зачатка. После того, как спора попадает на влажный субстрат, из нее вырастает тонкая обычно ветвистая, зеленая нить или пластинку водорослей. Эта маленькая нить (пластинка) называется протонемой. Через некоторое время на протонеме появляются почки, дающие начало взрослому растению мха. У настоящих мхов стебель (каулидий) и листочки (филлидии) ясно отличаются друг от друга; стебель чаще всего в нижней части бывает покрыт волосками или ризоидами. На верхушках главных стеблей или боковых веточек развиваются половые органы: антеридии♂ архегонии ♀, в которых образуются половые клетки. Внутри антеридия развиваются сперматозоиды, архегоний же содержит в себе яйцеклетку. Все стадии развития мха, начиная со споры и кончая стеблем с листочками и половыми органами, объединяются в понятие полового поколения или гаметофита.
Оплодотворение яйцеклетки сперматозоидом совершается с помощью капель воды внутри архегония в сырую погоду; после оплодотворения на гаметофите вырастает коробочка со спорангием, в котором после редукционного деления образуются споры. Коробочка сидит на тонкой ножке. Это – спорогон мха или бесполое поколение (спорофит). К моменту созревания спор коробочка открывается наверху крышечкой и споры высыпаются.
Цикл развития папоротника.

Спорофит так называется взрослое листостебельное растение, которое образует значительные заросли в лесах умеренного пояса. Спорофит является преобладающим поколением этих растений. Следующим этапом цикла развития папоротника является созревание органов бесполого размножения. Они называются спорангиями. Эти структуры имеют вид небольших коричневых бугорков, расположенных на нижней стороне листьев. Сверху они дополнительно защищены пленчатыми "покрывальцами". Спорангии папоротников собраны в группы, которые называются сорусами. В конце лета эти структуры темнеют. Это значит, что спорангии созрели. Далее они раскрываются, и в землю высыпаются тысячи мелких клеток. Это и есть споры. При наличии достаточного количества влаги они сразу начинают прорастать. Результатом развития спор является заросток. Это и есть особь полового поколения, которая является следующим звеном в цикле развития папоротника. Внешне он представляет собой пластину зеленого цвета сердцевидной формы. Заросток развивается на почве, к которой прикрепляется с помощью ризоидов. По мере развития гаметофита на его нижней стороне формируются органы полового размножения. В них созревают половые клетки двух видов: яйцеклетки и сперматозоиды. Оплодотворение у папоротников имеет свои особенности. Во-первых, мужские и женские половые клетки на одном заростке созревают в разное время. Поэтому слияние гамет возможно только между разными растениями. Такой тип оплодотворения называется перекрестным. Второй особенностью этого процесса у папоротников является обязательное наличие воды. Дело в том, что половые клетки споровых растений не могут самостоятельно двигаться. Поэтому сперматозоид может добраться до яйцеклетки только с помощью воды. Таким образом, папоротники хоть и относятся к группе первых наземных растений, но не утратили связь с прежней средой обитания. Далее из оплодотворенной яйцеклетки развивается растение бесполого поколения, на нем созревают споры, и процесс повторяется.
Папоротники относятся к наиболее древним споровым растениям. Обитают в самых разнообразных экологических условиях: в заболоченных местах и водоемах, лесах тропического и умеренного климата. Наиболее известные представители - щитовник мужской, орляк, страусник. В жизненном цикле папоротников, хвощей, плаунов доминирует спорофит.
Расцвет папоротников произошел около 358 млн лет назад и длился примерно 65 млн лет, в честь папоротников назван период палеозойской эры - карбон, или каменноугольный период, который длился указанный промежуток времени. Именно папоротникам принадлежит активная роль в углеобразовании: в карбоне леса заселяли древовидные папоротники, достигавшие 40 и более метров в высоту.

Древесные формы папоротников сохранились до наших дней, однако большинство представителей - травянистые растения, у которых отсутствует камбий, а значит, нет и вторичной древесины.
Папоротники относятся к группе сосудистых растений, так как имеют жилки - сосудисто-волокнистые пучки, в отличие от мхов, которые не имеют жилок и не являются сосудистыми растениями. Механическая прочность обеспечивается отложением склеренхимы вокруг проводящих пучков (жилок).

В отличие от мхов, папоротники имеют проводящую ткань в стеблях и корнях, состоящую из ксилемы и флоэмы. Заметьте, я написал "корень" - у мхов не было и корней, вместо них были ризоиды, выполняющие аналогичную корням функцию. Корни папоротников, хвощей и плаунов всегда придаточные и растут от видоизмененного побега - корневища.
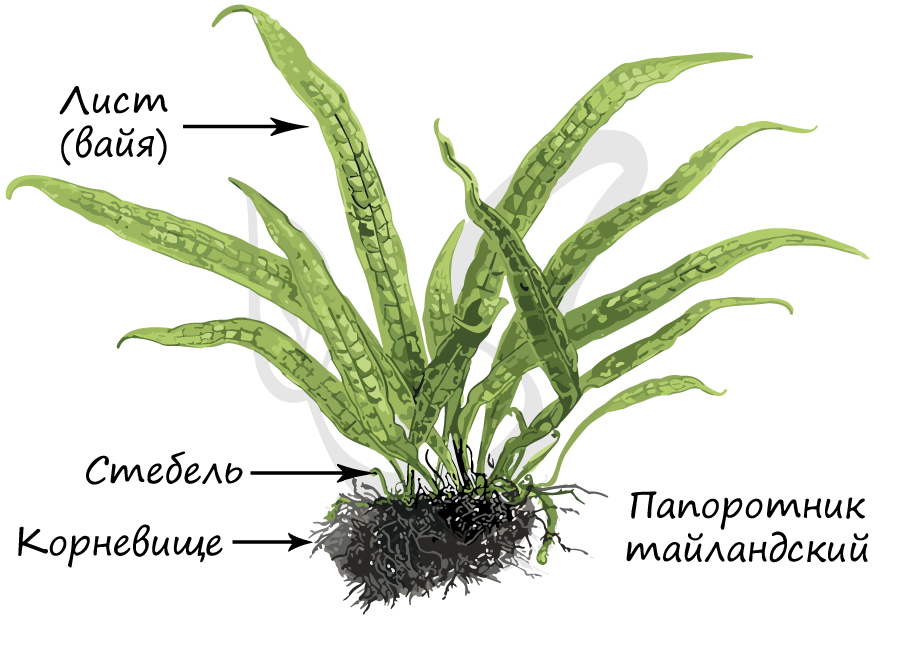
Строение
Рассмотрим строение папоротников на примере типичного представителя - щитовника мужского. Это широко распространенный папоротник, типичный для климата умеренных широт. Представляет собой многолетнее травянистое корневищное растение.
Образован пучком сильно расчлененных листьев, отходящих от корневища. Нарастают листья верхушкой, образуя завитки - "улитки".

Отметьте, что лист папоротника называется вайя (от греч. baion - пальмовая ветвь). В отличие от настоящих листьев, вайя имеет недетерминированный верхушечный рост. Листья имеют черешок, прикрепляющийся к стеблю, который может продолжаться в рахис - главную ось сложного листа, соответствующую центральной жилке.

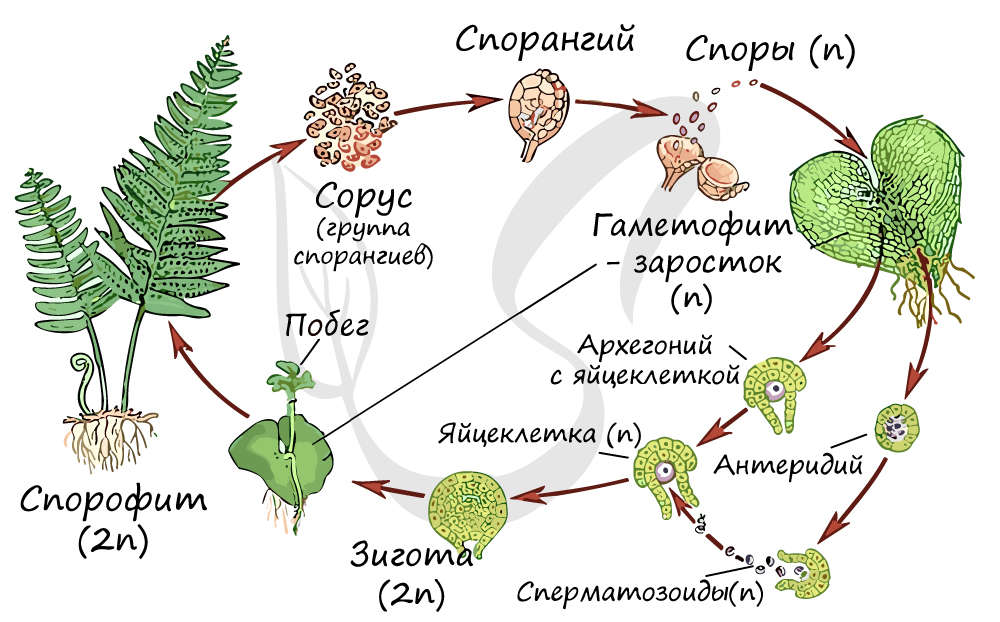
Жизненный цикл папоротниковидных
Листостебельное растение папоротника, изображенное выше - спорофит (2n). Спорофит доминирует в жизненном цикле папоротниковидных, в отличие от цикла мха, где спорофит, по сути, является придатком гаметофита (редуцирован). На нижней стороне вайи располагаются спорангии, собирающиеся в сорусы - группы близкорасположенных спорангиев. На спорофите (2n) в спорангии после мейоза образуются споры (n).

В 2012 году группа ученых под руководством Ксавьера Нобли из Университета Ниццы выяснили, что спорангий имеет специальный механизм "катапульты", споры из него вылетают со скоростью около 10 м/с.
Гаплоидные споры (n) прорастают в заросток (n), небольшую пластинку (несколько мм) сердцевидной формы. Заросток зеленого цвета, способен фотосинтезировать и прикрепляется к почве ризоидами. На нем образуются мужские и женские половые органы - соответственно антеридии и архегонии. Сперматозоид (n), образовавшийся в антеридии, благодаря воде (во время дождя) попадает в архегоний, где сливается с яйцеклеткой (n) и образуется зигота (2n).
Из зиготы развивается зародыш, который проникает в ткани архегонии с помощью особого приспособления - гаустории (от лат. haustor - черпающий, пьющий). Гаустория представляет собой ножку, внедряющуюся в ткани заростка и поглощающую из него питательные вещества. Начинается бурный рост зародыша, образуется побег, а затем взрослое растение - спорофит (2n). Цикл замыкается.
Значение папоротников
Папоротники являются главным компонентом многих лесных сообществ, звеном в цепи питания - продуцентами (производителями органических веществ). Человек использует папоротник в декоративных целях. Молодые побеги некоторых папоротников съедобны и употребляются в пищу: побеги орляка, листья страусника обыкновенного.
Щитовник мужской имеет медицинское значение: из его корневищ изготавливают противогельминтный препарат.
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
47.387. Царство Растения (мхи, папоротники, голосеменные)
Общая схема жизненного цикла высших растений
Отдел Моховидные
Как мы уже сказали, мхи – тупиковая ветвь в развитии растений на Земле. Они являются одними из самых примитивных растений, однако распространены достаточно широко в регионах с влажными почвами. Зачем им нужна влага? Сейчас мы ответим на этот вопрос.
У этих растений отсутствуют корни. Функцию прикрепления к субстрату и всасывания необходимых веществ выполняют ризоиды (выросты эпидермиса). Тело мхов часто (однако не всегда) разделено на стебель и листья. Ткани у мхов развиты гораздо меньше, чем у других высших растений, а ксилема и флоэма (компоненты проводящей ткани) в принципе отсутствуют.
Ниже представлено изображение одного из самых знаменитых представителей моховидных – кукушкиного льна. Цифрой 1 обозначено мужское растение, 2 – женское, под цифрой 3 скрыта коробочка, которая несет в себе споры.
Спорофит мхов представлен коробочкой на ножке, которая внедряется в ткань гаметофита (взрослого мха) и доставляет спорофиту воду и питательные вещества. Таким образом, спорофит мха полностью зависим от гаметофита и не способен к самостоятельному существованию. На спорофите образуются споры, которые способны прорастать лишь в благоприятных условиях. Из них развивается протонема (зеленая нить), которая дает начало гаметофиту.
Важно отметить, что для размножения мхам необходима вода. Их сперматозоиды переносятся к яйцеклеткам только в капельно-жидкой среде. Именно этот факт ограничивает распространение мхов (и других споровых растений) по Земле.
Экология моховидных
Моховидные - одни из первых наземных растений, мало изменившиеся за время своего существования. Они обитают во влажных местах, часто под пологом леса, а также на болотах и на полях с кислыми почвами. В настоящее время в этом отделе выделяют около 15 тысяч видов.
Они способны впитывать и удерживать большое количество влаги, и поэтому способствуют поддержанию водного режима местности. Также они часто обеспечивают условия для сохранения и прорастания спор голо- и покрытосеменных растений.
Способность мхов всасывать и удерживать в себе жидкость ранее использовалась при наложении гемостатических повязок в случае отсутствия более надежных средств.
Сфагнум – один из самых известных представителей отдела – не гниет в толще отложений, поэтому в течение многих лет участвует в формировании торфа, который широко используется в качестве топлива и источника органических удобрений.
Также из мхов выделяют некоторые химические вещества, используемые как в домашнем хозяйстве, так и в промышленности.

Папоротниковидные
Папоротниковидные – более развитые споровые растения по сравнению со мхами. Этот отдел растений возник очень давно, еще в период Девона. К настоящему виду отдел насчитывает около 12 тысяч видов. Папоротники широко распространены во влажных тропических лесах, где до сих пор встречаются их древовидные формы.
Как и мхи, папоротники чувствительны к наличию влаги в окружающей среде, так как от нее зависит их размножение и распространение.

На обратной стороне листа расположены сорусы – скопления спорангиев.

В жизненном цикле папоротников преобладает спорофит. Он представлен взрослым растением, которое мы можем видеть в лесах. Гаплоидные споры у папоротников формируются на обратной стороне листа и распространяются на большие территории при помощи ветра, из них в благоприятных условиях вырастает гаметофит – заросток. Это небольшое (обычно около полусантиметра) зеленое растение, живущее недолго и формирующее гаметы. Заросток часто имеет сердцевидную форму. Гаметофиты обычно двуполые. При слиянии гамет (в водной среде!) формируется новое растение – диплоидный спорофит, развивающийся из зиготы.
Экология папоротниковидных
Папоротники, как и мхи, способны существовать в основном только во влажной среде обитания, это ограничивает их распространение по земному шару. Однако число видов этих растений достаточно велико.
Папоротники являются важными компонентами многих растительных сообществ, особенно в тропических и субтропических лесах.
Человек использует папоротники в различных сферах: некоторые папоротники выращиваются в декоративных целях, некоторые употребляются в пищу, из отдельных видов получают экстракты лекарственных веществ.
Голосеменные
Голосеменные – потомки древних папоротниковидных, впервые возникшие около 350 млн лет назад (в конце девонского периода). Своего расцвета эта группа растений достигла в мезозойскую эру. Этот период был связан с образованием гор, поднятием материков над уровнем мирового океана и, следовательно, иссушением климата. Позже, с приходом покрытосеменных, голосеменные утратили свое господство.
Ранее в отделе выделяли 6 классов, однако 2 из них (семенные папоротники и бенннетиговые) полностью вымерли. В настоящее время класс включает в себя всего 900 видов, но, несмотря на малое разнообразие, эти виды очень широко распространены по Земле. В частности, в умеренных широтах Северного полушария они формируют огромные хвойные леса – тайгу.

Внешний вид голосеменных достаточно характерен: в основном это деревья, реже – кустарники. Листья у них имеют специфическую форму – в основном они представлены иглами, расположенными попарно и поодиночке, эти листья называются хвоей. Однако иногда встречаются и чешуевидные листья, значительно различающиеся по форме и размерам.
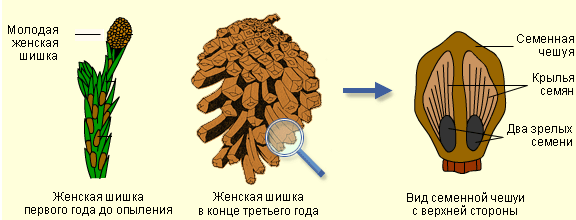
Важным шагом в эволюции семенных растений стало появление возможности размножаться при помощи семян, которые развиваются из семязачатков. У голосеменных семязачатки расположены открыто (голо) на семенных чешуях. Семенные чешуи обычно собраны в шишки, которые бывают мужскими и женскими. Хотим обратить ваше внимание, что хоть в шишках и происходит оплодотворение, они являются частью спорофита. Гаметофит, образующийся на семенных чешуях (из споры), настолько редуцирован, что не различим глазом. Редукция гаметофита также привела к тому, что для развития мужских гамет не нужны половые органы, поэтому антеридий (мужской половой орган) у голосеменных также редуцирован.
Перед вами строение молодой и зрелой женской шишки, а также внешний вид зрелого семени, расположенного на поверхности семенной чешуи.
На курсе нам предстоит изучить особенности цикла развития голосеменных, более сложного, чем циклы споровых растений.
Экология голосеменных
Голосеменным для размножения не нужна вода, поэтому их среда обитания более разнообразна по сравнению со споровыми растениями. Некоторые виды встречаются даже в пустынях, существуют тропические виды, но наибольшего распространения получила группа растений из класса Хвойные.
По своему значению в биосфере и в хозяйственной деятельности человека Хвойные уступают только Покрытосеменным. В настоящее время насчитывается более 500 видов хвойных растений, которые образуют леса на обширных территориях Северной Евразии и Северной Америки, также они распространены вдоль побережья Тихого океана.
Хвойные леса – среда обитания для многих животных из самых разных систематических групп. Деревья дают им пищу, защиту, являются местом для выведения и выращивания потомства. Залежи каменного угля, занимающие обширные территории Сибири, образовались в результате жизнедеятельности древних голосеменных.
Человек активно использует хвойные растения в своей хозяйственной деятельности. В частности, он заготавливает древесину, получает сырье для изготовления канифоли, скипидара, различных эфирных масел для парфюмерной промышленности, использует хвою в процессе заготовки лекарственных препаратов, в пищу идут семена некоторых хвойных – кедра, сибирской, итальянской сосен, из этих семян также делают масло. Некоторые Хвойные выращиваются как декоративные.
Такое активное использование лесов ведет к стремительному уменьшению их количества. Истреблению Хвойных также способствует строительство железных и шоссейных дорог, промышленных предприятий, городов, добыча полезных ископаемых (угля, нефти). Хвойные очень плохо переносят загазованность воздуха, копоть: многолетние хвоинки накапливают в себе ядовитые вещества, из-за чего отмирают раньше положенного им срока на 1-2 года. Также эти деревья очень чувствительны к пожарам, даже низовым (когда горит только подложка – мех, кустарники, а сами деревья не затрагиваются), так как их корневая система расположена в значительной мере поверхностно. Угнетение Хвойных растений – одна из серьезных экологических проблем современности.
Хуже всего освоена тема размножения растений и чередования поколений спорофита и гаметофита, поэтому на ней мы остановимся более подробно.
Развитие растений
Самое сложное для понимания, и в тоже время ключ для решения проблемы, заключено в представленной ниже схеме:

Основные понятия по данной теме:
Гаметофит – заросток, гаплоидное половое поколение в цикле развития растения, дающее споры. У споровых растений заростки формируются из спор, выпадающих из спорангия на почву, и живут самостоятельно, на них имеются архегонии и антеридии. Обычно эти заростки обоеполые. У семенных растений споры не выпадают из спорангиев и раздельнополые заростки развиваются внутри них. У голосеменных женский заросток – первичный эндосперм с двумя архегониями, у покрытосеменных – восьмиядерный зародышевый мешок (без архегониев). Мужской заросток у семенных растений – пыльца (не имеет антеридиев).
Оплодотворение – слияние сперматозоида (спермия) с яйцеклеткой. У споровых растений для оплодотворения нужна воды, которая переносит сперматозоиды, у семенных растений спермии (обычно их два) доставляются по пыльцевой трубке. У голосеменных в оплодотворении принимает участие один из спермиев. Для покрытосеменных характерно двойное оплодотворение.
Семя – орган размножения и расселения голосеменных и покрытосеменных. Образуется из семязачатка в результате оплодотворения. Состоит из кожуры, зародыша и запасающей ткани – эндосперма.
Семязачаток – видоизмененный мегаспорангий семенных растений, в котором происходит образование мегаспор, развитие женскоро гаметофита и оплодотворение, в результате чего образуется семя. Семязачатки голосеменных открыто лежат на чешуе шишек, у покрытосеменных – внутри завязи.
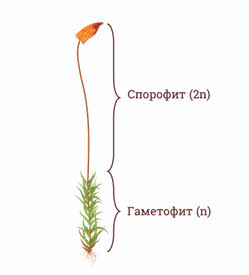
Спорофит – бесполое диплоидное поколение в цикле развития растения, которое чередуется с гаплоидным поколением – гаметофитом. Спорофит развивается из зиготы, на нем образуются спорангии со спорами. У бурых водорослей, папоротников, хвощей, плаунов спорофит и гаметофит представляют собой разные особи, при этом гаметофит микроскопически мал. У мхов оба эти поколения находятся на одной особи и гаметофитявляется листостебельным растением, а спорофит – коробочка на ножке. У голо- и покрытосеменных оба поколения также находятся на одном растении, но преобладает спорофит, а гаметофиты редуцированы и микроскопически малы.
Запомните, что споры образуются путем мейоза, а гаметы – путем митоза. Это характерно не только для растений, но и для некоторых простейших, имеющих сходство с растениями (например, для хламидомонады).
Чтобы окончательно разобраться в циклах развития растений, стоит посмотреть на следующие схемы:

Видеоурок знакомит учащихся со споровыми и семенными растениями, формирует общее представление об усложнении тела растений, строении и размножении печёночных и листостебельных мхов как представителей высших споровых растений. Во второй части видеоурока рассматривается жизненный цикл папоротникообразных, показано сходство жизненного цикла папоротников с жизненным циклом остальных споровых растений (плауновидные и хвощовые). Видеоурок способствует формированию навыков анализа, сравнения, нахождения черт сходства и различия на примере основных групп высших споровых растений.
В данный момент вы не можете посмотреть или раздать видеоурок ученикам
Чтобы получить доступ к этому и другим видеоурокам комплекта, вам нужно добавить его в личный кабинет, приобретя в каталоге.
Получите невероятные возможности



Конспект урока "Размножение мохообразных и папоротникообразных растений"
Высшие растения подразделяют на споровые и семенные.
К споровым относятся отделы Мохообразные и Папоротникообразные. Отдел Папоротникообразные включает классы: Папоротниковидные, Плауновидные и Хвощовые, или Клинолистовидные, Членистые.
Также вспомним, что когда организмы могут размножаться как бесполым, так и половым путём, то говорят о бесполом и половом поколениях данного вида. Если они закономерно сменяют друг друга, то такое явление называется чередованием поколений.
Границами, разделяющими бесполое и половое поколения в цикле развития растений, являются процесс образования спор с гаплоидным набором хромосом и оплодотворение, в результате которого восстанавливается диплоидный набор хромосом.
У растений выделяют диплоидное поколение – бесполое, или спорофит, и гаплоидное поколение – половое, или гаметофит.
Рассмотрим размножение представителей отдела Мохообразные.
Мохообразные – это единственный отдел высших растений, у которых гаметофит является преобладающим поколением. Растение, которое мы представляем себе как мох, и является гаметофитом. Мохообразные отличаются относительно простым строением.
Развитие мохообразных представляет собой чередование гаплоидной и диплоидной фаз. Гаметофит развивается из споры, ядро которой имеет гаплоидный набор хромосом, поэтому все клетки гаметофита тоже гаплоидные.
Первоначально из споры развивается протонема, имеющая вид ветвящейся нитчатой водоросли или пластинки. Протонема даёт начало половому поколению (гаметофиту). Спора не содержит запаса питательных веществ, поэтому молодой гаметофит должен их синтезировать самостоятельно посредством фотосинтеза. Наличие необходимой для этого хлоренхимы определяет зелёную окраску гаметофита.
У печёночных мхов многократное деление верхушечной клетки даёт начало пластинчатым структурам, на которых впоследствии разовьются половые органы.
У листостебельных мхов гаметофит имеет вид облиственных побегов, они развиваются из почек, которые формируются на протонеме. На гаметофорах образуются половые органы – женские архегонии и мужские антеридии.
Антеридии представляют собой овальные тельца, окружённые тонкой однослойной оболочкой.
Они наполнены сперматогенными клетками, которые при митотическом делении дают начало двум подвижным сперматозоидам, снабжённым двумя жгутиками.
Архегоний устроен сложнее и обычно представляет собой структуру, по своей форме напоминающую бутылку. В утолщённой части, называемой брюшком, находится крупная яйцеклетка, которая также образуется в результате митоза. Внутри суженной шейки в один ряд располагаются шейковые клетки, одна из которых – брюшная шейковая клетка – находится над яйцеклеткой.
Оплодотворение возможно только в капельно-жидкой водной среде. Через шейку сперматозоиды проникают в брюшко архегония и оплодотворяют находящуюся там яйцеклетку. В результате образуется диплоидная зигота, которая по прошествии определённого периода покоя даёт начало диплоидному поколению – спорофиту.
Внутри коробочки многочисленные материнские клетки спор делятся мейотически и дают начало гаплоидным спорам, посредством которых осуществляется бесполое размножение.
В коробочке печёночных мхов среди спор находятся пружинки – элатеры. При повышении влажности элатеры раскручиваются, а при понижении вновь скручиваются, при этом споры перемешиваются и разрыхляются, что способствует выталкиванию их из коробочки после того, как она раскрывается.
Споры, выпадающие из коробочки, подхватываются ветром и переносятся на большие расстояния. Подавляющее большинство погибает, попадая в неблагоприятную среду, но количество их столь велико, что часть обязательно попадает во влажную почву, где из них прорастает протонема, дающая начало молодому гаметофиту.
У мохообразных также широко распространено вегетативное размножение. У печеночников образуются выводковые тельца и выводковые корзиночки.
А у листостебельных мхов из структур гаметофита могут образовываться вторичные протонемы.
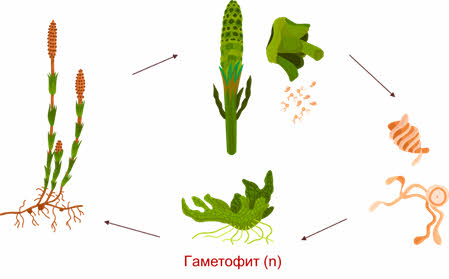
Рассмотрим размножение представителей отдела Папоротникообразные.
В наше время живут различные представители трёх основных классов этого отдела: папоротниковидные, хвощовые или клинолистовидные (членистые), и плауновидные.
У папоротникообразных преобладающим поколением является спорофит, его строение значительно сложнее, чем у мохообразных.
Спорофиты папоротникообразных имеют прекрасно развитую проводящую систему. Гаметофит, напротив, в значительной мере упростился.
Большая часть папоротников – это равноспоровые растения, но имеются и разноспоровые (например, сальвиниевые).
Споры имеют типичное строение. Их покровы образуют две оболочки: внутреннюю – интину и наружную – экзину. Большое количество спор рассеиваются, но лишь небольшая часть спор попадает в благоприятные условия, и после определённого периода покоя они прорастают.
Из споры прорастает гаметофит. Гаметофит имеет вид уплощённой зелёной пластинки, часто сердцевидной формы. Обычно гаметофит очень маленький – всего около 0,5 см в диаметре. На обращённой к субстрату поверхности гаметофита образуются многочисленные нитевидные ризоиды, которые связывают его с субстратом и способствуют поглощению воды.
Как правило, на одном гаметофите имеются антеридии и архегонии, то есть гаметофит однодомный. У большинства папоротников мужские органы возникают несколько раньше женских. Количество половых органов, образующихся на одном гаметофите папоротниковидных, относительно невелико.
Оплодотворение папоротниковидных осуществляется только в капельно-жидкой водной среде. Подвижные сперматозоиды, снабжённые многочисленными жгутиками, выходят из антеридиев и, привлекаемые веществами, которые выделяют архегонии, подплывают к ним, проникают через шейку в брюшко и сливаются с находящейся там яйцеклеткой. В результате образуется зигота с диплоидным набором хромосом. Несмотря на количество оплодотворённых яйцеклеток, лишь одна зигота развивается и даёт начало многоклеточному зародышу.
Остальные погибают и не мешают развитию молодого спорофита, который первоначально нуждается в значительном количестве органического материала. Вскоре у растущего зародыша образуются корни и фотосинтезирующие органы, после чего он способен самостоятельно жить независимом от материнского гаметофита. Сам гаметофит вскоре после этого отмирает.
Размножение представителей класса Плауновидные
Плаун представляет собой вечнозелёное многолетнее растение, у которого имеется стелющийся по земле длинный стебель. На этом ползучем стебле данного травянистого растения расположены многочисленные мелкие листья.
Листья плауновидных отличаются от листьев всех других высших растений, так как они представляют собой выросты стебля.
Листья могут быть вегетативными – (трофофиллы) или спороносными – (спорофиллы). Спорофиллы образуют скопления в виде спороносных зон, собранных в средней части стебля, или стробил, на концевых участках веточек.

Плауновые являются равноспоровыми. Одиночные спорангии развиваются в пазухах спорофиллов (спороносных листьев), которые собраны в стробилы.
Развитие спорангия и созревание продолжается от нескольких месяцев до нескольких лет. Зрелый спорангий раскрывается в сухую погоду.
Спорангии разноспоровых плауновидных различные. Крупные мегаспорангии развиваются на мегаспорофиллах, мелкие микроспорангии – на микроспорофиллах. При этом оба типа спорофиллов развиваются на одном и том же растении, будучи собранными в стробилы.
Размеры спор колеблются от 20 до 100 мкм – микроспоры, от 100 до 1500 мкм – мегаспоры. Количество микроспор всегда очень велико – до миллиона в спорангии. Количество мегаспор гораздо меньше: десятки или сотни, у некоторых видов в мегаспорангии развивается одна мегаспора. У многих разноспоровых из спорангиев выходят не споры, а женские и мужские гаметофиты.
У равно- и разноспоровых плауновидных дальнейшее развитие споры происходит различно.
У равноспоровых возможны два варианта прорастания спор: у одних спора прорастает через несколько дней после очень короткого периода покоя, у других спора долго находится в состоянии покоя и прорастает лишь через несколько лет. Споры постепенно погружаются в субстрат на несколько сантиметров.
Из-за отсутствия под землёй света гаметофит лишён возможности осуществлять фотосинтез, и он является сапрофитом, вступая в симбиоз с почвенными грибами. Гаметофит растёт до 15 лет. На верхней стороне гаметофита развиваются половые органы – однодомные заростки.
В результате оплодотворения в капельно-жидкой водной среде образуется диплоидная зигота. Период покоя отсутствует, и зигота начинает делиться. Наземная часть молодого спорофита зеленеет, в ней постепенно накапливается большое количество фотосинтезирующей ткани, достаточное для самостоятельного существования.
Иначе прорастают мега- и микроспоры всех папоротникообразных.
Гаметофит разноспоровых плауновидных наиболее упрощённый. Мужской гаметофит остаётся в микроспоре. Женский гаметофит развивается внутри мегаспоры благодаря наличию в ней большого количества питательных веществ, за счёт которых гаметофит растёт и развивается.
Чаще всего мегаспоры выходят из мегаспорангия, но у многих видов гаметофит может развиваться при нахождении мегаспоры в спорангии.
У разноспоровых плауновидных развитие зародыша значительно варьирует. Полностью сформированный зародыш состоит из стебелька с верхушечной точкой роста, двух листочков, корешка и ножки, с помощью которой зародыш получает из гаметофита все необходимые вещества. Со временем корешок и листочек прорывают ткань гаметофита, выходят наружу и, достигнув достаточного уровня развития, снабжают спорофит водой с минеральными веществами и продуктами фотосинтеза. После этого заросток постепенно отмирает.
Размножение представителей класса Хвощовые, или Клинолистовидные, Членистые
Самый малочисленный класс папоротникообразных – это хвощовые.
Все современные хвощовые являются равноспоровыми растениями. Их спорангии развиваются на особых, присущих только хвощовым, структурах – спорангиофорах.
Побеги, на которых они образуются, у большинства видов наряду со спорогенной функцией обладают и фотосинтетической. Такие вечнозелёные и многолетние побеги имеют очень высокую механическую прочность. У других видов (например, хвощ лесной, хвощ полевой и др.) зелёные однолетние побеги двух типов – чисто вегетативные и спорогенные.
На спорогенных вначале образуются стробилы, но после созревания спор и опадания стробилов эти побеги нельзя отличить от обычных вегетативных.
Стробилы хвощей образуются на верхушке главного побега. Прорастание споры начинается с деления её ядра.
Первоначально молодой гаметофит напоминает нитчатую водоросль, но со временем он становится сложно устроенным пластинчатым (заростком) с многочисленными уплощёнными зелёными выростами, направленными вверх.
На нижней поверхности заростка формируются бесцветные нитевидные ризоиды, снабжающие растение водой и минеральными веществами. В зависимости от условий развития размеры заростков варьируют от миллиметра до нескольких сантиметров.
У одних растений гаметофит двудомный, у других – однодомный. Мужские гаметофиты меньше женских. Оплодотворение происходит в капельно-жидкой водной среде. Зигота даёт начало зародышу, который состоит из корешка, стебелька и двух-трёх листочков. Вначале зародыш окружён тканью гаметофита, по мере развития корешок прободает её и попадает в почву, где начинает активно выполнять свои функции. Зародышевый корешок живёт недолго. Из первичного побега сначала образуется корневище, от узлов которого отходят придаточные корни, а затем надземные побеги.
Читайте также: