
Биологический этап эволюции кратко
Обновлено: 04.07.2024
В историческом развитии органического мира четко прослеживается наличие целого ряда этапов, свидетельствующих об усложнении и совершенствовании свойств организмов. Они наблюдаются уже на самых ранних этапах существования живого. Важнейшим из них является появление многообразия прокариот и эукариот.
Роль прокариот в эволюции биосферы
Появление автотрофов среди прокариот — крупное событие в эволюции живого мира.
Среди автотрофов — организмов, самостоятельно образующих органическое вещество из неорганических соединений, различают фототрофов, создающих органические соединения за счет энергии Солнца, и хемотрофов, живущих за счет энергии, высвобождающейся при химических реакциях окисления неорганических соединений (молекулярного водорода, окиси углерода, соединений серы или азота и др.).
Ученые полагают, что процесс появления хемосинтезирующих и фотосинтезирующих автотрофов начался еще на этапе коацерватных капель, когда их содержимое все более усложнялось. Происходившие при этом случайные мутации нуклеиновых кислот могли дать некоторые молекулы, по строению близкие к хлорофиллу, способные использовать энергию света.
Пока питательные вещества в окружающей водной среде были в избытке, это свойство не давало преимуществ в существовании организмам, имеющим активные пигменты. Но по мере увеличения численности организмов и сокращения питательных веществ в окружающей среде способность усваивать CO2 при участии энергии света оказалась полезной для автотрофов и их потомства, обеспечивая их выживание в конкурентной борьбе за существование.
Особенно важным для развития живой природы оказалось появление хлорофилла — зеленого пигмента, способного улавливать энергию солнечного света и осуществлять фотосинтез. Появление фотосинтеза произошло примерно 3,9 млрд лет назад. Оно стало крупнейшим прогрессивным событием в эволюции живого мира.
Роль эукариот в эволюции биосферы

С появлением эукариот началось становление и развитие новых крупных групп организмов — царств растений, животных и грибов. Пышный расцвет эукариотных форм жизни привел к возникновению в органическом мире таких крупных ароморфозов, как многоклеточность и половое размножение, обеспечивающее развитие организмов из зиготы.
Отпечатки червей на выветренной поверхности известняка (1200-600 млн лет назад, протерозой)
Развитие организма из зиготы, которая образуется при слиянии половых клеток двух родительских организмов, послужило началом регулярного появления и закрепления новых качеств у дочерних организмов, что позволяло последующим поколениям лучше приспосабливаться к жизни в изменяющихся условиях среды. После этого события, произошедшего около 1,9 млрд лет назад, заметно ускорились процессы прогрессивного развития живого.
Благодаря происходящим физико-химическим изменениям в биосфере формировалась атмосфера, в которой начал накапливаться свободный кислород. Вскоре произошла так называемая кислородная революция: установилась достаточно устойчивая концентрация свободного кислорода — 1% от нынешнего, современного, количества. Накопление свободного кислорода привело к возникновению первичного озонового экрана в верхних слоях биосферы, что обусловило ускорение развития жизни. Формирование озонового слоя началось в конце протерозоя (1200-600 млн лет назад).
Появление фотосинтезирующих организмов, особенно эукариот — водорослей, ускорило накопление свободного кислорода в атмосфере. Уже на границе силура и девона содержание свободного кислорода в атмосфере достигло 10% от современного уровня, а к концу палеозоя (около 250 млн лет назад), в пермском периоде, — приблизительно той же концентрации, которая наблюдается и в наше время.
Формы наземной жизни
Важным событием в эволюции органического мира был выход живых организмов на сушу. Первыми это сделали бактерии и цианобактерии. Ученые полагают, что это произошло около 3,5-3,2 млрд лет назад, то есть задолго до появления эукариот. Выйдя на обмелевшие участки суши, прокариоты начали процесс образования почвы. Спустя большой период времени на сушу вышли и эукариоты — растения, животные и грибы. Это произошло около 500-450 млн лет назад. С этих пор биологическая эволюция шла не только в водной, но и в наземно-воздушной среде.
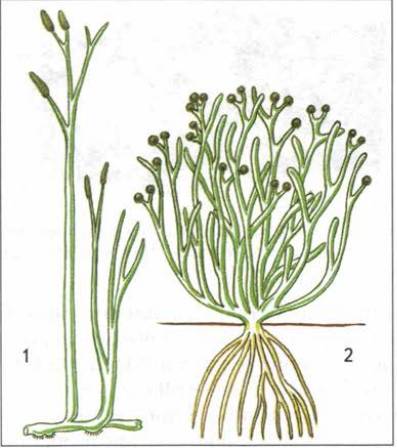
Первые растения, вышедшие на сушу, поселились на влажных прибрежных участках вдоль пресных водоемов. Это были теперь уже давно вымершие многоклеточные растения — риниофиты, произошедшие от зеленых многоклеточных водорослей. Почти одновременно с растениями на сушу вышли и первые животные — ракоскорпионы из паукообразных.
Первые наземные растительные организмы: 1 — риния; 2 — куксония
На суше организмы столкнулись с обилием света, его суточным и сезонным ритмом яркости и продолжительности. Это обусловило появление организмов, ведущих ночной или дневной образ жизни. При этом у многих видов наблюдалась выработка совместных, сопряженных друг с другом ритмов развития. Для лучшего улавливания света у растений развились листья. Эти и многие другие черты приспособленности появились у организмов в процессе эволюции в связи с тем, что они вышли из водной среды в новые разнообразные условия наземно-воздушной среды.
Выход растений и животных на сушу произошел сравнительно недавно в истории Земли. Но к этому времени в водах Мирового океана и пресных водоемах жизнь уже достигла достаточно высокого уровня развития. За многие миллионы лет путем длительной биологической эволюции здесь появились разнообразные бактерии, цианобактерии, простейшие, многоклеточные животные, растения и грибы. Значительная часть их вымерла, но многие группы древних организмов или производные от них существуют и в наше время.
Как в водной среде, так и на суше жизнь различных организмов протекала совместно — в сообществах (биогеоценозах). Распространяясь по земной поверхности, сообщества живых организмов все более и более меняли ее облик и создавали особые условия жизни на этих территориях. С появлением высокорослых растений и разнообразных животных образовывались такие природные сообщества, которые не только по горизонтали, но и по вертикали многометровым слоем жизни охватывали поверхность суши. При этом различные виды, обитая в сообществах рядом друг с другом, в процессе эволюции вырабатывали разные приспособительные свойства к совместной жизни.
Совместная жизнь различных видов в природных сообществах, появившаяся в биосфере еще на заре развития живого мира, является чрезвычайно важным фактором биологической эволюции, совершающейся на Земле.

Первые живые клетки уже обладали важнейшим свойством каждого живого организма — способностью к точному самовоспроизведению, самокопированию.
Питались они готовыми органическими веществами, которые образовались на ранних этапах формирования Земли абиогенным путем. По мнению большинства ученых, в период появления первых живых организмов свободного кислорода в атмосфере древней Земли еще не было, поэтому у них был анаэробный (безкислородный) тип дыхания. Таким образом, первыми живыми организмами на Земле были, по-видимому, гетеротрофные (питающиеся готовыми органическими веществами) бактерии- анаэробы (рис. 1).
Несмотря на то что бактерии-анаэробы возникли в глубокой древности, они широко распространены на Земле и в настоящее время. Их можно встретить и в банке с простоквашей, и в бочонке с солеными огурцами или капустой. Молочнокислые бактерии — факультативные анаэробы (они могут расти и развиваться в присутствии кислорода, но в процессе дыхания кислород не используют).

Рис. 1. Симбиотическая гипотеза происхождения эукариот
Анаэробами являются и многие почвенные бактерии, например возбудители столбняка, газовой гангрены, ботулизма. Все они облигатные анаэробы. В отличие от факультативных анаэробов, облигатные анаэробы не переносят присутствия кислорода в окружающей среде, кислород для них — яд. Именно поэтому опасность заражения столбняком гораздо выше, если рана колотая и инфекция в ней развивается без доступа кислорода. Значительно менее опасны открытые раны и ссадины. Газовая гангрена тоже, как правило, начинает развиваться после наложения на поврежденную конечность препятствующей доступу кислорода гипсовой повязки. Опасность тяжелого пищевого отравления — ботулизма — возникает при домашнем консервировании, когда воздух удален предварительным кипячением, а герметичная крышка препятствует поступлению кислорода извне. В случае засолки огурцов или грибов в открытой посуде возбудитель ботулизма не разовьется, поскольку он — облигатный анаэроб. При домашнем консервировании возбудитель ботулизма уничтожить чрезвычайно трудно, поскольку его споры выдерживают 5-6 часов непрерывного кипячения. Поэтому промышленное консервирование проводят перегретым паром под давлением при температуре не 100, а 130°С в течение 1-2 часов.
Бактерии-анаэробы древней Земли питались готовыми органическими веществами, образовавшимися в больших количествах на ранних этапах формирования Земли. Абиогенному синтезу органических веществ способствовали высокая температура атмосферы и бурная вулканическая деятельность. К моменту появления первых живых организмов Земля остыла и интенсивность абиогенного синтеза органических веществ значительно снизилась. Развитие анаэробов должно было неизбежно истощить запасы органических веществ, что, в свою очередь, привело бы к гибели всех живых организмов. Возможно, история развития жизни на Земле на этом и закончилась бы, если бы спустя всего 100 млн лет (3,4 млрд лет тому назад) под влиянием жесткой конкуренции за органические вещества на Земле не появилось новое поколение живых организмов - фото синтезирующие бактерии (см. рис. 1).
Уникальной особенностью этих живых существ была способность осуществлять фотосинтез, т.е. синтезировать органические вещества из неорганических с использованием энергии солнечного света. У первых фотосинтезирующих бактерий был необычный аноксигенный тип фотосинтеза (он протекает без выделения кислорода).
Как известно, строительными элементами, из которых фото- синтезирующие организмы создают органические вещества, являются углекислый газ и водород. Первые фотосинтезирующие бактерии отнимали водород не от воды, как это происходит у большинства современных фотосинтезирующих организмов, а от сероводорода (H2S), поскольку затраты энергии на отрыв атомов водорода от молекулы сероводорода в 7 раз меньше, чем на отрыв его от молекулы воды.
Фотосинтез с выделением кислорода появился позднее у цианобактерий (сине-зеленых водорослей). Именно цианобактерии впервые осуществили фотолиз воды, при котором с помощью энергии солнечного света водород, необходимый для биосинтеза органических веществ, отрывается от молекулы воды, а в качестве побочного продукта образуется свободный кислород.
Накопление в атмосфере свободного кислорода привело к коренному преобразованию условий жизни на Земле. К моменту появления первых живых организмов Земля сильно остывает, снижается количество грозовых разрядов в атмосфере, затухает вулканическая деятельность. Практически единственным источником энергии для абиогенного синтеза органических веществ является ультрафиолетовое излучение Солнца.
С появлением кислорода в верхних слоях атмосферы, на высоте 15-30 км, сформировался озоновый экран, защитивший живые организмы от губительного действия ультрафиолетового излучения, что послужило предпосылкой возникновения жизни не только в воде, но и на суше. Одновременно озоновый экран, снизив интенсивность падающего на Землю ультрафиолетового излучения, практически остановил абиогенный синтез органических веществ, вследствие чего дальнейшее существование жизни на Земле стало полностью зависеть от дея тельности фотосинтезирующих организмов.
Как отмечалось ранее, главный (целевой) продукт фотосинтеза — богатые энергией органические вещества, которые используются живыми организмами как для построения своего чела, так и для получения необходимой для их жизнедеятельности энергии, кислород же является побочным продуктом фотосинтеза. Поэтому для наиболее древних но происхождению живых организмов — бактерий-анаэробов и первых фотосинтезирующих бактерий кислород — это яд. Однако вслед за фотосинтезирующими бактериями на Земле появились живые организмы, которые научились не только защищаться от кислорода, но и использовать его — научились дышать кислородом. Это были бактерии-аэробы (или бактерии-окислители).
Биологические преимущества кислородного дыхания очевидны: при кислородном окислении органических веществ из единицы (например, из 1 г) органических веществ можно извлечь в 19 раз больше энергии, чем при бескислородном дыхании. Вследствие этою бактерии-аэробы оказались способными значительно экономнее расходовать органические вещества, чем анаэробы, что, в свою очередь, позволило им существовать в условиях относительно низких концентраций органических веществ.
Симбиотическая гипотеза происхождения эукариот
На ранних этапах биологической эволюции на Земле последовательно возникают, а затем сосуществуют 3 поколения прокариот: бактерии-анаэробы, фотосинтезирующие бактерии и бактерии- аэробы (см. рис. 1).
Фотосинтезирующие бактерии могли создавать органические вещества из неорганических, а бактерии-аэробы умели очень экономно их расходовать. Лишенные этих преимуществ бактерии-анаэробы вынуждены были эксплуатировать полезные свойства других живых организмов. Один из способов одностороннего использования одного организма другим — хищничество. На определенном этапе развития от бактерий-анаэробов произошли хищные амебовидные организмы, способные захватывать с помощью ложноножек и поглотать как фотосинтезирующих бактерий, так и бактерий-аэробов.
Однако не все амебовидные хищники переваривали захваченные бактерии, в некоторых случаях бактерии могли жить и размножаться внутри цитоплазмы хищника. Возникшее таким образом сообщество живых организмов обладало многими ценными свойствами: способностью к фотосинтезу, обусловленной деятельностью фотосинтезирующих бактерий, способностью к экономному и эффективному использованию органических веществ благодаря кислородному типу дыхания, характерному для бактерий-аэробов, и, наконец, способностью к активному передвижению и захвату добычи, свойственному хищной клетке-носите- лю. Со временем взаимовыгодные, симбиотические отношения этих трех групп организмов закрепились, стали устойчивыми: фотосинтезирующие бактерии превратились в хлоропласты, а аэробные бактерии-окислители - в энергетические станции клетки — митохондрии. Как митохондрии, так и хлоропласты и в настоящее время сохраняют собственный наследственный аппарат, размножаются независимо отделения клетки и наследуются через цитоплазму но материнской линии.
Для управления сложным сообществом живых организмов и защиты собственного генетического материала (ведь другие организмы, входящие в сообщество, имели свою генетическую программу) у клетки-носителя возникает специальная клеточная органелла - ядро.
Живые организмы, клетки которых имеют оформленное ядро, называются эукариотами (от греч. еu - хорошо, полностью и karyon — ядро). Все растения, животные и грибы — эукариоты. Наследственная информация в ядрах эукариотических клеток хранится в виде особых структур — хромосом, отчетливо видных под световым микроскопом в момент деления клетки. Первые эукариотические клетки появились на Земле около 2 млрд лет т.н.
Более древние по происхождению бактерии не имеют оформленного ядра.
Живые организмы, клетки которых не имеют оформленного ядра, называются прокариотами (от лат. pro — перед, раньше и греч. karyon — ядро). Все бактерии, в том числе и фотосинтезирующие, — прокариоты. Наследственная информация представлена в них одной-единственной кольцевой молекулой ДНК, лежащей непосредственно в цитоплазме и не различимой в обычный световой микроскоп.
Поскольку но современным научным представлениям вес эукариотические клетки представляют собой симбиотические сообщества двух или трех живых организмов, изложенную выше гипотезу происхождения эукариот называют симбиотической.
Первые эукариотические клетки, по-видимому, представляли собой амебовидные существа, многие из которых содержали как митохондрии, так и хлоропласты.
Около 1,5 млрд лет т.н. от них возникают более совершенные эукариотические организмы, способные к быстрому активному передвижению — древние жгутиковые (см. рис. 1). Принято считать, что жгутики, так же как в свое время митохондрии и хлоропласты, произошли от каких-то древних свободноживущих прокариот.
Древние жгутиковые, видимо, сочетали свойства растений и животных. Со временем те из них, которые оказались в среде с высоким содержанием органических веществ, утратили хлоропласты и превратились в одноклеточных животных — простейших, а сохранившие хлоропласты дали начало растениям. Естественно, наиболее древние по происхождению растения — одноклеточные, подвижные и имеют жгутики.
Дальнейший эволюционный прогресс животных связан с возрастанием роли активного передвижения, что вызвано необходимостью поиска пищи и захвата добычи. Совершенствуется и система управления движением, что, в конечном итоге, приводит к возникновению высокоорганизованной нервной системы и, наконец, интеллекта.
В то же время растения, обеспечивающие себя питанием за счет фотосинтеза, в процессе эволюции утрачивают способность к передвижению и приобретают множество приспособлений, повышающих эффективность фотосинтеза.
Таким образом, около 1,5 млрд лет т.н. от единого предка — древнего жгутикового возникают два важнейших царства живых организмов — царство растения и царство животные.

Креационизм: жизнь создана творцом — Богом.
Гипотеза биогенеза: согласно этой теории жизнь может зародиться только из живого.
Гипотеза панспермии (Г. Рихтер, Г. Гельмгольц, С. Аррениус, П. Лазарев): согласно этой гипотезе жизнь могла возникнуть один или несколько раз в космосе. На Земле жизнь появилась в результате занесения ее из космоса.
Гипотеза вечности жизни (В. Прейер, В.И. Вернадский): жизнь существовала всегда, проблемы происхождения жизни нет.
Теория абиогенеза: жизнь возникла из неживой материи путем самоорганизации простых органических соединений.
■ Для средних веков были характерны примитивные представления, допускавшие появление целых живых организмов из неживой материи (считалось, что лягушки и насекомые заводятся в сырой почве, мухи — из гнилого мяса, рыбы — из ила и т.д.).
■ Современной конкретизацией этой теории является коацерват-ная гипотеза Опарина — Холдейна.
Первыми носителями генетической информации стали молекулы РНК. Они образовывались с помощью протеиноидов, притягивающих определенные нуклеотиды, которые объединялись в цепочки РНК. Такая РНК несла информацию о структуре протеиноидов и притягивала к себе соответствующие аминокислоты, что приводило к воспроизводству точных копий протеиноидов. Позднее функции РНК перешли к ДНК (ДНК стабильнее РНК и может копироваться с большей точностью), а РНК стала выполнять роль посредника между ДНК и белком. В процессе эволюции преимуществом обладали те пробионты, у которых взаимодействие белков и нуклеиновых кислот было наиболее четким.
Эволюция пробионтов
Пробионты были анаэробными гетеротрофными прокариотами. Пищу и энергию для жизнедеятельности они получали из органических веществ абиогенного происхождения за счет анаэробного расщепления (брожения, или ферментации). Истощение запасов органических веществ усилило конкуренцию и ускорило эволюцию пробионтов.
В результате произошла дифференциация пробионтов. Одна их часть (примитивные предки современных бактерий), оставаясь анаэробными гетеротрофами, претерпела прогрессивное усложнение. Другие пробионты, содержащие определенные пигменты, приобрели возможность образовывать органические вещества путем фотосинтеза (сначала бескислородного, а затем — предки цианобактерий — с выделением кислорода). Т.е. возникли анаэробные автотрофные прокариоты, которые постепенно насыщали свободным кислородом атмосферу Земли.
С появлением кислорода возникли аэробные гетеротрофные прокариоты, существующие за счет более эффективного аэробного окисления органических веществ, образовавшихся в результате фотосинтеза.
Возникновение и эволюция эукариот и многоклеточных организмов
❖ Симбиотическая теория появления (около 1,5 млрд, лет назад) и эволюции эукариотических клеток (симбиогенез):
■ одна группа анаэробных гетеротрофных пробионтов вступила в симбиоз с аэробными гетеротрофными первичными бактериями, дав начало эукариотическим клеткам, имеющим в качестве энергетических органоидов митохондрии;
■ другая группа анаэробных гетеротрофных пробионтов объединилась не только с аэробными гетеротрофными бактериями, но и с первичными фотосинтезирующими цианобактериями, дав начало эукариотическим клеткам, имеющим в качестве энергетических органоидов хлоропласты и митохондрии. Клетки-симбионты с митохондриями в дальнейшем дали начало царствам животных и грибов; с хлоропластами — царству растений.
Усложнение эукариот привело к появлению клеток с полярными свойствами, способными к взаимному притяжению и слиянию, т.е. к половому процессу, диплоидности (следствие этого — мейоз), доминантности и рецессивности, комбинативной изменчивости и т.д.
❖ Гипотезы появления многоклеточных организмов (2,6 млрд, лет назад):
■ гипотеза гастреи (Э. Геккель, 1874 г.): предковыми формами многоклеточных были одноклеточные организмы, образовавшие однослойную сферическую колонию. Позднее за счет впя-чивания (инвагинации) части стенки колонии образовался гипотетический двуслойный организм — гастрея, подобный стадии гаструлы эмбрионального развития животных; при этом клетки наружного слоя выполняли покровную и двигательную функции, клетки внутреннего слоя — функции питания и размножения;
■ гипотеза фагоцителлы (И.И. Мечников, 1886 г.; эта гипотеза лежит в основе современных представлений о возникновении многоклеточное™): многоклеточные произошли от одноклеточных колониальных жгутиковых организмов. Способом питания таких колоний был фагоцитоз. Клетки, захватившие добычу, перемещались внутрь колонии, и из них образовывалась ткань — энтодерма, выполняющая пищеварительную функцию. Клетки, оставшиеся снаружи, выполняли функции восприятия внешних раздражений, защиты и движения; из них впоследствии развилась покровная ткань — эктодерма. Часть клеток специализировалась на выполнении функции размножения. Постепенно колония превратилась в примитивный, но целостный многоклеточный организм — фагоцителлу. Подтверждением этой гипотезы служит ныне существующий, промежуточный между одной и многоклеточными, организм трихоплакс, строение которого соответствует строению фагоцителлы.
Основные этапы эволюции растений
Исторические этапы
Разделение эукариот на несколько ветвей, от которых произошли растения, грибы и животные (около 1—1,5 млрд, лет назад). Первые растения были водорослями, большая часть которых свободно плавала в воде, остальные прикреплялись ко дну.
Появление первых наземных растений — риниофитов (около 500 млн. лет назад в результате процесса горообразования и сокращения площади морей часть водорослей оказалась в мелких водоемах и на суше; некоторые из них погибли, другие адаптировались, приобретя новые признаки: у них образовались ткани, которые затем дифференцировались на покровные, механические и проводящие; бактерии, взаимодействуя с минералами земной поверхности, образовали на суше почвенный субстрат). Споровое размножение риниофитов.
Вымирание риниофитов и появление плаунов, хвощей и папоротников (около 380-350 млн. лет назад); возникновение вегетативных органов (что повысило эффективность функционирования отдельных частей растений); появление семенных папоротниковидных и хвойных.
Появление голосеменных растений (около 275 млн. лет назад), которые могли обитать в более сухой среде; вымирание семенных папоротников и древовидных споровых растений; у высших наземных растений постепенная редукция гаплоидного поколения (гаметофита) и преобладание диплоидного поколения (спорофита).
Появление диатомовых водорослей (около 195 млн. лет назад).
Появление покрытосеменных растений (около 135 млн. лет назад); расцвет диатомовых водорослей.
Вымирание многих видов растений (около 2,5 млн. лет назад), упадок древесных форм, расцвет травянистых; приобретение растительным миром современных форм.
Биологические этапы

1. Переход от гаплоидности к диплоидности. Диплоидность смягчает влияние неблагоприятных рецессивных мутаций на жизнеспособность и дает возможность накопить резерв наследственной изменчивости. Этот переход прослеживается и при сопоставлении современных групп растений. Так, у многих водорослей все клетки, кроме зигот, гаплоидны. У мхов преобладает гаплоидное поколение (взрослое растение) при сравнительно слабом развитии диплоидного (органы спороношения). У более высокоорганизованных бурых водорослей наряду с гаплоидными существуют и диплоидные особи. Но уже у папоротников преобладает диплоидное поколение, а у голосеменных (сосны, ели и др.) и покрытосеменных растений (многие деревья, кустарники, травы) самостоятельно существуют только диплоидные особи (см. рис.).
2. Утрата связи процесса полового размножения с водой, переход от наружного оплодотворения к внутреннему.
3. Разделение тела на органы (корень, стебель, лист), развитие проводящей системы, усложнение строения тканей.
4. Специализация опыления с помощью насекомых и распространение семян и плодов животными.
Основные этапы эволюции животных
❖ Важнейшие биологические этапы эволюции:
■ возникновение многоклеточное и все большее расчленение и дифференциация всех систем органов;
■ возникновение твердого скелета (наружного у членистоногих, внутреннего у позвоночных);
■ развитие центральной нервной системы;
■ развитие общественного поведения в разных группах высокоорганизованных животных, которое, вместе с накоплением ряда крупных ароморфозов, привело к возникновению человека и человеческого общества.

Важнейшие ароморфозы и их результаты

Геохронологическая шкала Земли
❖ Катархейская эра (4,7-3,5 млрд, лет назад): климат очень жаркий, сильная вулканическая деятельность; происходит химическая эволюция, возникают биополимеры.
❖ Архейская эра (3,5-2,6 млрд, лет назад) — эра зарождения жизни. Климат жаркий, активная вулканическая деятельность; возникновение жизни на Земле, появление на границе водной и наземно-воздушной сред первых организмов (анаэробных ге-теротрофов) — пробионтов. Появление анаэробных автотрофных организмов, архебактерий, цианобактерий; образование отложений графита, серы, марганца, слоистых известняков как результат жизнедеятельности архебактерий и цианобактерий. В конце архея — возникновение колониальных водорослей. Появление кислорода в атмосфере.
❖ Протерозойская эра (2,6-0,6 млрд, лет назад) — эра ранней жизни; делится на ранний протерозой (2,6-1,65 млрд, лет назад) и поздний протерозой (1,65-0,6 млрд, лет назад). Характеризуется интенсивным горообразованием, многократными похолоданиями и оледенениями, активным формированием осадочных пород, образованием в атмосфере кислорода (в конце эры — до 1%), началом формирования защитного озонового слоя в атмосфере Земли. В органическом мире: развитие одноклеточных прокариотических и эукариотических фотосинтезирующих организмов, возникновение полового процесса, переход от ферментации к дыханию (ранний протерозой); появление низших водных растений — строматолитов, зеленых водорослей и др. (поздний протерозой), а к концу эры — всех типов беспозвоночных многоклеточных (кроме хордовых): губок, кишечнополостных, червей, моллюсков, иглокожих и др.
❖ Палеозойская эра (570-230 млн. лет назад) — эра древней жизни; делится на 6 периодов: кембрий, ордовик, силур, девон, карбон и пермь.
■ Кембрий (570-490 млн. лет назад): климат умеренный, материк Пангея начал погружаться в воды океана Тетис. В органическом мире: жизнь сосредоточена в морях; эволюция многоклеточных форм; расцвет основных групп водорослей (зеленых, красных, бурых и др.) и морских беспозвоночных животных с хитиновофосфатной раковиной (особенно трилобитов и археоцеатов).
■ Ордовик (490-435 млн. лет назад): климат теплый, погружение Пангеи достигает максимума. В конце периода — освобождение от воды значительных территорий. В органическом мире: обилие и разнообразие водорослей; появление кораллов, морских иглокожих, полухордовых (граптолитов), первых хордовых (бесчелюстных рыб) и первых наземных растений — риниофи-тов. Господство трилобитов.
■ Силур (435-100 млн. лет назад): климат засушливый и прохладный; происходит подъем суши и интенсивное горообразование; концентрация О2 в атмосфере достигает 2%; завершается формирование защитного озонового слоя. В органическом мире: заселение суши сосудистыми растениями (риниофитами) и формирование на ней почвы; возникновение современных групп водорослей и грибов; расцвет в морях трилобитов, граптолитов, кораллов, ракоскорпионов; появление челюстных хордовых (панцирных и хрящевых рыб) и первых наземных членистоногих (скорпионов).
■ Девон (400-345 млн. лет назад): климат резко континентальный; оледенение, дальнейший подъем суши, полное освобождение от моря Сибири и Восточной Европы; концентрация О2 в атмосфере достигает современной (21%). В органическом мире: расцвет риниофитов, а затем (к концу периода) их вымирание; появление основных групп споровых растений (мохообразных, папоротниковидных, плауновидных, хвощевидных), а также примитивных голосеменных (семенных папоротников); расцвет древних беспозвоночных, а затем вымирание многих их видов, как и большинства бесчелюстных; появление бескрылых насекомых и паукообразных; расцвет в морях панцирных, кистеперых и двоякодышащих рыб; выход на сушу первых четвероногих позвоночных (стегоцефалов) — предков земноводных.
■ Карбон (каменноугольный период) (345-280 млн. лет назад): климат жаркий и влажный (в Северном полушарии), холодный и сухой (в Южном полушарии); материки низменные с обширными болотами, в которых шло образование каменного угля из стволов папоротниковидных. В органическом мире: расцвет древовидных споровых хвощевидных (каламитов), плауновидных (лепидодендронов и сигиллярий) растений и семенных папоротниковидных; появление первых голосеменных (хвойных); расцвет раковинных амеб (фораминифер), морских беспозвоночных, хрящевых рыб (акул); появление на суше первых амфибий, древних пресмыкающихся (котилозавров) и крылатых насекомых; вымирание граптолитов и панцирных рыб.
■ Пермь (280-240 млн. лет назад): усиливается засушливость, наступает похолодание, происходит интенсивное горообразование. В органическом мире: исчезновение лесов из древовидных папоротников; распространение голосеменных (гинкговых, хвойных); начало расцвета стегоцефалов и пресмыкающихся; распространение головоногих моллюсков (аммонитов) и костистых рыб; уменьшение количества видов хрящевых, кистеперых и двоякодышащих рыб; вымирание трилобитов.
❖ Мезозойская эра (240-67 млн. лет назад) — средняя эра в развитии жизни на Земле; делится на 3 периода: триас, юра, мел.
■ Триас (240-195 млн. лет назад): климат засушливый (появляются пустыни); начинается дрейф и разделение континентов (материк Пангея разделяется на Лавразию и Гондвану). В органическом мире: вымирание семенных папоротников; господство голосеменных (саговниковых, гинкговых, хвойных); развитие пресмыкающихся; появление головоногих моллюсков (белемнитов), первых яйцекладущих млекопитающих (триконодонтов) и первых динозавров; вымирание стегоцефалов и многих видов животных, процветавших в палеозойскую эру.
■ Юра (195-135 млн. лет назад): климат засушливый, материки подняты над уровнем моря; на суше большое разнообразие ландшафтов. В органическом мире: появление диатомовых водорослей; господство папоротников и голосеменных растений; расцвет головоногих и двустворчатых моллюсков, пресмыкающихся и гигантских ящеров (ихтиозавров, бронтозавров, диплодоков и др.); появление первых зубастых птиц (археоптериксов); развитие древних млекопитающих.
■ Мел (135—67 млн. лет назад): климат влажный (много болот); во многих районах похолодание; продолжается дрейф континентов; происходит интенсивное отложение мела (из раковин форам инифер). В органическом мире: господство голосеменных растений, сменяющееся их резким сокращением; появление первых покрытосеменных растений, их преобладание во второй половине периода; формирование кленовых, дубовых, эвкалиптовых и пальмовых лесов; расцвет летающих ящеров (птеродактилей и др.); начало расцвета млекопитающих (сумчатых и плацентарных); к концу периода вымирание гигантских ящеров; развитие птиц; появление высших млекопитающих.
❖ Кайнозойская эра (началась 67 млн. лет назад и продолжается по настоящее время) делится на 2 периода: третичный (палеоген и неоген) и четвертичный (антропоген).
■ Третичный период (от 67 до 2,5 млн. лет назад): климат теплый, к концу прохладный; завершение дрейфа континентов; материки приобретают современные очертания; характерно интенсивное горообразование (Гималаи, Альпы, Анды, Скалистые горы). В органическом мире: господство однодольных покрытосеменных и хвойных растений; развитие степей; расцвет насекомых, двустворчатых и брюхоногих моллюсков; вымирание многих форм головоногих моллюсков; приближение видового состава беспозвоночных к современному; широкое распространение костистых рыб, занимающих пресноводные водоемы и моря; дивергенция и расцвет птиц; развитие и расцвет сумчатых и плацентарных млекопитающих, сходных с современными (китообразных, копытных, хоботных, хищных, приматов и др.), в палеогене — начало развития антропоидов, в неогене — появление предков человека (дриопитеков).
■ Четвертичный период (антропоген; начался 2,5 млн. лет назад): резкое похолодание климата, гигантские материковые оледенения (четыре ледниковых периода); формирование ландшафтов современного типа. В органическом мире: исчезновение в результате оледенений многих древних видов растений, господство двудольных покрытосеменных; упадок древесных и расцвет травянистых форм растений; развитие многих групп морских и пресноводных моллюсков, кораллов, иглокожих и др.; вымирание крупных млекопитающих (мастодонт, мамонт и др.); появление, доисторическое и историческое развитие человека: интенсивное развитие коры головного мозга, прямохождение.
Биологическая эволюция - это длительный процесс изменения живых существ под влиянием изменяющихся внешних условий.
• менялась морфология - появлялись новые ткани (зародышевые листки), органы и системы органов;
• менялась физиология - простейшие процессы добавлялись, усложнялись и совершенствовались;
• изменялся генетический состав живых организмов;
• на макроуровне (макроэволюция): появление новых типов, отделов, классов живых организмов и т.д.
• микроуровень - образование новых видов;
• часто это не образование чего-то нового, а упрощение в строении или вообще исчезновение старого - многие виды вымерли (даже сейчас, на момент нашего с вами существования) - многие виды живых организмов вымирают
Получается, что эволюция идет (это очень важно: эволюция идет и сейчас!) в двух направлениях:

Результаты эволюции.

Эволюционный процесс приводит к сохранению наиболее приспособленных особей в популяциях и вымиранию наименее приспособленных.
Что такое приспособленность? По-научному, это адаптация Так вот, эволюция привела к двум видам адаптации:
• на макроуровне - появление ароморфозов - прогрессивных изменений в строении (это могут быть целые системы органов);
• на микроуровне - появление идиоадаптаций - тоже изменения, но исключительно для приспособления к данной среде обитания, не прогрессивные.
Появляются новые ткани, развиваются и совершенствуются органы и системы органов.
Например, у наземных растений, в отличие от водорослей, появились ткани и органы.
Ткани и органы - те же, но немного видоизменяются, например: листья растений
разных регионов - в засушливых местах - колючки кактусов, широкие и большие листья- в тропических лесах (но все это - листья покрытосеменных растений)
Организм усложняется, преобразуется в другую таксономическую группу например:
рыбы: дыхание - жаберное, 1 круг кровообращения, сердце - 2 камеры амфибии: дыхание - легкими, 2 круга кровообращения, сердце - 3 камеры.
Организм принадлежит той же самой таксономической группе например:
ласты кита, лапки крота, ноги лошади - это все конечности млекопитающих.
Примеры заданий.

Конечно, это - ароморфоз!

1. Покровительственная окраска - идиоадаптация (мимикрия);
2. Редукция (исчезновение) пальцев у копытных - так они приспособились к жизни в степях, но при этом как были млекопитающими, так и остались - идиоадаптация;
3. Половое размножение - серьезное изменение, т.к. не было раньше, ароморфоз;
4. Шерсть у млекопитающих - один из основных признаков класса, значит, ароморфоз;
5. Кутикула - плотная кожица у растений - приспособление к среде, к защите от внешних условий, значит, идиоадаптация;
6. Мимикрия - как и пункт 1 - идиоадаптация
4) появление организмов, способных усваивать энергию света, разлагать воду и использовать продукты разложения (кислород и восстановленные органич. вещества) для биосинтеза; образование структуры трофич. уровней, биоценозов и формирование совр. биосферы;
5) появление разумных живых существ.
Первые два этапа относят к периоду хим. (предбиол.) эволюции; собственно Э. б. начинается с 3-го этапа. 30 , т. 10 лет) невероятна. В этом первая проблема Э. б.
Предлагаемое решение проблемы сводится к следующему. Первичный гиперцикл содержит всего две компоненты: двойную спираль (биспираль) ДНК произвольной последовательности и белок, имеющий форму чехла, окружающего биспираль, в периодически изменяющихся условиях (темп-ры, растворимости и т. д.) функционирующий как репликаза. Синтез такого белка может происходить путём полимеризации аминокислот, предварительно адсорбированных на биспирали ДНК (полимеризация на гетерогенном катализаторе), образующийся полимер - слепок с поверхности катализатора.
При адсорбции аминокислот на биспирали ДНК возможно избират. связывание аминокислот с определ. кодо-нами, имеющими к ним хим. сродство. Благодаря этому при последующей полимеризации образуется белок-чехол, аминокислотная последовательность к-рого коррелирует с последовательностью кодонов в матричной ДНК. Вероятность такого процесса не мала, поскольку он протекает без участия кода, трансляции, аппарата биосинтеза и не нуждается в содержащейся в них информации.
След. проблема - образование адаптера, т. е. молекул, комплементарных, с одной стороны, к кодону, а с другой- к аминокислотному остатку. Синтез их также может происходить на основе гетерогенного катализа. Набор адаптеров в случае отсутствия обратного вырождения (т. е. адаптеров, комплементарных к одинаковым кодо-нам и разным аминокислотам) существенно ускоряет репродукцию гиперцикла; наличие же обратного вырождения ведёт к прекращению репродукции и гибели гиперциклов.
Совокупность одинаковых гиперциклов, образовавшихся в результате комплементарной авторепродукции одной биспирали ДНК с одинаковым набором адаптеров, представляет собой первичный живой объект, обладающий информацией о коде. Популяция с др. предшественником (биспирали ДНК с др. последовательностью) обладает, вообще говоря, др. набором адаптеров, поскольку избирательность взаимодействия аминокислот и кодонов не столь велика, чтобы обеспечить однозначность алгоритма соответствия. Поэтому множество объектов с разными предшественниками ещё не представляет собой первичную биосферу, поскольку каждый из объектов обладает своим вариантом кода.
Проблема образования единого кода сводится к проблеме выбора в условиях сильного антагонистич. взаимодействия разных объектов, когда их адаптеры смешиваются и возникает обратное вырождение. Выбор единого кода описывается матем. моделью типа Вольтера - Лотки:
где -концентрация объектов i -го ( j- гo )типа, t - время; первый член описывает репродукцию, второй- антагонистич. взаимодействие разных объектов, третий- эффект "тесноты", т. е. конкуренцию одинаковых объектов. В случае, когда внутривидовой антагонизм меньше межвидового ( а 10 лет не мала, и процесс образования гена, кодирующего один столб, вполне вероятен. Накопление семи копий этого гена и соединение их в цепь (за счёт блочных мутаций), кодирующую весь мембранный белок, тоже достаточно вероятно.
3. Согласно принципу преемственности, возникновение новых ф-ций не должно мешать выполнению и сохранению прежних. В сложных организмах эти условия дополнительны (дуальны). Так, воздействие мутагенных факторов полезно для новых ф-ций и вредно для прежних. Проблема дуальности была решена путём удвоения всего генетич. материала (за счёт блочной мутации) и появления перекрёстного полового размножения. При этом отбор особей женского пола происходил по фенотипич. признакам (анатомическим и физиологическим), способствующим защите репродуктивных органов и сохранению генома. Напротив, отбор мужских особей происходил по признакам, способствующим накоплению новой информации.
4. Обмен генетич. информацией между клетками увеличивает фонд запасённых генов, что полезно в дивергентных стадиях. Он может происходить: а) в результате внедрения участков генома (плазмид) одной клетки в другую и встраивания его в ДНК; такой процесс происходит в колониях одноклеточных организмов, при внедрении чужеродной клетки в организм и между разными клетками одного организма; б) в результате внедрения одной клетки в другую и образования симбиотич. организма (напр., хлоропластов растит. клеток и митохондрий); в) при скрещивании; в этом случае увеличение информации происходит не путём увеличения ДНК, а благодаря удвоению числа хромосом.
3. Дискуссионные вопросы теории биологической эволюции
1) Согласно теории естеств. отбора, любая нейтральная генетич. информация должна элиминироваться как лишняя нагрузка, не дающая эволюц. преимуществ. Согласно нейтралистской теории эволюции, нейтральная информация должна сохраняться как материал, необходимый для выбора подходящего варианта при дивергенции. Разрешение этого противоречия состоит в том, что Э. б. представляет собой чередование конвергентных и дивергентных стадий. При конвергенции часть информации действительно элиминируется как избыточная, но значит. часть её остаётся на случай перехода в дивергентную фазу.
2) Согласно ортодоксальному дарвинизму, благоприятные фенотипич. признаки, приобретённые в результате упражнений, но не в результате мутаций, не могут передаваться потомству. Однако противоположное утверждение Ж. Б. Ламарка (J. В. Lamarck) не исключал и Ч. Дарвин (С. Darvin). Молекулярной основой первого утверждения является догма Крика, согласно к-рой белки (и др. метаболиты) не могут влиять на геном. Однако известные ныне и перечисленные выше механизмы генной регуляции допускают нарушение догмы Крика из-за влияния белков на эпигенетич. процессы, образование плазмид и внедрение их в геном репродуктивных органов. Важность разрешения этой проблемы в том, что даже редкие случаи наследования благоприобретённых признаков существенно ускоряют эволюц. процесс.
3) Ряд заболеваний связан с тем, что в клетку внедряется патогенный вирус, размножается в ней (активный вирус) и (или) встраивается в геном и размножается при его редупликации (провирус). В клетках организма обнаружен аппарат, способствующий этим процессам. Возникает вопрос: каким образом в Э. б. возник и сохранился аппарат, явно не способствующий выживанию данного организма?
Возможное решение вопроса состоит в том, что геном вируса аналогичен плазмиде и аппарат, приспособленный для обмена генетич. информацией, способствует развитию вирусных заболеваний. Последнее - плата организма за ускорение Э. б., в результате к-рой этот организм образовался.
Лит.: Оно С., Генетические механизмы прогрессивной эволюции, пер. с англ., M., 1973; Шноль С. Э., Физико-химические факторы биологической эволюции, M., 1979; Волькенштейн M. В., Молекулярная биофизика, M., 1975; его же, Общая биофизика, M., 1978; Романовский Ю. M., Степанова H. В., Чернавский Д. С., Математическое моделирование в биофизике, M., 1975; их же, Математическая биофизика, M., 1984; Геодакян В. А., Эволюционная теория пола, "Природа", 1991, № 8, с. 60. Д. С. Чернявский.
Физическая энциклопедия. В 5-ти томах. — М.: Советская энциклопедия . Главный редактор А. М. Прохоров . 1988 .
Читайте также: