
Типы пищевых отношений в природе доклад
Обновлено: 02.07.2024
Природные экосистемы — устойчивые системы. Эта устойчивость обусловлена множеством взаимосвязей и взаимоотношений.
Различают горизонтальные и вертикальные отношения между особями в экосистемах. Горизонтальные — отношения между организмами одного трофического уровня, вертикальные — между организмами разных трофических уровней, т. е. с разным типом питания.
Вертикальные отношения более разнообразны и объединяются в два типа: мутуализм (взаимопомощь) и антагонизм — уничтожение одних особей другими в процессе питания. Все отношения в экосистемах — “полезные”; “вредных” организмов в природе нет. Главный тип горизонтальных отношений — это конкуренция за ресурсы.
Взаимосвязи организмов в экосистемах весьма многообразны. Наиболее важное значение имеют пищевые, или трофические, взаимосвязи: один организм поедается другим, тот — третьим и т. д. Ряд таких звеньев называется пищевой (трофической) цепью.
Пищевые цепи — последовательность организмов разных трофических уровней, в которых каждое предыдущее звено служит пищей для последующего; это основной канал переноса вещества и энергии в экосистемах.
В природе пищевые цепи редко изолированы друг от друга; в основном они взаимосвязаны и образуют сложную пищевую сеть (рис. 1.5).

Рис. 1.5. Пищевые отношения: а — пищевые цепи; б — пищевая сеть (Б. Небел)
Несмотря на многообразие возможных пищевых цепей и сложность пищевых сетей, все они соответствуют простой общей схеме: организмы последующего звена поедают организмы предыдущего, и таким образом осуществляется цепной перенос энергии и вещества, лежащий в основе круговорота веществ в природе. При каждом переходе от звена к звену теряется большая часть (до 90%) потенциальной энергии, рассеивающейся в виде тепла. По этой причине число звеньев в цепи ограничено и не превышает 4-5. Принципиальная схема пищевой цепи приведена на рис. 1.6.
Основу любой пищевой цепи составляют автотрофные организмы, или продуценты — преимущественно зеленые растения. Растения являются первичными поставщиками вещества и энергии для всех других организмов в цепях питания. Следующие звенья цепи питания занимают гетеротрофные организмы — консументы, потребляющие органические вещества. Начиная со звена продуцентов, имеются два основных пути использования энергии. Во-первых, она используется травоядными животными (фитофагами), во-вторых, ее потребляют сапрофаги в виде уже отмерших тканей (например, при разложении прошлогодней травы). Сапрофаги (преимущественно грибы, черви, микроорганизмы) получают необходимую энергию, разлагая мертвое органическое вещество. В соответствии с этим существует два вида пищевых цепей: цепи поедания и цепи разложения (рис. 1.7). Пищевые цепи разложения так же важны, как и цепи поедания.
Пищевые сети в микробных сообществах очень сложны. Главное их отличие от пищевых цепей, связывающих растения и животных, состоит в том, что бактерии, как правило, не поедают друг друга, а потребляют продукты жизнедеятельности своих соседей. Соседи при этом не только не страдают, но и выигрывают, поскольку накопление в окружающей среде продуктов жизнедеятельности может привести к их гибели.
Положение вида в цепи питания и его отношения с партнерами определяют его экологическую нишу (место, занимаемое в биогеоценозе).
Все виды, образующие пищевую цепь, существуют за счет продуцентов, т. е. за счет органического вещества, созданного зелеными растениями. При этом всего около 20% энергии, получаемой от Солнца, используется продуцентами, из которой 0,3% расходуется на фотосинтез. Более половины энергии, связанной при фотосинтезе, тут же расходуется в процессе дыха-

Рис. 1.6. Схема пищевой цепи и переноса вещества (сплошная линия) и энергии (пунктирная линия)

Рис. 1.7. Цепи поедания (выедания) и разложения
ния самих растений, что связано с окислением органического вещества (например, глюкозы) под действием кислорода воздуха:

Другая часть энергии переносится посредством ряда организмов по пищевым цепям.
Большая часть энергии, содержащейся в пище, расходуется животными на различные процессы жизнедеятельности, превращаясь при этом в тепло и рассеиваясь. Только 5-10% энергии пищи расходуется на построение тела животного, т. е. его биомассы.
Биомасса растений составляет первый трофический уровень. Биомасса 2-го трофического уровня (фитофаги) в 20~50 раз меньше, поскольку растительноядные съедают около 10% биомассы растений, из которых на дыхание и с экскрементами теряется 90% энергии. Биомасса плотоядных по тем же причинам в 10 и более раз меньше, чем растительноядных (правило 10%).
В процессе питания в экосистеме осуществляется передача вещества и энергии с одного трофического уровня на другой. Часть вещества и энергии при этом теряется. Соотношение между продуцентами, консументами и редуцентами в естественных экосистемах графически можно представить в виде пирамиды (рис. 1.8).

Рис. 1.8. Пирамида биомасс наземных экосистем (Б. Небел)
Это соотношение может быть выражено в единицах биомассы (сырой или сухой массы) — пирамида биомасс, числе особей — пирамида чисел или энергии, заключенной в особях — пирамида энергии.
Сопоставление биомасс суши и океана показано в табл. 1.4.
Сопоставление биомасс суши и океана
Наименование
Количество биомассы, млрд т
Живая масса
Сухая масса
Вес живое вещество
Продуктивность различных экосистем представлена в табл. 1.5.
Продуктивность различных экосистем (ЭЭС, К. С. Лосев)
Площадь 10 6 км 2
Биомасса растений, ГтС
Первичная продуктивность, ГтС/га год
Примечание. ГтС — гигатонна углерода, МтС — мегатонна углерода.
Звено пищевой цепи составляет уровень экологической пирамиды. Трофический уровень представляют в виде прямоугольника, длина или площадь которого пропорциональны биомассе, числу особей или энергии. Основанием экологической пирамиды служит первый трофический уровень — продуценты. Без непрерывного образования им органического вещества экосистема съела бы сама себя и прекратила бы свое существование. В наземных экосистемах с повышением трофического уровня запас биомассы уменьшается (см. рис. 1.8). В среднем каждое следующее звено в цепи питания содержит в 10 раз меньше биомассы, чем предыдущее звено.
В некоторых водных экосистемах запас биомассы увеличивается при переходе от продуцентов к консументам. Продуцентами в водных экосистемах являются водоросли — фитопланктон. Одноклеточные водоросли, преобладающие в нем, живут от нескольких дней до нескольких недель. Организмы — потребители планктона и последующих уровней живут дольше и накапливают большую биомассу. Как следствие, в водных экосистемах экологическая пирамида оказывается “перевернутой”, т. е. биомасса консументов выше, чем продуцентов (рис. 1.9).

Рис. 1.9. Вид экологической пирамиды в водной экосистеме
Такая “перевернутая” пирамида может наблюдаться не во все сезоны года. Летом, в период массового развития планктона (“цветения” воды), его биомасса может быть выше биомассы 2-го и 3-го трофических уровней.
В ряде глубоководных и подземных экосистем практически нет звена продуцентов — их продукция поступает от поверхностных экосистем.
Количество звеньев (трофических уровней) в наземных экосистемах может достигать четырех: растения — фитофаг — мелкий хищник — крупный хищник; в водных — шести: фитопланктон — растительноядный зоопланктон — плотоядный зоопланктон — мелкие рыбы — средние рыбы — крупные рыбы.
Структура живого в гидросфере представлена на рис. 1.10.

Рис. 1.10. Структура живого в гидросфере
Кроме того, существуют детритные пищевые цепи (их создают детритофаги, питающиеся мертвым органическим веществом): мертвый лист — дождевой червь — скворец — сокол. В пищевых сетях один и тот же вид становится жертвой разных хищников, и один хищник может питаться разными жертвами. На конце пищевых цепей до естественной смерти не доживает никто, рано или поздно каждый становится жертвой.
Из сказанного следует, что пищевые цепи — это путь однонаправленного потока солнечной энергии, поглощенной фо- тоавтотрофами (продуцентами), через живые организмы экосистемы, возвращаемой в окружающую среду в виде неэффективной тепловой энергии, а также движение питательных веществ от продуцентов к консументам, затем к продуцентам (биологический круговорот веществ) (см. рис. 1.6).
Производительность экосистемы измеряется массой органического вещества, созданного экосистемой за единицу времени на единицу площади, и называется биологической продуктивностью. Единицы измерения продуктивности: г/м 2 в день; кг/м 2 в год;
т/км 2 в год. Различают первичную биологическую продукцию, которую создают растения в процессе фотосинтеза, и вторичную биологическую продукцию, которую создают гетеротрофы в результате переработки растительной и животной массы.
Пищевые цепи реализуются через отношения “хищник — жертва” (хищник поедает жертву), “паразит— хозяин” (паразит питается хозяином). Хищник поедает жертву, паразит наносит вред, но не уничтожает хозяина.
В природе нет полезных и вредных организмов, все они вместе и по-разному вносят свой вклад в экологическое равновесие.

В природе ни один вид не существует в полной изоляции – все организмы взаимодействуют как с абиотической средой, так и с другими организмами.
Существуют следующие типы влияния одних организмов на другие:
- Положительные (+) – взаимодействие организмов в общей среде, при котором один организм получает выгоду от другого;
- Отрицательные (–) – взаимодействие организмов, при котором одному организму наносится вред из-за другого;
- Нейтральные (0) – когда организмы из общей среды обитания не оказывают никакого влияния друг на друга.
Основные типы отношений между организмами
Симбиотические отношения

Симбиоз (+ / +) описывает тесное и постоянное (долгосрочное) взаимодействие между двумя видами.
Симбиотические отношения подразделяются на облигатные (нужны для выживания) или факультативные (выгодные, но не строго необходимые).
Симбиотические отношения могут быть полезны как для одного, так и для обоих организмов в партнерстве:
- Мутуализм (+ / +) – оба вида извлекают выгоду от взаимодействия (анемон защищает рыбу-клоуна, рыба-клоун обеспечивает анемон питательными фекалиями);
- Комменсализм (0 / +) – один вид получает выгоду, а другой остается невредим (ракообразные прикрепляются к китам, чтобы добраться до мест богатых планктоном);
- Паразитизм (– / +) – один из видов получает выгоду в ущерб другому (например, клещи и блохи питаются кровью своего хозяина).
Хищник-Жертва

Хищничество (– / +) – это биологическое взаимодействие, при котором один организм (хищник) охотится и питается другим организмом (добычей).
Поскольку хищник полагается на добычу как на источник пищи, численность их популяций неразрывно связана между собой:
- Если численность популяции добычи сокращается (например, из-за чрезмерной охоты), численность хищников будет сокращаться по мере усиления внутривидовой конкуренции;
- Если численность популяция добычи растет, численность хищников будет увеличиваться в результате изобилия источника пищи.
Нейтрализм

Нейтрализм (0 / 0) описывает взаимосвязь между двумя видами , которые взаимодействуют, но никак не влияют друг на друга. Примеры истинного нейтрализму практически невозможно доказать. На практике термин используется для описания ситуаций, где взаимодействия пренебрежимо малы или незначительны.
Аменсализм

Аменсализм (– / 0) – межвидовые отношения, при которых одному организму наноситься вред, а другой остается невредимым. Хорошим примером аменсализма является крупный рогатый скот, который во время передвижения затаптывает копытами траву.
Конкуренция

Конкуренция (– / –) описывает взаимодействие между двумя организмами, при котором приспособленность одного снижается из-за присутствия другого.
Конкуренция может быть внутривидовой (между представителями одного вида) или межвидовой (между представителями разных видов).
Нехватка ресурсов (например, еды, воды или территории) обычно вызывают один из двух типов конкуренции:
Пищевые (трофические) связи - основные биотические связи. Они поддерживают жизнь организмов. Каждый вид служит источником энергии для других. Одни виды потребляют живую пишу, другие -мертвые остатки. Существование пищевых отношений между организмами противостоит геометрической прогрессии размножения, регулируя численность видов.[ . ]
В природе пищевые отношения могут осуществляться только в сосуществовании обеих форм, ибо трудно предположить, что животные, существуя и питаясь совместно, совершенно бы не оказывали друг на друга никакого влияния. Как мы видели, даже в экспериментальных условиях для получения величин, характеризующих только простые отношения, требовалось содержание животных в изолированном состоянии. Следовательно, простые отношения являются понятием, которое никогда в чистом виде не может иметь адекватного отображения в природе, и если мы вводим и анализируем его, то делаем это с единственной целью — количественно оценить различные элементы в общем механизме явления.[ . ]
Вопросам питания и пищевых отношений в экосистемах, как факторам, влияющим на поведение нектона, уделялось достаточно внимания. Трофологические исследования проводили по стандартной методике, принятой в ТИНРО-Центре.[ . ]
Подводя итоги главы о пищевых отношениях рыб, мы приходим к следующим выводам.[ . ]
Эволюционное значение пищевых отношений может стать объектом научного анализа лишь на основании четкого понимания следующего звена процесса — функциональной зависимости интенсивности размножения и, в равной степени, скорости вымирания животных от характера их питания. В настоящее время последний вопрос находится в состоянии почти полной неизученное™, особенно в отношении выяснения количественных зависимостей. Поэтому ведущим принципом наших исследований явилось изучение действия, оказываемого пищевыми отношениями на характер и интенсивность питания различных животных одного или нескольких видов.[ . ]
Новая глава в вопросе о пищевых отношениях у рыб начата Шорыгиным и его сотрудниками. Исследования Шорыгина представляют для нас особый интерес, так как, во-первых, объектом его работ, как указывалось, являются рыбы, и, во-вторых, этот автор впервые количественными методами изучения подошел непосредственно к вопросу питания, рассматривая его с точки зрения пищевых отношений.[ . ]
Следствием сложившихся пищевых отношений является расхождение доминант элективности, что в конечном итоге должно с неизбежностью привести к расхождению общих привычек питания. Эти же наблюдения позволяют говорить об относительности и условности отнесения той или иной формы к определенной экологической нише.[ . ]
Лишев М. Н. 1950 — Питание и пищевые отношения хищных рыб в бассейне Амура. Труды Амурской ихтиол, экспед., т. I.[ . ]
У яйца и эмбриона ведущие отношения возникают обычно на почве дыхания и защиты от хищников (Крыжановекий, 1949). В этих направлениях и наблюдается наибольшее разнообразие приспособлений. Пищевые взаимосвязи не имеют значения, так как питание идет за счет желтка, накопленного материнским организмом. В следующие периоды развития у рыбы в связи с переходом на внешнее питание ведущими становятся пищевые отношения. У взрослой рыбы, помимо пищевых, существенное значение приобретают взаимосвязи, возникающие на почве размножения.[ . ]
Следовательно, конкурентные пищевые отношения нарйду с непосредственным истреблением одними животными других являются стержневыми элементами борьбы за существование, й йх анализдолжен приблизить нас к познанию одного из важнейших явлений природы.[ . ]
Разобранные примеры косвенных пищевых отношений, протекающих в условиях полной монофагии, при всей своей кажущейся искусственности, имеют достаточно широкое распространение в природе. В -наиболее чистом виде монофагия присуща отдельным стадиям насекомых, главным образом растительноядных, реже хищных. Но и другие животные, даже высокопластичные в выборе пищи, часто сталкиваются с условиями, когда питание происходит в основном за счет одной, более или менее узкой категории пищевых объектов. Вместе с тем, анализ тенденции элективности, произведенный Шорыгиным для естественных условий и подтвержденный нами экспериментально, показал, что для воблы данный вид моллюсков не является предпочитаемым в комплексе других пищевых форм.[ . ]
Таким образом, формы косвенных пищевых отношений мы различаем по составу питающихся животных (гомоконкуренция и гетероконкуренция), по характеру пищи (монофагия и полифагия1), по характеру изменения рационов (качественные и количественные изменения), по механизму осуществления отношений (отношения простые и осложненные).[ . ]
В разрешении вопросов косвенных пищевых отношений больше, чем в других областях трофологии, нашел применение метод математической интерпретации получаемых результатов. В них не только используются для измерения- интенсивности процесса конкуренции различные количественные показатели, но и самый анализ его осуществляется с помощью более или менее сложного математического аппарата.[ . ]
Знание закономерностей„ межвидовых пищевых отношений имеет важное значение и при планировании акклиматизации новых видов рыб в те или иные водоемы.[ . ]
Отсюда, исходя из принципов проблемы пищевых отношений, экологическая ниша может быть определена как такое положение занимающего ее вида в экосистеме, когда показатели гетероконкуренции имеют наименьшее значение.[ . ]
Нам кажется, что перечисленные формы пищевых отношений достаточно полно охватывают различные случаи изучаемого .явления. С другой стороны, как будет показано ниже, они доступны для конкретного анализа, который в конечном счете и является основной задачей наших исследований.[ . ]
Поскольку интенсивность осложненных отношений является функцией совместного питания большей или меньшей группы животных, естественно было предположить, что чем выше численность группы, тем напряженнее сложатся косвенные пищевые отношения, давая более высокие значения показателя ц.[ . ]
При рассмотрении с точки зрения изучения пищевых отношений некоторого биологического комплекса, необходимо разграничивать группы организмов, слагающиеся в экологически однородные категории, т. е. занимающие определенное положение в системе трофических взаимоотношений. Характерной особенностью таких категорий или уровней, по терминологии Хатчинсона1, является использование для питания практически одинакового пищевого материала и отсутствие внутри уровня отношений типа хищник добыча.[ . ]
Следовательно, в простейшем случае, когда пищевые отношения определяются исключительно запасами пищи, приходящейся на каждого отдельного животного, их количественной мерой служит указанная функция.[ . ]
Методическое пособие по изучению питания и пищевых отношений рыб в естественных условиях.[ . ]
Интересным продолжением анализа косвенных пищевых отношений у рыб являются работы Никольского. На них мы остановимся подробнее при обсуждении полученных нами мате-< риалов.[ . ]
Благодаря определенной последовательности пищевых отношений различаются отдельные трофические уровни переноса веществ и энергии в экосистеме, связанные с питанием определенной группы организмов. Так, первый трофический уровень во всех экосистемах образуют продуценты — растения; второй — первичные консументы — фитофаги, третий — вторичные консументы — зоофаги и т.д. Как уже отмечалось, многие животные питаются не на одном, а на нескольких трофических уровнях (примером могут служить диеты серой крысы, бурого медведя и человека).[ . ]
В свете данных по анализу явления косвенных пищевых отношений по-новому приходится толковать причины, определяющие структуру некоторого совместно обитающего комплекса животных.[ . ]
В настоящем заключительном разделе о косвенных пищевых отношениях мы попытаемся рассмотреть некоторые общие вопросы, или непосредственно вытекающие из результатов произведенных опытов, или тесно с ними соприкасающиеся. Анализ вопросов, имеющих второстепенный характер, дан нами при изложении экспериментального материала.[ . ]
Забегая вперед, укажем еще на одно подразделение косвенных пищевых отношений: на простые и осложненные. Разграничение этих понятий относится непосредственно к механизму осуществления отношений и будет разобрано в следующей главе.[ . ]
При перенесении данного принципа на изучение проблемы косвенных пищевых отношений необходимо отметить следующее. Наблюдаемое в природе расхождение видов по различным нишам и ослабление этим путем гетероконкуренции по сравнению с внутривидовыми отношениями не только не является противоречием полученным экспериментальным данным, но, наоборот, целиком их подтверждает.[ . ]
Консервированные компоты, натуральные пюре из плодов и ягод — то ценные в пищевом отношении продукты.[ . ]
Принципиальная разница этих двух кардинальных условий осуществления косвенных пищевых отношений заключается, следовательно, в том, что в первом случае ухудшение условий питания, являющееся естественным результатом конкуренции, не может быть компенсировано хотя бы частичным переходом данного животного на качественно иной рацион. Во втором случае такая перемена неизбежно осуществляется. Переход же на иной рацион вызовет изменение показателей элективности, следовательно, и весь .анализ процесса потребует привлечения иного аппарата исследования.[ . ]
Отличительной особенностью нашего подхода к разбираемой проблеме является то, что пищевые отношения нами рассматриваются в первую очередь не как фактор эволюции вида, но, главным образом, как экологический процесс, выражаемый в определенных, количественно измеряемых категориях.[ . ]
Указывалось также, что основным фактором, обусловливающим наличие и интенсивность осложненных отношений, является причина рефлекторного порядка, мыслимая как взаимное отпугивание, взаимные помехи, взаимное беспокойство и пр. Заметим, что наличие нервных причин в осуществлении пищевых отношений отмечалось и ранее.[ . ]
Синтезом вопросов об интенсивности питания и об избираемости пищи является учение о конкурентных пищевых отношениях— центральная проблема современной трофологии. Теоретическое и практическое значение данной проблемы трудно переоценить. Даже простое перечисление вопросов, входящих в нее и в значительной степени определяемых ею, заняло бы слишком много места. Поэтому мы укажем лишь на наиболее разработанные направления современного учения о косвенных пищевых отношениях.[ . ]
Расс м а тр и в а е м а я смешанная популяция характеризуется, во-первых, значительным повышением показателей пищевых отношений в результате совместного питания особей двух ¡видов, -во-вторых, более активным воздействием карпов на золотых рыбок-, чем этих последних на карпов и, в-третьих, одновременным сдвигом как общих рационов, так и их качественного состава (рис. 49,Л).[ . ]
Эта схема, представляя, казалось бы, стройную картину, страдает определенной погрешностью. Например, конкурентные пищевые отношения могут быть как в пределах одного вида, так и между двумя или многими видами, причем межвидовая борьба . за пищу, как будет показано, является более интенсивной, чем внутривидовая. С другой стороны, прямая борьба с организованными врагами иногда наблюдается и внутри одного вида (например в случае каннибализма).[ . ]
В основном встречается в местах с каменистым дном (курья Зыряновская, северная часть озера). В уловах чаще попадается весной и осенью. Это тугорослый вид. В пищевом отношении является конкурентом других видов рыб, потребляя ценные пищевые компоненты. Также наносит ущерб ихтиофауне, поедая икру.[ . ]
Экологическое взаимодействие двух видов, из которых один является хищником, а другой — жертвой, за последние годы было предметом обширных исследований как в отношении накопления эмпирического материала, полученного в природных и в экспериментальных условиях, так и по линии теоретического, главным образом, математического анализа явления. Вместе с тем, следует признать, что закономерности пищевых отношений разных видов животных даже в предельно простом случае, когда популяция одного вида служит пищевым материалом для представителей другого, в значительной части остаются не разъясненными и даже просто выпавшими по тем или иным причинам из круга проблем, уже подвергнутых изучению.[ . ]
Пища — важнейший экологический фактор. Ее качество и количество способны изменять плодовитость, продолжительность жизни, развитие и смертность живых существ. Помимо этого, разнообразие пищевых рационов лежит в основе многочисленных морфологических, физиологических и экологических адаптаций. Действительно, большинство жизненных приспособлений и функций любых видов организмов так или иначе связаны с питанием. Ими являются приспособления растений к почвам, растительноядных животных к поискам корма, хищников — к захвату добычи. Весьма тонки взаимные приспособления цветов и их опылителей, плодов и распространителей семян, даже между паразитами и их хозяевами, между хищниками и их жертвами.[ . ]
Таким образом, картофель, давая большую продукцию сухой массы и крахмала с гектара, представляет очень ценную культуру среди полевых растений, особенно возделываемых для технических целой; в пищевом отношении он может лишь частично заменить зерновые хлеба.[ . ]
Последней по времени является общая концепция Лысенко [51] по вопросам межвидовой и внутривидовой конкуренции. Построенная на основании теоретического обобщения агробиологического материала, эта концепция иногда используется и при анализе пищевых отношений животных.[ . ]
Формулы (6. 1.4) и (6. 1. 5) показывают, что эффективность передачи энергии в трофической цепи и соотношение биомасс популяций сообщества зависят от многих параметров. Наиболее существенно на увеличении У2/Р1 сказывается рост коэффициента усвоения пищи (и2), однако в природных условиях эта величина довольно стабильна. Доступность кормовых объектов (£12) и степень напряженности пищевых отношений (а12 и ¡312) также влияют на энергетическую эффективность популяции в сторону ее увеличения. Что касается интенсивности промысла (то ), то с увеличением этого параметра степень его влияния на рост вылова (У2) не прерывно падает — это связано с уменьшением биомассы В2 или сокращением промысловых запасов.[ . ]
Чрезвычайно высокая интенсивность потоков вещества из неорганической природы в живые тела давно привела бы к полному исчерпанию запасов необходимых для жизни соединений, то есть биогенных элементов. Но этого не происходит, и жизнь не прекращается, так как указанные элементы постоянно возвращаются в окружающую среду. И происходит это благодаря биоценозам, в которых в результате пищевых отношений между видами синтезированные растениями сложные органические вещества превращаются в конце концов в такие простые соединения, как диоксид углерода, вода, ряд элементов, которые могут быть снова использованы растениями в процессе фотосинтеза. Так возникает биологический круговорот вещества. Следовательно, биоценоз, будучи и сам по себе сложной системой живых организмов, является частью еще более сложной системы. В последнюю, помимо живых организмов, входит и их неживое окружение, которое содержит различные вещества и энергию, необходимые для развития и обеспечения жизнедеятельности.[ . ]
Рыбы частиковых пород (елец, плотва, ерш и др.), конкурируя с сиговыми во время их преднерестового нагула, значительно ухудшают его условия, выедая запасы кормовых организмов не только в соровой системе, но и в других участках поймы и русла рек (особенно в низовьях). Об этом говорят и наши наблюдения, и результаты исследований прошлых лет (Москаленко, 1958). Данному обстоятельству должно уделяться особое внимание в годы с относительно низким уровнем весенне-летнего паводка, когда пищевые отношения рыб-конку-рентов из-за бедности ресурсов кормовых объектов обостряются необычайно, что в конечном итоге весьма отрицательно сказывается не столько на линейном и весовом приростах, упитанности производителей сиговых рыб, сколько на своевременности созревания их половых продуктов, массовости нереста и плодовитости — основных показателей, определяющих объем и темпы естественного воспроизводства популяций изученных нами видов рыб.[ . ]
Мы отдаем себе отчет, что принципиально иная трактовка общеэкологических положений должна быть обоснована более солидно, чем это возможно сделать на основании данных, имеющихся в нашем распоряжении. И если мы решаемся, может быть с несколько подчеркнутой категоричностью, говорить о вопросах, по существу достаточно спорных, то делаем это из стремления указать на возможность нового подхода и нового толкования некоторых основных экологических понятий. При анализе явлений косвенных пищевых отношений мы не касаемся вопросов эволюционного порядка. Выше неоднократно подчеркивалось, что основной нашей задачей, определившей весь характер работы, является попытка понять механизм пищевой конкуренции.[ . ]
Под кондицией обычно понимают состояние упитанности животного, обусловленное главным образом его кормлением, содержанием и характером использования. В отличие от конституции, определяемой в значительной мере наследственностью и являющейся в некоторой степени пожизненным свойством животного, его кондиция в течение жизни и даже в течение одного года может меняться. Существенная особенность животных, в значительной мере определяющая их кондиции, заключается в их способности накапливать резервы питательных веществ. Животные с достаточными и разносторонними резервами отличаются более крепким здоровьем, большей устойчивостью против неблагоприятных влияний среды, лучше используют корм и дают более ценную в пищевом отношении продукцию.[ . ]
§ 9. Законы и следствия пищевых отношений
Вспомните
Пищевые
связи
Пищевые отношения не только обеспечивают энергетические потребности организмов. Они играют в природе и другую важную роль — удерживают виды в сообществах, регулируют их численность и влияют на ход эволюции. Пищевые связи чрезвычайно разнообразны.
Типичные хищники тратят много сил на то, чтобы выследить добычу, догнать ее и поймать (рис. 40). У них развито специальное охотничье поведение.

Рис. 40.Гепард в погоне за добычей
Им надо много жертв в течение жизни. Обычно это сильные и активные животные.
Паразиты всю жизнь проводят в одном или двух, реже — трех хозяевах. Они живут в условиях избытка пищи, которую не надо активно добывать, и используют хозяев как среду своего обитания. У них упрощено строение и ослаблены связи с внешним миром.
Животные-собиратели тратят энергию на поиск семян или насекомых, т. е. мелкой добычи. Овладение найденным кормом для них не представляет труда. У них развита поисковая активность, но нет охотничьего поведения.
Пасущиеся виды не тратят много сил на поиск корма, обычно его достаточно много вокруг, и основное время у них уходит на поглощение и переваривание пищи.
В водной среде широко распространен такой способ овладения пищей, какфильтрация, на дне — заглатывание и пропускание через кишечник грунта вместе с пищевыми частицами.
Последствия пищевых связей наиболее ярко проявляются в отношениях хищник — жертва (рис. 41).
Если хищник питается крупными, активными жертвами, которые могут убегать, сопротивляться, прятаться, то в живых остаются те из них, кто делает это лучше других, т. е. имеет более зоркие глаза, чуткие уши, развитую нервную систему, мускульную силу. Таким образом, хищник ведет отбор на совершенствование жертв, уничтожая больных и слабых. В свою очередь, и среди хищников тоже идет отбор на силу, ловкость и выносливость. Эволюционное следствие этих отношений — прогрессивное развитие обоих взаимодействующих видов: и хищника, и жертвы.
Если же хищники питаются малоактивными либо мелкими, не способными сопротивляться им видами, это приводит к другому эволюционному результату. Погибают те особи, которых хищник успевает заметить. Выигрывают менее заметные или чем-то неудобные для захвата жертвы. Так осуществляется естественный отбор на покровительственную окраску, твердые раковины, защитные шипы и иглы и другие орудия спасения от врагов. Эволюция видов идет в сторону специализации по этим признакам.
Самый существенный результат трофических взаимосвязей — сдерживание роста численности видов. Существование пищевых отношений в природе противостоит геометрической прогрессии размножения.
Для каждой пары видов хищника и жертвы результат их взаимодействия зависит прежде всего от их количественных соотношений. Если хищники ловят и уничтожают своих жертв примерно с той же скоростью, с какой эти жертвы размножаются, то они могут сдерживать рост их численности. Именно такие результаты этих взаимосвязей чаще всего характерны для устойчивых природных сообществ. Если скорость размножения жертв выше, чем скорость поедания их хищниками, происходит вспышка численности вида. Хищники уже не могут сдерживать его численность. Это тоже временами встречается в природе. Обратный результат — полное уничтожение жертвы хищником — в природе очень редок, а в экспериментах и в нарушенных человеком условиях встречается чаще. Связано это с тем, что с падением численности какого-либо вида жертв в природе хищники переключаются на другую, более доступную добычу. Охота только за редким видом отнимает слишком много энергии и становится невыгодной.

В первой трети нашего века было открыто, что отношения хищник — жертва могут быть причиной регулярных периодических колебаний численности каждого из взаимодействующих видов. Это мнение особенно окрепло после результатов исследований русского ученого Г. Ф. Гаузе. В своих экспериментах Г. Ф. Гаузе изучал, как изменяется в пробирках численность двух видов инфузорий, связанных отношениями хищник — жертва (рис. 42). Жертвой был один из видов инфузорий-туфелек, питающийся бактериями, а хищником — инфузория-дидиниум, поедающая туфелек.
Вначале численность туфельки росла быстрее, чем численность хищника, который вскоре получил хорошую кормовую базу и тоже стал быстро размножаться. Когда скорость поедания туфелек сравнялась со скоростью их размножения, рост численности вида прекратился. А так как дидиниумы продолжали ловить туфелек и размножаться, скоро выедание жертв намного превысило их пополнение, количество туфелек в пробирках начало резко снижаться. Спустя некоторое время, подорвав свою кормовую базу, прекратили деление и начали погибать дидиниумы. При некоторых модификациях опыта цикл повторился сначала. Беспрепятственное размножение оставшихся в живых туфелек вновь увеличило их обилие, а вслед за ними пошла вверх и кривая численности дидиниумов. На графике кривая численности хищника следует за кривой жертвы со сдвигом вправо, так что изменения их обилия оказываются несинхронны.
Таким образом было доказано, что взаимодействия хищника и жертвы могут при известных условиях приводить к регулярным циклическим колебаниям численности обоих видов. Ход этих циклов можно рассчитать и предсказать, зная некоторые исходные количественные характеристики видов. Количественные законы взаимодействия видов в их пищевых связях очень важны для практики. В рыболовстве, добыче морских беспозвоночных, пушном промысле, спортивной охоте, сборе декоративных и лекарственных растений — везде, где человек уменьшает в природе численность нужных ему видов, он с экологической точки зрения выступает по отношению к этим видам в роли хищника. Поэтому важно уметь предвидеть последствия своей деятельности и организовать ее так, чтобы не подорвать природные запасы.

Г.Ф. Гаузе (1910 -1986) 'российский ученый
В рыболовстве и промысле необходимо, чтобы при снижении численности видов нормы промысла также уменьшались, как это бывает в природе, когда хищники переключаются на более легко доступную добычу (рис. 43).
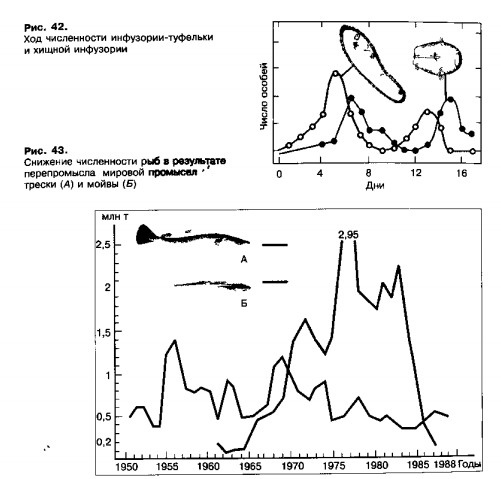
Если же, наоборот, стремиться всеми силами добывать сокращающийся вид, он может не восстановить свою численность и прекратить свое существование. Таким образом, в результате перепромысла, по вине людей уже исчез с лица Земли ряд видов, бывших когда-то очень многочисленными: американские бизоны, европейские туры, странствующие голуби и другие.
При случайном или намеренном уничтожении хищников какого- либо вида сначала возникают вспышки численности его жертв. Это тоже приводит к экологической катастрофе либо в результате подрыва видом собственной кормовой базы, либо — распространения инфекционных заболеваний, которые часто бывают гораздо более губительны, чем деятельность хищников. Возникает явление экологического бумеранга, когда результаты оказываются прямо противоположными начальному направлению воздействия. Поэтому грамотное использование природных экологических законов — основной путь взаимодействия человека с природой.

• Примеры и дополнительная информация
2. Мировой промысел трески происходил в значительной мере стихийно и не был обоснован биологическими характеристиками. Общая добыча достигала 1,4 млн т в год. Это оказалось значительно больше, чем могло быть воспроизведено, поэтому и численность трески, и ее добыча упали в 7—10 раз. Когда в Баренцевом море стадо трески пришло в упадок (70—80-е гг.), резко возросла численность мойвы — главной жертвы трески. Рыбаки переключились на эту рыбку, вылавливая примерно две трети ее общей массы. В результате перевылова численность мойвы также упала. Треска, как и все хищные рыбы, питается всеми мелкими рыбами, включая собственных мальков. При малочисленности мойвы она стала выедать свою молодь, поэтому стадо потеряло возможность восстановиться.
3. В ходе эволюции у жертв вырабатываются самые различные приспособления к защите от хищников. Например, у мельчайших водных коловраток в присутствии других, хищных коловраток вырастают длинные шипы панциря.
Эти шипы сильно мешают хищникам заглатывать жертвы, так как буквально встают у них поперек глотки. Такая же защита возникает у мирных рачков дафний — против других хищных рачков. Хищник, захватив дафнию, перебирает ее своими ножками и переворачивает, чтобы выесть с мягкой брюшной стороны. Шипы мешают ему, и добыча часто теряется. Выяснилось, что у жертв шипы вырастают в ответ на присутствие в воде продуктов обмена веществ хищников. Если врагов в водоеме нет, шипы у жертв не появляются.
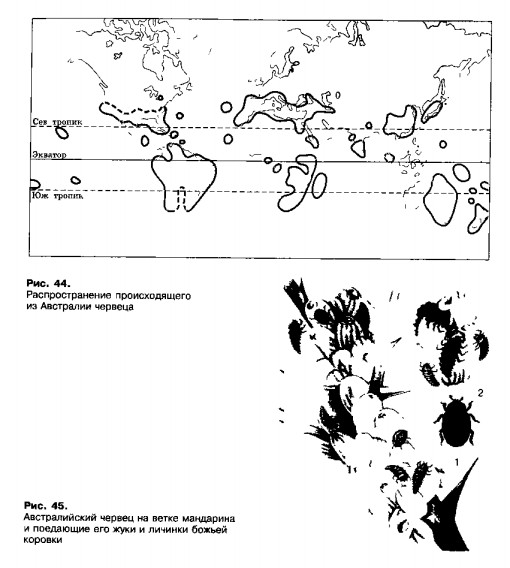
4. Один из первых примеров успешного применения хищника для подавления численности вредителя — использование божьей коровки родолии в борьбе с австралийским желобчатым червецом (рис. 44, 45).
Этот червец— малоподвижное насекомое, сосущее цитрусы, в 1872 г. случайно был занесен в Калифорнию, где у него не было природных врагов. Он быстро размножился и стал опасным вредителем, из-за которого садоводы терпели огромные убытки. Для борьбы с червецом из Австралии ввезли его естественного врага — мелкую божью коровку родолию. В 1889 г. около 10 тыс. жуков были расселены по сотням садов на юге Калифорнии. Уже через несколько месяцев зараженность деревьев червецом резко упала. Коровка прижилась в Калифорнии, и массового размножения червецов больше не наблюдалось. Этот успех повторился в пятидесяти странах мира, вАзде, где родолию выпускали против желобчатого червеца. Родолия более чувствительна к ядохимикатам, чем червец! Поэтому там, где цитрусовые обрабатывались ядами против других вредителей, численность червеца вскоре достигала гигантских масштабов.
5. Рыжие лесные муравьи питаются многими видами беспозвоночных животных, но основу их добычи всегда составляют наиболее массовые виды. В период вспышки численности вредителей леса муравьи питаются преимущественно ими. Подсчитано, что в сибирских лесах обитатели одного крупного муравейника уничтожают задень до 100 тыс. личинок малого елового пилильщика, 10— 12 тыс. бабочек серой лиственничной листовертки. Это значит, что при наличии 5—8 крупных муравейников на гектар можно не беспокоиться о поражении деревьев этими вредителями, муравьи сдержат рост их численности.

1. Всегда ли птицы, привлеченные в древесные насаждения искусственными дуплянками, могут снизить численность вредных насекомых?
2. Создавая математическую модель изменения численности хищника и жертвы, А. Лотка и В. Вольтерра допустили, что количество хищников зависит только от двух причин: числа жертв (чем больше кормовая база, тем интенсивнее размножение) и скорости естественной гибели хищников. При этом они понимали, что сильно упростили отношения, имеющиеся в природе. Укажите, в чем заключается это упрощение.
3. Лось — самый крупный современный олень. Живет в лесных районах, питается порослью лиственных деревьев и высокотравьем. В начале XX столетия численность его в Европе сильно сократилась. Однако начиная с 20-х гг. и особенно в 40-е гг. она начала восстанавливаться в результате охраны лося, омоложения лесов и сокращения числа волков. Укажите, какие именно пищевые связи сыграли роль в восстановлении вида. Почему в настоящее время разрешена умеренная охота на лося?
1. В одном из опытов мелкие насекомые-паразиты искали и заражали своими яйцами куколки-пупарии комнатной мухи. В разных вариантах опыта 40 паразитам предлагали разное число куколок: 25, 50, 100, 200 и 300. Число зараженных куколок оказалось соответственно 18, 32, 48, 54, 62. Начертите график числа зараженных пупариев, приходящихся на одного паразита, при увеличении численности жертв. Рассчитайте, какую долю куколок мух заражают паразиты при разной численности жертв. В каком из вариантов опыта они наиболее эффективно влияли на численность куколок?
2. В одном из лесных хозяйств учитывали гусениц хвойной листовертки — вредителя хвойных пород, а среди них — число здоровых и зараженных паразитами. По полученным данным начертите графики изменения общей численности гусениц и числа зараженных. Сравните и объясните ход кривых. Как зависит доля зараженных гусениц от общей численности хозяина листовертки? Могут ли паразиты сдерживать рост численности листовертки?

Темы для дискуссий.
1. Хотя расчеты и опыты свидетельствуют, что в природе между каждой парой видов хищник — жертва могут возникать колебательные циклы, в природе такие циклы наблюдаются редко. Почему?
2. В дальневосточных лесах ведут интенсивный промысел ценного лекарственного растения — женьшеня Вид находится на грани исчезновения. Какие меры вы приняли бы для его сохранения? Какое отношение к этим мероприятиям имеет понимание связей хищник — жертва?
3. Долгое время у нас в стране поощрялась охота на волков и за каждого убитого зверя выдавали премию. Затем охоту на волка полностью запретили. В настоящее время в ряде районов этот запрет вновь снят и часть волков разрешают отстреливать. Как вы думаете, чем можно объяснить такую непоследовательность в распоряжениях природоохранительных органов?
4. В природе отношения хищник — жертва между конкретными видами существуют миллионы лет. Современный человек, вступая в такие же отношения с видами дикой природы (охота, рыбная ловля, сбор лекарственных и пищевых растений, цветов и т. п.), быстро подрывает их численность. Почему так происходит? Может ли изменить эти результаты знание и применение экологических правил?
5. Предположим, что вы должны установить норму вылова ценного вида рыб. Какими сведениями об этом виде вы должны располагать, чтобы рассчитать эту норму? Что произойдет в случае завышения нормы вылова? ее занижения?
Чернова Н. М., Основы экологии: Учеб. дня 10 (11) кл. общеобразоват. учеб. заведений/ Н. М. Чернова, В. М. Галушин, В. М. Константинов; Под ред. Н. М. Черновой. — 6-е изд., стереотип. — М.: Дрофа, 2002. — 304 с.
Учебники и книги по всему предметам, домашняя работа, онлайн библиотеки книжек, планы конспектов уроков по экологии, рефераты и конспекты уроков по экологии для 10 класса скачать
Если у вас есть исправления или предложения к данному уроку, напишите нам.
Если вы хотите увидеть другие корректировки и пожелания к урокам, смотрите здесь - Образовательный форум.
Читайте также: