
Аэробные окислительные процессы кратко
Обновлено: 02.07.2024
Клеточное дыхание — это окисление органических веществ в клетке, в результате которого синтезируются молекулы АТФ. Исходным сырьем (субстратом) обычно служат углеводы, реже жиры и еще реже белки. Наибольшее количество молекул АТФ дает окисление кислородом, меньшее – окисление другими веществами и переносом электронов.
Углеводы, или полисахариды, перед использованием в качестве субстрата клеточного дыхания распадаются до моносахаридов. Так у растений крахмал, а у животных гликоген гидролизуются до глюкозы.
Глюкоза является основным источником энергии почти для всех клеток живых организмов.
Первый этап окисления глюкозы — гликолиз. Он не требует кислорода и характерен как при анаэробном, так и аэробном дыхании.
Биологическое окисление
Клеточное дыхание включает в себя множество окислительно-восстановительных реакций, в которых происходит перемещение водорода и электронов от одних соединений (или атомов) к другим. При потери электрона каким-либо атомом происходит его окисление; при присоединении электрона — восстановление. Окисляемое вещество — это донор, а восстанавливаемое — акцептор водорода и электронов. Окислительно-восстановительные реакции, протекающие в живых организмах носят название биологического окисления, или клеточного дыхания.
Наиболее распространенным в природе конечным акцептором электронов является кислород, который восстанавливается. При аэробном дыхании в результате полного окисления органических веществ образуются углекислый газ и вода.
Биологическое окисление протекает по-этапно, в нем участвуют множество ферментов и соединения, переносящие электроны. При ступенчатом окислении электроны перемещаются по цепи переносчиков. На определенных этапах цепи происходит выделение порции энергии, достаточной для синтеза АТФ из АДФ и фосфорной кислоты.
Биологическое окисление весьма эффективно по-сравнению с различными двигателями. Около половины выделяющейся энергии в конечном итоге фиксируется в макроэргических связях АТФ. Другая часть энергии рассеивается в виде тепла. Поскольку процесс окисления ступенчатый, то тепловая энергия выделяется понемногу и не повреждает клетки. В то же время она служит для поддержания постоянной температуры тела.
Аэробное дыхание
Различные этапы клеточного дыхания у аэробных эукариот происходят
в матриксе митохондрий – цикл Кребса, или цикл трикарбоновых кислот,
на внутренней мембране митохондрий – окислительное фосфорилирование, или дыхательная цепь.
На каждом из этих этапов из АДФ синтезируется АТФ, больше всего на последнем. Кислород в качестве окислителя используется только на этапе окислительного фосфорилирования.
Суммарные реакции аэробного дыхания выглядит следующим образом.
Дыхательная цепь: 12H2 + 6O2 → 12H2O + 34АТФ
Таким образом биологическое окисление одной молекулы глюкозы дает 38 молекул АТФ. На самом деле нередко бывает меньше.
Анаэробное дыхание
Большинство анаэробов — это микроорганизмы. Однако к организмам, использующим анаэробное дыхание, относятся также дрожжи, ряд червей-паразитов. Способностью к анаэробному дыханию также обладают определенные ткани. Например, мышечные клетки, которые периодически могут испытывать недостаток кислорода.
При анаэробном дыхании в окислительных реакциях акцептор водорода НАД не передает водород в конечном итоге на кислород, которого в данном случае нет.
В качестве акцептора водорода может быть использована пировиноградная кислота, образующаяся при гликолизе.
У дрожжей пируват сбраживается до этанола (спиртовое брожение). При этом в процессе реакций образуется также углекислый газ и используется НАД:
CH3COCOOH (пируват) → CH3CHO (ацетальдегид) + CO2
Молочнокислое брожение происходит в животных клетках, испытывающих временный недостаток кислорода, и у ряда бактерий:
CH3COCOOH + НАД · H2 → CH3CHOHCOOH (молочная кислота) + НАД
Оба брожения не дают выхода АТФ. Энергию в данном случае дает только гликолиз, и составляет она всего две молекулы АТФ. Значительная часть энергии глюкозы так и не извлекается. Поэтому анаэробное дыхание считается малоэффективным.
Аэробное дыхание – это процесс, при котором организмы используют кислород для превращения топлива, такого как жиры и сахара, в химическую энергию. По сравнению, анаэробное дыхание не использует кислород.
Аэробное дыхание намного эффективнее, и вырабатывает АТФ намного быстрее, чем анаэробное дыхание, Это потому, что кислород является отличным акцептором электронов для химических реакций, участвующих в производстве АТФ.

В процессе гликолиза две молекулы АТФ потребляются и четыре производятся. Это приводит к чистому усилению двух молекул АТФ, продуцируемых для каждой молекулы сахара, расщепленной в результате гликолиза. На этом сходство аэробного и анаэробного дыхания заканчивается.
В клетках с кислородом и аэробным дыханием молекула сахара распадается на две молекулы пируват, В клетках, которые не имеют кислорода, молекула сахара расщепляется на другие формы, такие как лактат.
Различия
После гликолиза различные химические процессы дыхания могут идти несколькими путями:
- Клетки, использующие аэробное дыхание, продолжают свою цепочку переноса электронов в высокоэффективном процессе, который в результате дает 38 молекул АТФ из каждой молекулы сахара.
- Клетки, которые лишены кислорода, но обычно не используют анаэробное дыхание, как наши мускул клетки могут оставлять конечные продукты гликолиза сидя, получая только две АТФ на молекулу сахара, которую они расщепляют. Это неэффективный метод получения энергии дыханием.
- Клетки, которые сделаны для анаэробного дыхания, такие как многие типы бактерии, может продолжить цепочку переноса электронов, чтобы извлечь больше энергии из конечных продуктов гликолиза.
После гликолиза клетки, которые не используют кислород для дыхания, но переходят в цепь переноса электронов, могут использовать другой акцептор электронов, такой как сульфат или нитрат, для продвижения своей реакции.
Эти процессы представляют собой тип анаэробного дыхания, называемого «ферментация «. Некоторые типы реакций брожения производят алкоголь и углекислый газ. Вот как алкогольные напитки и хлеб сделаны.
Аэробное дыхание, с другой стороны, отправляет остатки пирувата после гликолиза по совершенно другому химическому пути, этапы которого подробно обсуждаются ниже.
Шаги Аэробного Дыхания
Общее уравнение
Уравнение для аэробного дыхания описывает реагенты и продукты всех его стадий, включая гликолиз. Это уравнение:
1 глюкоза + 6 O2 → 6 CO2 + 6 H2O + 38 АТФ
Таким образом, 1 молекула шестиуглеродной глюкозы и 6 молекул кислорода превращаются в 6 молекул углекислого газа, 6 молекул воды и 38 молекул АТФ. Реакции аэробного дыхания можно разбить на четыре стадии, описанные ниже.
гликолиз
Гликолиз является первой стадией аэробного дыхания и происходит в цитоплазма клетки. Он включает в себя расщепление 1 молекулы сахара с шестью углеродами на две молекулы пирувата с тремя углеродами. Этот процесс создает две молекулы АТФ.
Общее уравнение выглядит следующим образом:
C6H12O6 + 2 ADP + 2 PI + 2 NAD + → 2 Пируват + 2 ATP + 2 NADH + 2 H + + 2 H2O
Этот процесс уменьшает кофактор NAD + до NADH. Это важно, так как позже в процессе клеточное дыхание, NADH будет способствовать формированию гораздо большего количества АТФ через митохондрии «s цепь переноса электронов.
На следующей стадии пируват перерабатывают для превращения его в топливо для цикла лимонной кислоты, используя процесс окислительного декарбоксилирования.
Окислительное декарбоксилирование пирувата
2 (Пируват– + Коэнзим A + NAD + → Ацетил-КоА + CO2 + NADH)
Окислительное декарбоксилирование, иногда называемое реакцией связи или реакцией перехода, является связующим звеном между гликолизом и циклом лимонной кислоты. Это, как и гликолиз, происходит в цитоплазме. В этом процессе пируват объединяется с коферментом А для получения ацетил-КоА.
Эта реакция перехода важна, потому что ацетил-КоА является идеальным топливом для цикла лимонной кислоты, который, в свою очередь, может привести в действие процесс окислительного фосфорилирования в митохондриях, которые производят огромное количество АТФ.
Больше NADH также создано в этой реакции. Это означает больше топлива для создания большего количества АТФ позже в процессе клеточного дыхания.
Цикл лимонной кислоты

2 (ACETYL COA + 3 NAD + + FAD + ADP + PI → CO2 + 3 NADH + FADH2 + ATP + H + + COENZYME A)
Реакция происходит дважды для каждой молекулы глюкозы, поскольку существует два пирувата и, следовательно, две молекулы ацетил-КоА, генерируемые для входа в цикл лимонной кислоты.
И NADH, и FADH2 – еще один переносчик электронов для цепи переноса электронов – созданы. Все NADH и FADH2, созданные на предыдущих этапах, теперь вступают в игру в процессе окислительного фосфорилирования.
Таким образом, в каждом цикле цикла два атома углерода вступают в реакцию в форме ацетил-КоА. Они производят две молекулы углекислого газа. Реакции генерируют три молекулы NADH и одну молекулу FADH. Одна молекула АТФ производится.
Окислительного фосфорилирования
Окислительное фосфорилирование является основной энергией, обеспечивающей стадию аэробного дыхания. Он использует свернутые мембраны в митохондриях клетки для производства огромного количества АТФ.
34 (ADP + PI + NADH + 1/2 O2 + 2H + → ATP + NAD + + 2 H2O)
В этом процессе NADH и FADH2 отдают электроны, полученные ими из глюкозы на предыдущих этапах клеточного дыхания, в цепь транспорта электронов в мембране митохондрий.
Цепочка переноса электронов состоит из ряда белковые комплексы которые встроены в митохондриальную мембрану, включая комплекс I, Q, комплекс III, цитохром С и комплекс IV.
Все это в конечном итоге служит для передачи электронов с более высоких на более низкие энергетические уровни, собирая энергию, выделяемую в процессе. Эта энергия используется для питания протонных насосов, которые приводят к образованию АТФ.
Так же, как натриево-калиевый насос клеточная мембрана протонные насосы митохондриальной мембраны используются для генерации градиент концентрации которые могут быть использованы для питания других процессов.
В частности, белки канала представляют собой синтез АТФ, которые являются ферментами, которые производят АТФ. Когда протоны проходят через АТФ-синтаза они управляют образованием АТФ.
Это также шаг, который требует кислорода. Без молекул кислорода, чтобы принять истощенные электроны в конце цепи переноса электронов, электроны отступили бы, и процесс создания АТФ не мог бы продолжаться.
![]()
Аэробное дыхание – вот почему нам нужны и пища, и кислород, поскольку оба они необходимы для выработки АТФ, который позволяет нашим клеткам функционировать. Мы вдыхаем O2 и выдыхаем одинаковое количество молекул CO2. Откуда появился атом углерода? Это происходит от пищи, такой как сахар и жир, которую вы съели.
Именно поэтому вы дышите тяжелее и быстрее, выполняя сжигание калорий. Ваше тело использует кислород и сахар с большей скоростью, чем обычно, и вырабатывает больше АТФ для питания ваших клеток, а также больше отходов CO2.
Хотя наши клетки обычно используют кислород для дыхания, когда мы используем АТФ быстрее, чем доставляем молекулы кислорода в наши клетки, наши клетки могут выполнять анаэробное дыхание, чтобы удовлетворить свои потребности в течение нескольких минут.
Интересный факт: накопление лактата в результате анаэробного дыхания является одной из причин, почему мышцы могут чувствовать боль после интенсивные упражнения!
Гликолиз - процесс окисления глюкозы, при котором из одной молекулы глюкозы образуются две молекулы пировиноградной кислоты, не является мембранозависимым процессом. Он происходит в цитоплазме. Однако ферменты гликолиза связаны со структурами цитоскелета. Суть гликолиза состоит в том, что молекула глюкозы (C6H12O6) без участия кислорода распадается на две молекулы пировиноградной кислоты (СН3СОСООН). При этом окисление идет за счет отщепления от молекулы глюкозы четырех атомов водорода, связывающихся со сложным органическим веществом НАД с получением двух молекул НАД•Н. Выделяющаяся при этом энергия запасается (40% от общего количества) в виде макроэргических связей двух молекул АТФ. 60% энергии выделяется в виде тепла. При последующем окислении НАД•Н получается еще 6 молекул АТФ. Таким образом, полный энергетический выход гликолиза в анаэробных условиях составляет 8 молекул АТФ.
Аэробный процесс гликолиза (10 реакций), уравнение (с образованием пирувата):
Анаэробный процесс гликолиза (11 реакций), уравнение (с образованием лактата):
Схема процесса гликолиза и его реакции

На схеме в рамках обозначены исходные субстраты и конечные продукты гликолиза, цифрами в скобках - число молекул.
ATP (АТФ) - это аденозинтрифосфорная кислота, универсальный источник энергии
ADP (АДФ) - это аденозиндифосфат, нуклеотид, участвует в энергетическом обмене
NAD (НАД) - никотинамидадениндинуклеотидфосфата
NADH (НАД•Н) - востановленная форма NAD
Таблица процесс гликолиза его реакции
Для распада и частичного окисления молекулы глюкозы требуется протекание 11 сложных последовательных реакций.
Гликолиз (от греч. glycys – сладкий и lysis – растворение, распад) – это последовательность ферментативных реакций, приводящих к превращению глюкозы в пируват с одновременным образованием АТФ.
При аэробных условиях пируват проникает в митохондрии, где полностью окисляется до СО2 и Н2О. Если содержание кислорода недостаточно, как это может иметь место в активно сокращающейся мышце, пируват превращается в лактат.
Итак, гликолиз – не только главный путь утилизации глюкозы в клетках, но и уникальный путь, поскольку он может использовать кислород, если
последний доступен (аэробные условия), но может протекать и в отсутствие кислорода (анаэробные условия).
Анаэробный гликолиз – сложный ферментативный процесс распада глюкозы, протекающий в тканях человека и животных без потребления кислорода. Конечным продуктом гликолиза является молочная кислота. В процессе гликолиза образуется АТФ. Суммарное уравнение гликолиза можно представить следующим образом:

В анаэробных условиях гликолиз – единственный процесс в животном организме, поставляющий энергию. Именно благодаря гликолизу организм человека и животных определенный период может осуществлять ряд физиологических функций в условиях недостаточности кислорода. В тех случаях, когда гликолиз протекает в присутствии кислорода, говорят об аэробном гликолизе .
Последовательность реакций анаэробного гликолиза, так же как и их промежуточные продукты, хорошо изучена. Процесс гликолиза катализируется одиннадцатью ферментами, большинство из которых выделено в гомогенном, клисталлическом или высокоочищенном виде и свойства которых достаточно известны. Заметим, что гликолиз протекает в гиало-плазме (цитозоле) клетки.
Первой ферментативной реакцией гликолиза является фосфорилирование, т.е. перенос остатка ортофосфата на глюкозу за счет АТФ. Реакция катализируется ферментом гексокиназой:
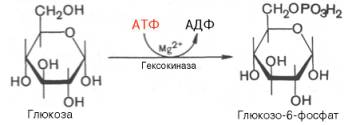
Образование глюкозо-6-фосфата в гексокиназной реакции сопровождается освобождением значительного количества свободной энергии системы и может считаться практически необратимым процессом.
Наиболее важным свойством гексокиназы является ее ингибирование глюкозо-6-фосфатом, т.е. последний служит одновременно и продуктом реакции, и аллостерическим ингибитором.
Фермент гексокиназа способен катализировать фосфорилирование не только D-глюкозы, но и других гексоз, в частности D-фруктозы, D-маннозы и т.д. В печени, кроме гексокиназы, существует фермент глюкокиназа, который катализирует фосфорилирование только D-глюкозы. В мышечной ткани этот фермент отсутствует (подробнее см. главу 16).
Второй реакцией гликолиза является превращение глюкозо-6-фос-фата под действием фермента глюкозо-6-фосфатизомеразы во фруктозо-6-фосфат:
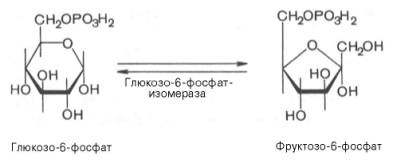
Эта реакция протекает легко в обоих направлениях, и для нее не требуется каких-либо кофакторов.
Третья реакция катализируется ферментом фосфофруктокиназой; образовавшийся фруктозо-6-фосфат вновь фосфорилируется за счет второй молекулы АТФ:
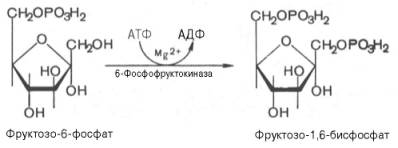
Данная реакция аналогично гексокиназной практически необратима, протекает в присутствии ионов магния и является наиболее медленно текущей реакцией гликолиза. Фактически эта реакция определяет скорость гликолиза в целом.
Фосфофруктокиназа относится к числу аллостерических ферментов. Она ингибируется АТФ и стимулируется АМФ . При значительных величинах отношения АТФ/АМФ активность фосфофруктокиназы угнетается и гликолиз замедляется. Напротив, при снижении этого коэффициента интенсивность гликолиза повышается. Так, в неработающей мышце активность фосфофруктокиназы низкая, а концентрация АТФ относительно высокая. Во время работы мышцы происходит интенсивное потребление АТФ и активность фосфофруктокиназы повышается, что приводит к усилению процесса гликолиза.
Четвертую реакцию гликолиза катализирует фермент альдолаза. Под влиянием этого фермента фруктозо-1,6-бисфосфат расщепляется на две фосфотриозы:
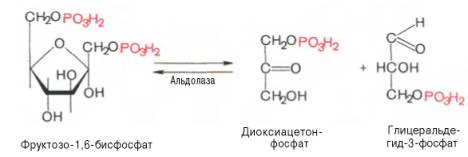
Эта реакция обратима. В зависимости от температуры равновесие устанавливается на различном уровне. При повышении температуры реакция сдвигается в сторону большего образования триозофосфатов (дигидро-ксиацетонфосфата и глицеральдегид-3-фосфата).
Пятая реакция – это реакция изомеризации триозофосфатов. Катализируется ферментом триозофосфатизомеразой:

Равновесие данной изомеразной реакции сдвинуто в сторону дигид-роксиацетонфосфата: 95% дигидроксиацетонфосфата и около 5% глице-ральдегид-3-фосфата. В последующие реакции гликолиза может непосредственно включаться только один из двух образующихся триозофосфатов, а именно глицеральдегид-3-фосфат. Вследствие этого по мере потребления в ходе дальнейших превращений альдегидной формы фосфотриозы ди-гидроксиацетонфосфат превращается в глицеральдегид-3-фосфат.
Образованием глицеральдегид-3-фосфата как бы завершается первая стадия гликолиза. Вторая стадия – наиболее сложная и важная. Она включает окислительно-восстановительную реакцию (реакция гликолитической оксидоредукции), сопряженную с субстратным фосфорилированием, в процессе которого образуется АТФ.
В результате шестой реакции глицеральдегид-3-фосфат в присутствии фермента глицеральдегидфосфатдегидрогеназы, кофермента НАД и неорганического фосфата подвергается своеобразному окислению с образованием 1,3-бисфосфоглицериновой кислоты и восстановленной формы НАД (НАДН). Эта реакция блокируется йод- или бромацетатом, протекает в несколько этапов:
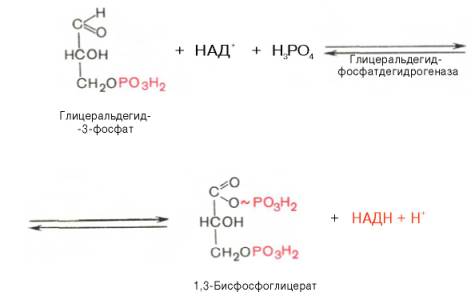
Седьмая реакция катализируется фосфоглицераткиназой, при этом происходит передача богатого энергией фосфатного остатка (фосфатной группы в положении 1) на АДФ с образованием АТФ и 3-фосфогли-цериновой кислоты (3-фосфоглицерат):
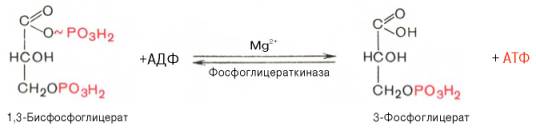
Таким образом, благодаря действию двух ферментов (глицеральде-гидфосфатдегидрогеназы и фосфоглицераткиназы) энергия, высвобождающаяся при окислении альдегидной группы глицеральдегид-3-фосфата до карбоксильной группы, запасается в форме энергии АТФ. В отличие от окислительного фосфорилирования образование АТФ из высокоэнергетических соединений называется субстратным фосфорилированием.
Восьмая реакция сопровождается внутримолекулярным переносом оставшейся фосфатной группы, и 3-фосфоглицериновая кислота превращается в 2-фосфоглицериновую кислоту (2-фосфоглицерат).
Реакция легкообратима, протекает в присутствии ионов Mg 2+ . Кофактором фермента является также 2,3-бисфосфоглицериновая кислота аналогично тому, как в фосфоглюкомутазной реакции роль кофактора выполняет глюкозо-1,6-бисфосфат:

Девятая реакция катализируется ферментом енолазой, при этом 2-фосфоглицериновая кислота в результате отщепления молекулы воды переходит в фосфоенолпировиноградную кислоту (фосфоенолпируват), а фосфатная связь в положении 2 становится высокоэргической:
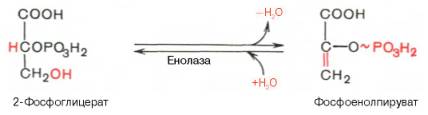
Енолаза активируется двухвалентными катионами Mg 2+ или Мn 2+ и ингибируется фторидом.
Десятая реакция характеризуется разрывом высокоэргической связи и переносом фосфатного остатка от фосфоенолпирувата на АДФ (субстратное фосфорилирование). Катализируется ферментом пируваткиназой:
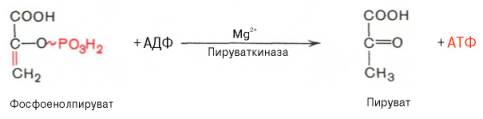
Для действия пируваткиназы необходимы ионы Mg 2+ , а также одновалентные катионы щелочных металлов (К + или др.). Внутри клетки реакция является практически необратимой.
В результате одиннадцатой реакции происходит восстановление пировиноградной кислоты и образуется молочная кислота. Реакция протекает при участии фермента лактатдегидрогеназы и кофермента НАДН, образовавшегося в шестой реакции:

Последовательность протекающих при гликолизе реакций представлена на рис. 10.3.
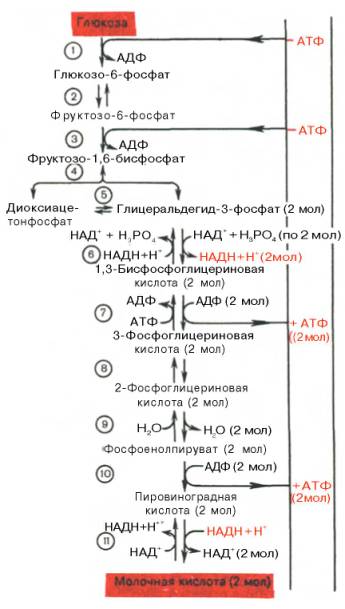
Рис. 10.3. Последовательность реакций гликолиза.
1 - гексокиназа; 2 - фосфоглюкоизоме-раза; 3 - фосфофруктокиназа; 4 - альдо-лаза; 5 - триозофосфатизомераза; 6 - гли-церальдегидфосфатдегидрогеназа; 7 -фосфоглицераткиназа; 8 - фосфоглицеромутаза; 9 - енолаза; 10 - пируватки-наза; 11 - лактатдегидрогеназа.
Реакция восстановления пирувата завершает внутренний окислительно-восстановительный цикл гликолиза. НАД + при этом играет роль промежуточного переносчика водорода от глицеральдегид-3-фосфата (6-я реакция) на пировиноградную кислоту (11-я реакция), при этом сам он регенерируется и вновь может участвовать в циклическом процессе, получившем название гликолитический оксидоредукции.
Биологическое значение процесса гликолиза заключается прежде всего в образовании богатых энергией фосфорных соединений. На первых стадиях гликолиза затрачиваются 2 молекулы АТФ (гексокиназная и фосфофрук-токиназная реакции). На последующих образуются 4 молекулы АТФ (фосфоглицераткиназная и пируваткиназная реакции). Таким образом, энергетическая эффективность гликолиза в анаэробных условиях составляет 2 молекулы АТФ на одну молекулу глюкозы.
Как отмечалось, основной реакцией, лимитирующей скорость гликолиза, является фосфофруктокиназная. Вторая реакция, лимитирующая скорость и регулирующая гликолиз – гексокиназная реакция. Кроме того, контроль гликолиза осуществляется также ЛДГ и ее изоферментами.
В тканях с аэробным метаболизмом (ткани сердца, почек и др.) преобладают изоферменты ЛДГ1 и ЛДГ2 (см. главу 4). Эти изоферменты инги-бируются даже небольшими концентрациями пирувата, что препятствует образованию молочной кислоты и способствует более полному окислению пирувата (точнее, ацетил-КоА) в цикле трикарбоновых кислот.
В тканях человека, в значительной степени использующих энергию гликолиза (например, скелетные мышцы), главными изоферментами являются ЛДГ5 и ЛДГ4. Активность ЛДГ5 максимальна при тех концентрациях пирувата, которые ингибируют ЛДГ1. Преобладание изоферментов ЛДГ4 и ЛДГ5 обусловливает интенсивный анаэробный гликолиз с быстрым превращением пирувата в молочную кислоту.
Как отмечалось, процесс анаэробного распада гликогена получил название гликогенолиза. Вовлечение D-глюкозных единиц гликогена в процесс гликолиза происходит при участии 2 ферментов – фосфорилазы а и фосфо-глюкомутазы. Образовавшийся в результате фосфоглюкомутазной реакции глюкозо-6-фосфат может включаться в процесс гликолиза. После образования глюкозо-6-фосфата дальнейшие пути гликолиза и гликогенолиза полностью совпадают:
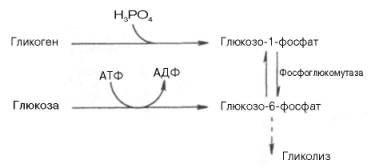
В процессе гликогенолиза в виде макроэргических соединений накапливаются не две, а три молекулы АТФ (АТФ не тратится на образование глюкозо-6-фосфата). Кажется, что энергетическая эффективность глико-генолиза выглядит несколько более высокой по сравнению с процессом гликолиза, но эта эффективность реализуется только при наличии активной фосфорилазы а. Следует иметь в виду, что в процессе активации фосфо-рилазы b расходуется АТФ (см. рис. 10.2).
Читайте также:
- Морфология и анатомия культуры их роль в исследовании конфигурации культуры кратко
- Правомерное поведение социально психологические и юридические аспекты тгп кратко
- Подходы к управлению организацией кратко
- Список для военкомата от школы образец
- Приказ о внесении изменений в тарификационный список доу