
Доклад на тему половой отбор
Обновлено: 17.05.2024
Любовь Петровна Малиновская — аспирант Новосибирского государственного университета, научный сотрудник лаборатории молекулярной цитогенетики птиц Института цитологии и генетики СО РАН. Область научных интересов — молекулярная цитогенетика птиц.
Анна Александровна Торгашёва — кандидат биологических наук, старший научный сотрудник, заведующая лабораторией молекулярной цитогенетики птиц Института цитологии и генетики СО РАН. Научные интересы связаны с молекулярной биологией мейоза, эволюционной цитогенетикой животных.
Виген Артаваздович Геодакян на съезде Вавиловского общества генетиков и селекционеров, проходившем в Московском государственном университете имени М. В. Ломоносова в июне 2009 г. Фото А. А. Торгашёвой
В этой статье мы хотим рассказать историю, которая, на первый взгляд, противоречит идеям Геодакяна. И на второй — тоже. Но как жаль, что его больше нет с нами. Почему-то нам кажется, что он и в ней нашел бы подтверждение своей теории. С теплыми воспоминаниями и светлой грустью мы посвящаем эту статью его доброй памяти.
Неправильное правило Хаксли — Холдейна
Гетерогаметный — значит, имеющий разные половые хромосомы: XY у самцов млекопитающих и двукрылых (включая комаров и муху дрозофилу) или ZW у самок птиц и бабочек. Соответственно, гомогаметным полом оказываются самки ХХ у первых и самцы ZZ у последних. т.е. правило Хаксли — Холдейна говорило не о самцах и самках, а о гетеро- и гомогаметном полах. В чем здесь фокус? Почему нужно блокировать рекомбинацию у гетерогаметного пола?
Рекомбинация наряду с мутациями служит мощным генератором генетической изменчивости. Но между ними есть большая разница. Мутационный процесс — это своего рода проявление энтропии на молекулярно-генетическом уровне. За счет неизбежных ошибок удвоения и ошибочной репарации (исправления) этих ошибок в ДНК возникают мутации — искажения генетической информации, упорядоченной предшествующим отбором, т.е. мутации — это неизбежное нарастание хаоса.
Рекомбинация — это хорошо упорядоченная, запрограммированная и затратная генерация хаоса (рис. 1). В ходе рекомбинации в ДНК запрограммированно вносится множество вполне фатальных повреждений — двунитевых разрывов. Эти разрывы затем репарируются: иногда правильно — с восстановлением исходной информации, но чаще неправильно — не по той цепи, которой нужно, а то и вовсе крест-накрест. Эти перекресты приводят к тому, что родительские комбинации аллелей, которые, между прочим, позволили родителям дожить до брака и приступить к производству потомков, разрушаются. Вместо них возникают новые, ранее не опробованные комбинации, которые вполне могут оказаться как хуже, так и лучше родительских [4].
Рис. 1. Рекомбинация гомологичных хромосом [6]. Розовым цветом обозначена ДНК материнской хромосомы, синим — отцовской, зеленым — ДНК, достроенная в ходе репарации разрывов
Таким образом, от правила Хаксли — Холдейна осталось только то, что самцы отличаются от самок. Да и то оказалось, что половые различия в уровне рекомбинации сильно варьируют и в размахе (когда сильные, когда слабые, когда и вовсе отсутствуют), и в знаке (чаще всего больше у самок, но бывает и у самцов). И если эти различия есть, то они никак не связаны с гетерогаментостью. И все же, почему рекомбинация, как правило, выше у самок?
Половой отбор как универсальное объяснение всего на свете
Половой отбор — это красивая и остроумная теория. Но при чем здесь рекомбинация? Как пониженная рекомбинация у самцов может им помочь в борьбе за самок? Как может самка определить рекомбинационные способности самца и зачем ей это нужно? Уровень рекомбинации — это такой признак самца, который можно оценить, только скрестившись с ним и посмотрев на разнообразие полученных потомков. Но если потомки уже получены, значит выбор уже сделан и оценка рекомбинации уже никому не нужна. Но даже если бы самка могла определить способности самца заранее, почему ей должны нравиться мало рекомбинирующие самцы?
Рис. 2. Половой отбор убегающего признака и рекомбинация. А — аллель, контролирующий у самок предпочтение самцов с длинным хвостом; а — нет предпочтений у самок; Т — аллель, контролирующий у самцов развитие длинного хвоста; t — нормальный хвост у самцов
От моделей к фактам
Теорий, которые объясняют половые различия в рекомбинации (гетерохиазмию), более чем достаточно. С фактами ситуация гораздо хуже. Данных мало, и они довольно противоречивы. Теоретики по определению не занимаются экспериментами, а используют в своих моделях данные, полученные другими людьми и для других целей. В итоге довольно часто им приходится сравнивать, фигурально выражаясь, ужа и ежа просто потому, что других данных нет.
С птицами, которые интересны тем, что у них гетерогаметный пол — самки, ситуация не лучше. Данные по рекомбинации у обоих полов получены для девяти видов, которых тоже разделяют десятки миллионов лет независимой эволюции и отличают друг от друга множество особенностей их биологии. Половые различия по рекомбинации у птиц непредсказуемо варьируют: у близких видов они могут сильно различаться по знаку и размаху [10]. Поэтому мы решили получить собственные данные по рекомбинации для двух видов птиц, которые разошлись относительно недавно в ходе эволюции и сходны по всем особенностям биологии, кроме одной: у одного вида нет полового отбора, а у другого он доказан наблюдениями и экспериментами.
Две ласточки
Мы сравнили два вида ласточек — деревенскую ласточку (Hirundo rustica) и бледную береговушку (Riparia diluta) по параметрам рекомбинации у самцов и самок [11]. Современные цитологические методы позволяют выявлять точки рекомбинации прямо на препаратах хромосом (рис. 3).
Рис. 3. Микрофотография сперматоцита деревенской ласточки после иммуноокрашивания белков осей хромосом (красный), центромер (синий) и рекомбинационных узелков (зеленый) [10]. Цифрами обозначены макрохромосомы, буквами — половые хромосомы. Фото Л. П. Малиновской
Почему мы выбрали именно эти виды? Потому что они состоят в относительно близком родстве (около 16 млн лет дивергенции) и довольно похожи в отношении экологии, образа жизни и систем размножения. У них сходны районы миграции и гнездования. Оба вида образуют большие колонии. Оба практикуют относительно строгую моногамию, совместное высиживание яиц и выкармливание потомков.
Различия между ними состоят в том, что у береговушек практически невозможно отличить самцов от самок, в то время как самцы европейского подвида деревенской ласточки (ареал которого доходит до Байкала) отличаются от самок большей яркостью оперения и, главное, большей длиной хвостовых направляющих перьев (косиц). По этому признаку у деревенской ласточки идет довольно интенсивный половой отбор. Самки отдают безусловное предпочтение самцам с более длинными косицами. Такие самцы оставляют больше потомков. Они раньше других находят себе пару, чаще других вступают во внебрачные отношения с соседними самками, а их собственные самки чаще производят две кладки в сезон. Эти различия в репродуктивном успехе были подтверждены как прямыми наблюдениями, так и экспериментами с искусственным удлинением или укорочением косиц [11].
Брачные пары деревенской ласточки (слева) и бледной береговушки. Фото Е. Шнайдер
Рис. 4. Половые различия по числу (вверху) и распределению (внизу) рекомбинационных узелков на хромосомах деревенской ласточки и бледной береговушки [10]
Рекомбинация на концах хромосом важна и полезна для правильного их расхождения в мейозе, но мало что дает для перетасовки аллелей. В то время как рекомбинация в средних частях хромосом дает широкий и разнообразный набор новых аллельных комбинаций в потомстве. За счет этих двух особенностей — более высокой частоты рекомбинации вообще и в средних частях хромосом в частности — самки деревенской ласточки разрушают старые комбинации аллелей и создают новые гораздо быстрее, чем самцы того же вида. При этом у бледной береговушки — морфологически мономорфного вида — мы не обнаружили значительных половых различий ни по числу рекомбинационных узелков, ни по их распределению. И, что особенно интересно, они очень похожи по этим признакам на самцов деревенской ласточки.
Логично допустить, что у общего предка наших двух видов ласточек самцы и самки были похожи друг на друга и по морфологии, и по уровню рекомбинации. Затем половой отбор у деревенских ласточек привел к возникновению полового диморфизма по обоим этим признакам. Но почему это произошло не за счет снижения рекомбинации у самцов, а за счет ее повышения у самок? Можно думать, исходно низкий уровень рекомбинации у обоих полов позволил возникнуть и закрепиться неравновесию по сцеплению между аллелями, контролирующими проявление вторичных половых признаков у самцов, и предпочтение к этим признакам у самок. Поскольку половая разница в рекомбинации у деревенской ласточки наиболее выражена в макрохромосомах, можно предположить, что именно там находятся гены, законсервированные половым отбором. Сильный отбор по тем районам хромосом, где они находятся, должен вести к накоплению вредных мутаций в этих и соседних районах. Увеличение частоты рекомбинации в макрохромосомах в женском мейозе может быть направлено на очищение от этих мутаций. И кроме того, вспомните, что самки любят самцов не просто с длинным хвостом, но с самым длинным. Отсюда следует, что самки должны интенсивно тасовать свои аллели, надеясь снабдить своих сыновей более длинными хвостами. Так что наши данные не так уж и противоречат идеям Геодакяна. Да, сами самки, по его теории, должны быть более мономорфными, чем самцы. Но при этом они могут генерировать более разнообразное потомство, чтобы их дочерям в следующем поколении было, из кого выбирать.
Конечно, эта гипотеза весьма умозрительна. Нам нужно больше данных о величине и направлении гетерохиазмии среди близких видов птиц с явным половым отбором и без него. Нам нужны данные о локализации генов, контролирующих вторичные половые признаки самцов и особенности половых предпочтений у самок. И нам очень не хватает Вигена Артаваздовича, чтобы обсудить с ним эти данные. Он бы обязательно придумал им правильное объяснение.
Работа выполнена при поддержке Российского научного фонда (проект 20-64-46021).
Литература
1. Геодакян В. А. Эволюционная роль половых хромосом (новая концепция) // Генетика. 1998; 34(8): 1171–1184.
2. Haldane J. B. S. Sex-ratio and unisexual sterility in hybrid animals // J. Genet. 1922; 12: 101–109. DOI: 10.1007/BF02983075.
3. Huxley J. S. Sexual difference of linkage in Gammarus chevreuxi // J. Genet. 1928; 20: 145–156. DOI: 10.1007/BF02983136.
4. Bishop D. K., Zickler D. Early decision; meiotic crossover interference prior to stable strand exchange and synapsis // Cell. 2004; 117(1): 9–15. DOI: 10.1016/s0092-8674(04)00297-1.
5. Sardell J. M., Kirkpatrick M. Sex differences in the recombination landscape // Am. Nat. 2020; 195: 361–379. DOI: 10.1086/704943.
6. Миллер Дж. Соблазняющий разум: Как выбор сексуального партнера повлиял на эволюцию человеческой природы. М., 2020.
7. Fisher R. A. The Genetical Theory of Natural Selection. Oxford, 1930.
8. Trivers R. Sex Differences in Rates of Recombination and Sexual Selection // The Evolution of Sex: an Examination of Current Ideas / Michod R. E. and B. R. Levin (eds). Sunderland, 1988; 270–286.
9. Mank J. E. The evolution of heterochiasmy: The role of sexual selection and sperm competition in determining sex-specific recombination rates in eutherian mammals // Genet. Res. 2009; 91: 355–363. DOI: 10.1017/S0016672309990255.
10. Malinovskaya L. P., Tishakova K., Shnaider E. P., Borodin P. M., Torgasheva A. A. Heterochiasmy and sexual dimorphism: The case of the barn swallow (Hirundo rustica, Hirundinidae, aves) // Genes (Basel). 2020; 11(10): 1119. DOI: 10.3390/genes11101119.
11. Møller A. P. Sexual Selection and the Barn Swallow. Oxford, 1994.
** См. например: Бородин П. М. Генетическая рекомбинация в свете эволюции // Природа. 2007. № 1. С. 14–22.
- Для учеников 1-11 классов и дошкольников
- Бесплатные сертификаты учителям и участникам
Половой отбор
у человека и животных
Борьба за существование приводит к гибели значительного числа особей в каждом поколении любого вида и выборочном участии особей в размножении.
Частный случай естественного отбора - половой отбор, обеспечивающий развитие признаков, связанных с функцией размножения. Он выражается в соперничестве особей одного пола (обычно мужского) за спаривание с особями другого пола. В результате полового отбора в процессе эволюции развились вторичные половые признаки. Особи с резко выраженными вторичными половыми признаками легче привлекают внимание особей другого пола, что ведёт к их преимущественному размножению. В результате полового отбора сохраняются признаки бесполезные для выживания вида, но привлекательные для самок. Например, тяжёлые разветвлённые рога у оленей, длинный тяжёлый хвост у самцов некоторых птиц. Половой отбор отсутствует у многих видов животных и у растений, он выражен у птиц и млекопитающих в связи с развитием нервной системы и её сигнальной деятельности.
Половой отбор у людей
Половой отбор у людей имеет свои отличительные особенности.
Предки современных женщин не знали проблемы менструальной напряженности. Раньше женщина непрерывно находилась в состоянии беременности и менструальная напряжённость возникала 10-20 раз за всю жизнь, в то время как у современной женщины - 12 раз в год.
Половой отбор у людей связан с социальным расслоением. Сильные имеют доступ к власти, благам цивилизации и покупают красивых женщин. Самые агрессивные, беспринципные мужчины обладают самыми красивыми женщинами. Появляются дети с высокими стартовыми возможностями.
Слабые имеют минимум социальных благ, нехватку денег. Они спиваются и теряют квартиры. Дети обычно следуют по пути родителей.
Современная женщина не хочет производить потомство по воле природных ритмов, а реализует своё право на свободу через регулирование рождаемости. Самый жестокий способ - аборт. В начале 20 века изобрели надёжный способ контрацепции - презерватив, затем гормональные контрацептивы. Отделение секса от размножения предоставило женщине большие возможности для профессиональной реализации, снизило финансовую зависимость от мужчины. Вместе с тем, свобода секса способствовала резкому подъёму урогенитальных инфекций.
Половой отбор накладывает отпечаток на динамику старения мужчин и женщин. Мужчина может стать отцом даже в преклонном возрасте, поэтому стареет более плавно. Старение женщины протекает гораздо драматичнее.
Если существует столкновение интересов родителей и детей, гены которых на 50% одинаковы, то насколько более жестким должен быть конфликт между супругами, вовсе не связанными родством?. Все, что есть между ними общего, - это их генетические вклады в одних и тех же детей, по 50% от каждого из родителей. Поскольку и отец, и мать заинтересованы в благополучии разных половинок одних и тех же детей, обоим может быть выгодно кооперироваться для выращивания этих детей. Если, однако, одному из родителей удастся вложить в каждого ребенка меньше той доли ценных ресурсов, которую положено внести ему (или ей), он окажется в лучшем положении, потому что сможет больше затратить на других детей, зачатых с другим половым партнером, и тем самым распространить большее число своих генов. Таким образом, каждого из партнеров можно рассматривать как индивидуума, который стремится эксплуатировать другого, пытаясь заставить его внести больший вклад в выращивание потомков. Разногласия возникают по поводу того, кто должен нести бремя расходов по выращиванию всех этих детей. Каждый индивидуум хочет, чтобы у него выжило как можно больше детей. Чем меньше ему или ей приходится вкладывать в каждого из своих детей, тем больше детей он или она смогут иметь. Очевидный способ достигнуть этого желаемого положения вещей состоит в том, чтобы склонить своего брачного партнера внести в каждого ребенка больше, чем положенная ему или ей доля ресурсов, а затем предоставить ему или ей свободу заводить других детей с другими партнерами. Такая стратегия была бы желательна для обоих полов, однако осуществить ее самке труднее. Поскольку мать с самого начала вкладывает в ребенка - в форме своего крупного богатого питательными веществами яйца - больше, чем отец, она уже в момент зачатия принимает на себя более серьезные "обязательства" в отношении каждого ребенка, чем это делает отец. Если ребенок гибнет, то она теряет больше, чем отец. Точнее, ей пришлось бы внести больший, чем отцу, вклад в будущем, чтобы родить на замену нового ребенка и довести его до того же возраста. Если бы она попыталась оставить ребенка на отца, а сама ушла бы к другому самцу, то отец мог бы за счет относительно небольших потерь для себя отомстить ей, в свою очередь бросив ребенка. Поэтому, по крайней мере на ранних стадиях развития ребенка, если кто-то кого-то и бросает, то чаще отец уходит от матери, чем наоборот. Кроме того, следует ожидать, что самки будут вкладывать в детей больше, чем самцы, не только в самом начале, но и на всем протяжении развития. Например, у млекопитающих именно самка вынашивает плод в собственном теле, вырабатывает молоко, которым кормит родившегося детеныша, на нее ложится главная часть тягот по его выращиванию и защите. Женский пол находится в положении эксплуатируемого и исходная эволюционная основа для его эксплуатации тот факт, что яйцеклетки крупнее, чем сперматозоиды.
Половой отбор у животных
Половой отбор основан на селективной неравноценности особей одного пола у раздельнополых организмов (обычно у животных). Это особая форма индивидуального отбора, в которой участвуют представители только одного пола (обычно самцы) данной популяции.
Половой диморфизм у животных нередко выражается в различиях по двум группам признаков. Первичные половые признаки — это признаки, непосредственно относящиеся к размножению. Здесь мы не будем их рассматривать. Вторичные половые признаки, имеющиеся у самцов, помогают им отыскивать брачных партнеров. К этим вторичным половым признакам и обращена теория полового отбора.
Дарвин представлял себе половой отбор как процесс, дополняющий более общий и более широко распространенный процесс естественного отбора. Последний, по теории Дарвина, создал адаптивные признаки данного вида в целом, в том числе изначальные приспособления, общие для обоих полов, а также первичные половые различия, непосредственно связанные с размножением. Естественный отбор в первоначальном: дарвиновском смысле не объяснял вторичные половые признаки. Такие признаки, как рога оленя или яркое оперение селезней, не представляют собой приспособлений, благоприятных для вида в целом; не являются они также необходимыми и для размножения. Однако они, по-видимому, повышают вероятность успешности спариваний для тех самцов, которые ими обладают. Теория полового отбора была введена для того, чтобы объяснить развитие таких специальных мужских признаков.
Следует отметить, что эти два типа полового отбора необязательно исключают друг друга. Так, у полигинного бычка Cottus bairdi обнаружен половой отбор обоих типов ( Downhower et al ., 1983*).
Изучение полового отбора начиная с 1871 г. шло извилистым путем. Во времена Дарвина эта тема была предметом разногласий. Уоллес ( Wallace , 1889*) допускал роль полового отбора в развитии признаков, необходимых самцам в драках, но не в развитии демонстрационных признаков. Затем эта проблема была предана забвению. Когда в начале современного периода эволюционных исследований она вновь выступила на сцену в работах Фишара ( Fisher , 1930*) и Гексли ( Huxley , 1938*), ситуация была совершенно иной: те же самые явления рассматривались с других альтернативных, точек зрения и теория полового отбора должка была дебютировать вновь. Этот второй дебют оказался удачным: в настоящее время половой отбор является предметом активных исследований и ему посвящена обширная литература*.
Возникновение всего обширного спектра вторичных половых признаков — результат совместного действия различных селективных процессов, поэтому трудно выделить здесь эффекты одного только полового отбора.
В последние годы делаются попытки распространить концепцию полового отбора на двудомные растения. Эти попытки выражаются в истолковании известных фактов биологии размножения растений в терминах сильно расширенной концепции полового отбора. В частности, указывают на очень большое число пыльцевых зерен по сравнению с числом яйцеклеток и возникающую в результате конкуренцию между этими зернами ( Stephenson , Bertin , 1983; Willson , 1983*). Однако такое положение вещей — результат исходного разделения труда между самцами и самками; это непременное предварительное условие для процесса полового отбора, как уже упоминалось, а не его результат. Явление несовместимости у растений, некоторые типы пыльцы которых успешно прорастают лишь в определённом семязачатке, рассматривается как выбор со стороны особи женского пола ( Stephenson , Bertin , 1983; Willson , Burley , 1983*). В этом случае концепция такого выбора расширяется, охватывая иммунологические реакции организмов, лишенных нервной системы или умственных способностей. Межсамцовый отбор теоретически возможен у двудомных растений. Однако эта проблема в настоящее время далека от решения из-за отсутствия критического подхода, а также необходимых данных.
Признаки самцов, связанные с драками между ними
Роль полового отбора ясно выражена в развитии признаков, связанных с драками между самцами и с явлением доминирования. Для полового отбора этого типа необходимо наличие трёх условий: 1) конкуренция между самцами за самок; 2) генотипические различия между самцами, определяющие их конкурентоспособность в борьбе за самок; 3) репродуктивное преимущество самцов, добившихся успеха, над другими самцами.
Полигиния часто встречается у млекопитающих и иногда у птиц. Среди млекопитающих она наблюдается у оленей, крупного рогатого скота, овец, у большинства антилоп, у слонов, тюленей, морских львов, моржей и павианов; среди птиц — у кур, фазанов и павлинов. У самцов этих животных хорошо развиты вторичные половые признаки в отличие от самцов родственных неполигинных групп. Так, у полигинных кур, фазанов и павлинов самцы гораздо крупнее, драчливее, и оперение у них наряднее, чем у самок, а у моногинных— серой куропатки, грауса и тундряной куропатки — различия между особями разного пола относительно невелики. У моржей и морских львов самцы очень крупные; самцов многих копытных украшают рога; у павианов самцы крупные и агрессивные. В отличие от этого у моногинных волков и некоторых моногинных видов обезьян, а также у ряда кошек, у которых детенышей выращивает в основном мать, и у колониальных, но неполигинных грызунов, самцы и самки почти не отличаются друг от друга по размерам и силе ( Darwin , 1871, гл. 8*).
Поскольку признаки, связанные с драками между самцами, наблюдаются преимущественно в полигинных группах, можно считать, что они возникли в результате полового отбора. Корреляция между диморфизмом по общим размерам и системой спаривания выражена менее ясно, не говоря уже о многочисленных исключениях и осложнениях ( Ralls , 1977*).
У млекопитающих и птиц наблюдается также и другая корреляция — между системой спаривания и вкладом родителей в заботу о потомстве ( Trivers , 1972; Zeveloff , Boyce , 1980*).
У птиц самцы и самки принимают более или менее одинаковое участие в различных аспектах заботы о потомстве, т. е. в постройке гнезда и выкармливании птенцов. Стратегия размножения, характерная для большинства птиц, делает необходимым сотрудничество обеих родительских особей. Это ведет к моногамии, которая широко распространена у птиц. При моногамии, когда у каждого самца есть брачный партнер, конкуренции между самцами за самок не возникает. Соответственно не развиваются и признаки, необходимые самцам в драках и для завоевания доминирующего положения.
Полигинные птицы, для которых характерно доминирование самцов, представляют собой исключение, подтверждающее правило.
У млекопитающих стратегия размножения основана, напротив, на преимущественной роли матери в заботе о потомстве, что обусловлено вынашиванием зародыша во чреве и выкармливанием детеныша собственным молоком. Вклад самцов в заботу о потомстве часто пренебрежимо мал. Свободные от этой заботы самцы получают возможность конкурировать за самок и устраивать гаремы. Это создаёт почву для полового отбора по признакам, связанным с доминированием самцов, которые часто встречаются у млекопитающих. Моногамные млекопитающие с их равноправием между полами — исключение, подтверждающее правило.
Признаки, определяющие привлекательность самцов
Гораздо менее ясна роль полового отбора в развитии демонстрационных признаков самцов, таких, как разного рода украшения, пение и феромоны, в неполигинных группах животных. Названным признакам можно дать различные объяснения, не обязательно связанные с половым отбором. Кроме того, существуют проблемы, касающиеся адекватности самого селективного механизма. Половой отбор по признакам, связанным с украшениями и демонстрационным поведением самцов, подразумевает выбор, осуществляемый самками сугубо индивидуально, а это фактор, не поддающийся количественной оценке.
Отдают ли самки предпочтение самым привлекательным или самым сильным самцам, и постоянны ли они в своих предпочтениях? А у неполигинных животных, у которых число самок и самцов одинаково и они образуют брачные пары, оставят ли самки, спаривающиеся с самцами, которых они сразу предпочли всем остальным, больше потомков, чем те, которым пришлось выбирать вторично? Половой отбор по признакам, определяющим привлекательность самцов, был бы эффективен, если бы все или большинство самок предпочитали самцов какого-то одного типа, но выполняется ли это условие в природе? Что происходит, если самки различаются в отношении предпочитаемого типа самца? Если же для всех или для большинства самок данного вида существует одинаковый стандарт привлекательности самцов, то каким образом этот общий для всего вида стандарт возник впервые?
В некоторых недавних исследованиях обнаружена связь между предпочтениями самок и репродуктивным успехом самцов избираемого типа. У бабочек Colias ( С. eurytheme и С. philodice ) самки делают выбор между самцами на основании кинетики полета. Предпочитаемые самцы достигают наибольшего успеха в спаривании и, по-видимому, обладают также наилучшей способностью к полету ( Watt et al ., 1986*). У лягушки Physalaemus pustalosus в Панаме самки предпочитают определённый тип звуковых сигналов. Издающие такие крики самцы имеют больше шансов на спаривание. Эти успешно спаривающиеся самцы, кроме того, старше и крупнее, так что в этом случае, так же как и у бабочек Colias , по-видимому, играет роль приспособленность ( Ryan , 1980; 1983; 1985*). Самки бычка Cottus bairdi предпочитают спариваться с крупными самцами, причём успешно спаривающиеся самки откладывают более жизнеспособные яйца ( Downhower et al ., 1983*). В этих и в сходных случаях выбор самок основан на признаке, связанном с общей приспособленностью самцов.
Подобные случаи не позволяют объяснить обширный класс демонстрационных признаков самцов, которые, казалось бы, не дают своим обладателям никаких преимуществ и даже могут оказывать пагубное воздействие на их жизнеспособность. Принято считать, что длинные хвосты у самцов некоторых райских птиц снижают их приспособленность. Брачные призывы самцов панамских лягушек повышают вероятность уничтожения их хищниками — летучими мышами ( Trachops cirrhosus ) ( Tuttle , Ryan , 1981*).
Есть основания сомневаться в том, что механизм полового отбора может создавать и действительно создаёт хорошо заметные и не адаптивные демонстрационные признаки самцов. Возникновение этого класса признаков можно, однако, объяснить другим сочетанием селективных сил, действие которых слагается из ряда последовательных стадий. Вначале таким демонстрационным признакам благоприятствует обычный индивидуальный отбор, способствуя ухаживанию и спариванию. Конкретные демонстрационные признаки и ритуалы ухаживания в пределах данного вида, вероятно, могли иметь расовые особенности, как, например, локальные диалекты у некоторых видов птиц. Расовые различия должны были бы вызывать некоторую начальную репродуктивную изоляцию. Дальнейшая дивергенция на видовом уровне обычно должна усиливать эту экологическую изоляцию и лежащие в её основе признаки, как побочный результат такой дивергенции. И наконец, демонстрационные признаки и ритуалы ухаживания дивергентных видов должны дифференцироваться ещё сильнее в результате отбора, направленного на этологическую изоляцию как таковую, с тем чтобы подавить межвидовую гибридизацию.
Эта гипотеза согласуется с той большой ролью, которую играют демонстрационные признаки самца и выбор со стороны самки в этологической изоляции. Выбор самкой брачного партнера в сущности касается межвидовых отношений в не меньшей мере, чем внутривидовых. Самки обычно принимают самцов своего вида, но отвергают самцов с демонстрационными признаками чужого вида. Короче говоря, наличие у самцов неадаптивных демонстрационных признаков можно объяснять как результат видообразования.
Связь между этологической изоляцией, видообразованием и половым отбором была отмечена недавно у рыб семейства Cichlidae из африканских озер и у группы Drosophila sitvestris с Гавайских островов ( Dominey , 1984; Carson , 1986; Mayr , 1988*). Эти авторы считают половой отбор движущей силой в развитии этологической изоляции и видообразовании. Такое объяснение соответствует имеющимся фактам. Равным образом, однако, можно считать, что главным определяющим фактором является видообразование, как это описано выше. Мне, вообще, не кажется необходимым привлекать половой отбор в строгом смысле для объяснения именно этих и других сходных случаев.



Что произошло? Ведь самок никто не отбирал, только самцов!
Именно Рональд Фишер первым понял, что не только брачные украшения самцов, но и вкусы и избирательность самок тоже наследуются и эволюционируют – по тем же правилам, что и остальные признаки (Дарвин предполагал, что это какое-то врожденное чувство прекрасного).
Эта простая мысль сделала теорию полового отбора полностью работоспособной. Однако идеи Фишера о половом отборе тоже не были восприняты научным сообществом. Они почти забылись к тому времени, когда их снова выкопали из забвения представители следующего поколения биологов-теоретиков, сумевшие, наконец, разработать убедительные и понятные математические модели. Лишь с третьей попытки теория полового отбора получила заслуженное признание.

Три важнейших идеи (модели), развитые в рамках теории полового отбора: фишеровское убегание, индикаторы приспособленности, принцип гандикапа.

Допустим, в популяции появился мутантный аллель, влияющий на брачные предпочтения самок. Допустим, самки с этим геном выбирают самых длинноухих самцов. В популяции существует небольшая нейтральная (не влияющая на приспособленность) изменчивость по длине ушей. Самки с мутантным геном поначалу не имеют никакого преимущества, но и особого вреда их странные вкусы им не приносят. Поэтому мутантный аллель имеет шанс за счет дрейфа достичь некоторой заметной частоты в генофонде популяции. Вот тут‑то и вступает в действие механизм “убегания”. Длинноухие самцы получают репродуктивное преимущество, потому что им доступны все самки, а короткоухие самцы могут спариться только с теми самками, у которых нет мутантного аллеля. Длинноухие самцы начинают оставлять больше потомков, чем короткоухие.
Потомство от браков длинноухих самцов с мутантными самками наследует не только “гены длинноухости” (от отца), но и аллель предпочтения длинноухих самцов (от матери). В результате эти аллели начинают распространяться.
Как только длинноухие самцы начинают оставлять в среднем больше потомства, чем короткоухие, самкам становится выгодно выбирать длинноухих партнеров, потому что тогда их сыновья унаследуют длинноухость, привлекут больше самок и оставят больше потомства. В результате те самки, которые предпочитают длинноухих самцов, начинают оставлять больше внуков, то есть получают репродуктивное преимущество.

Бывшая случайная прихоть превращается в полезную адаптацию. Возникает положительная обратная связь, в результате которой в генофонде быстро распространяются гены длинноухости и гены любви к длинноухим.
Длинноухость выгодна, потому что самки любят длинноухих, а любить длинноухих выгодно, потому что выгодна длинноухость. И никому нет дела до того, нужны ли длинные уши зачем‑то еще. В этом и состоит суть фишеровского убегания.
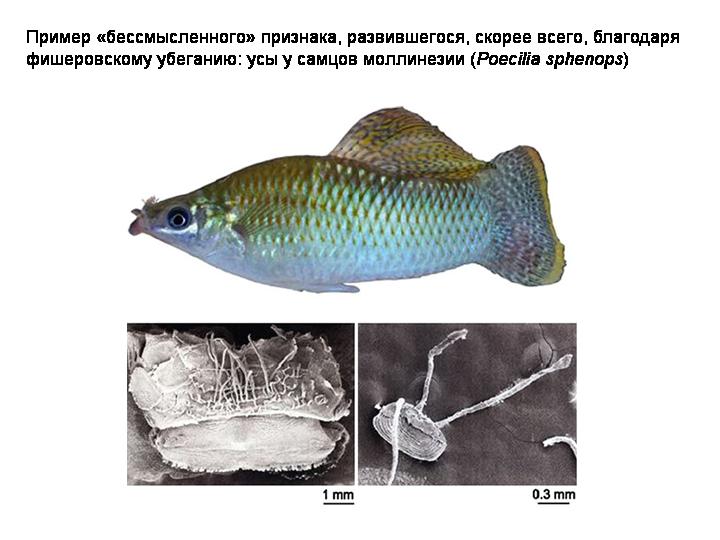



П.О. СПОСОБСТВУЕТ СИМПАТРИЧЕСКОМУ ВИДООБРАЗОВАНИЮ
Половой отбор может способствовать симпатрическое видообразование. Ясно, что без полового отбора и избирательного скрещивания симпатрическое видообразование едва ли возможно. До недавних пор все думали, что для симпатрического видообразования самки двух зарождающихся видов должны иметь разные половые предпочтения. И было не очень понятно, с чего бы вдруг у самок в одной популяции возникли разные вкусы. Но на самом деле, как было недавно показано, половой отбор стимулирует симпатрическое видообразование даже в том случае, если вкусы у всех самок остаются одинаковыми.
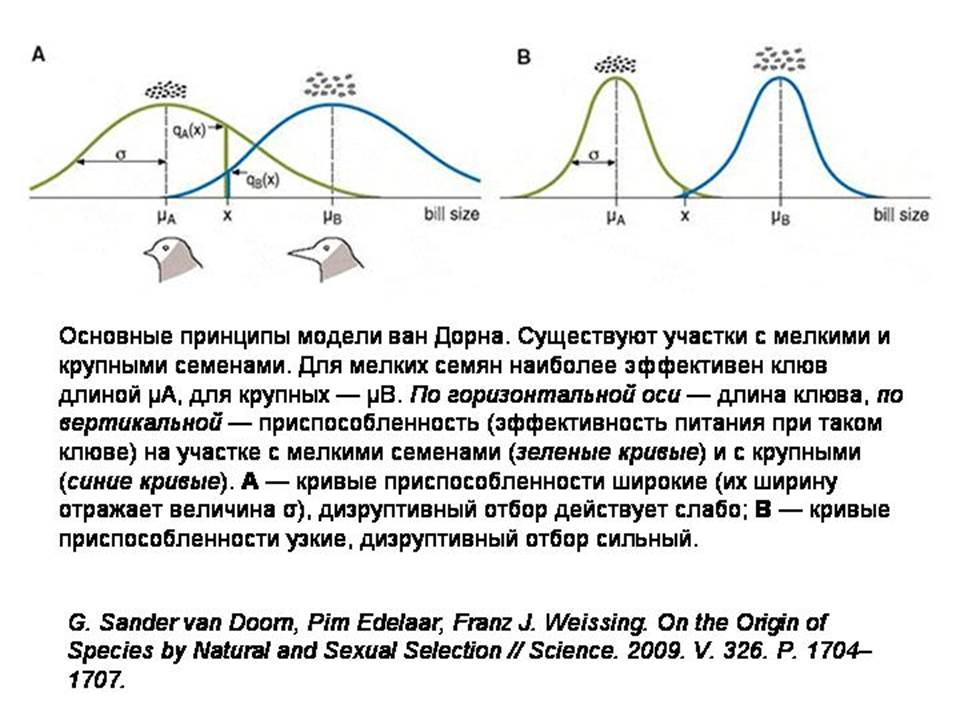
Модель ван Дорна показывает, что даже если все самки в популяции имеют одинаковые половые предпочтения, их привередливость может радикально повысить вероятность симпатрического видообразования.
Понятно, что половой отбор по индикаторам приспособленности повышает эффективность обычного отбора – как отрицательного (очищающего), так и положительного отбора, то есть фиксации полезных мутаций.
Допустим, существует популяция птиц с клювами средней длины. Местность, где они живут, состоит из участков двух типов. На одних участках преобладают растения с крупными семенами, на других — с мелкими. Для разгрызания крупных семян удобнее иметь длинный клюв, для мелких — короткий. В такой ситуации может начаться дизруптивный отбор. Это значит, что преимущество получат птицы с более длинными или более короткими клювами, но не с промежуточными. Известно, что у реальных птиц, попавших в подобные условия, может произойти симпатрическое видообразование. Остается только понять, каким образом видам удается разделиться, если их представители, перелетая с участка на участок, то и дело встречаются друг с другом и никто не мешает им образовывать смешанные пары и производить гибридное потомство с промежуточными по длине клювами.
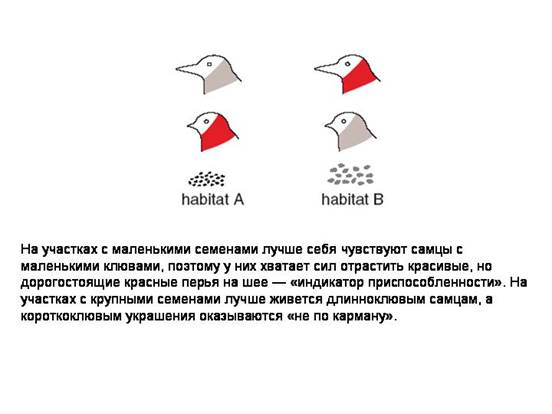
Главными параметрами модели являются интенсивность миграций молодых птиц и ширина кривой приспособленности (величина σ на рисунке). Чем больше сигма, тем легче разгрызать семена неподходящим по размеру клювом. Вместе эти две величины задают интенсивность дизруптивного отбора.
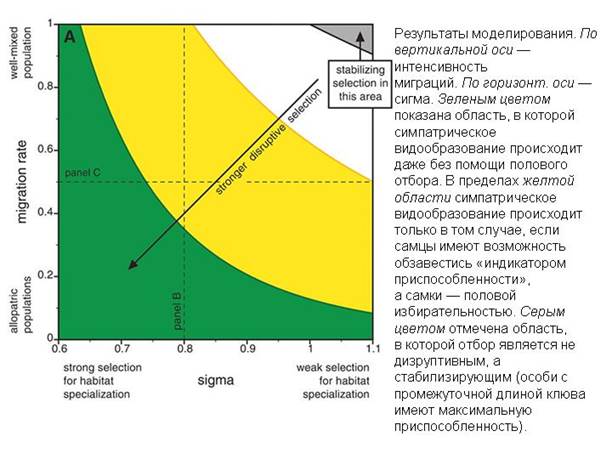
ПРИМЕР ПАРАЛЛЕЛЬНОГО СИМПАТРИЧЕСКОГО ВИДООБРАЗОВАНИЯ НА ОСТРОВАХ ТРИСТАН-ДА-КУНЬЯ
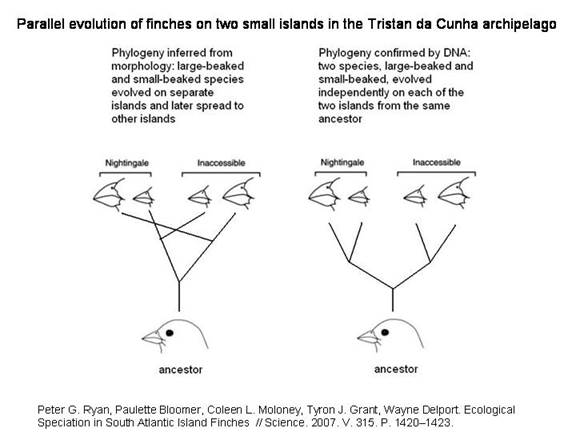
Считалось, что вьюрки рода Nesospiza, обитающие на двух маленьких островах архипелага Тристан-да-Кунья, относятся к двум видам: один с большим клювом, другой с маленьким. Генетический анализ показал, что эти птицы произошли от одной предковой формы, которая когда-то заселила оба острова, а затем на каждом из них подразделилась на большеклювую и мелкоклювую разновидности.


Что произошло? Ведь самок никто не отбирал, только самцов!
Именно Рональд Фишер первым понял, что не только брачные украшения самцов, но и вкусы и избирательность самок тоже наследуются и эволюционируют – по тем же правилам, что и остальные признаки (Дарвин предполагал, что это какое-то врожденное чувство прекрасного).
Эта простая мысль сделала теорию полового отбора полностью работоспособной. Однако идеи Фишера о половом отборе тоже не были восприняты научным сообществом. Они почти забылись к тому времени, когда их снова выкопали из забвения представители следующего поколения биологов-теоретиков, сумевшие, наконец, разработать убедительные и понятные математические модели. Лишь с третьей попытки теория полового отбора получила заслуженное признание.
Три важнейших идеи (модели), развитые в рамках теории полового отбора: фишеровское убегание, индикаторы приспособленности, принцип гандикапа.
Допустим, в популяции появился мутантный аллель, влияющий на брачные предпочтения самок. Допустим, самки с этим геном выбирают самых длинноухих самцов. В популяции существует небольшая нейтральная (не влияющая на приспособленность) изменчивость по длине ушей. Самки с мутантным геном поначалу не имеют никакого преимущества, но и особого вреда их странные вкусы им не приносят. Поэтому мутантный аллель имеет шанс за счет дрейфа достичь некоторой заметной частоты в генофонде популяции. Вот тут‑то и вступает в действие механизм “убегания”. Длинноухие самцы получают репродуктивное преимущество, потому что им доступны все самки, а короткоухие самцы могут спариться только с теми самками, у которых нет мутантного аллеля. Длинноухие самцы начинают оставлять больше потомков, чем короткоухие.
Потомство от браков длинноухих самцов с мутантными самками наследует не только “гены длинноухости” (от отца), но и аллель предпочтения длинноухих самцов (от матери). В результате эти аллели начинают распространяться.
Как только длинноухие самцы начинают оставлять в среднем больше потомства, чем короткоухие, самкам становится выгодно выбирать длинноухих партнеров, потому что тогда их сыновья унаследуют длинноухость, привлекут больше самок и оставят больше потомства. В результате те самки, которые предпочитают длинноухих самцов, начинают оставлять больше внуков, то есть получают репродуктивное преимущество.
Бывшая случайная прихоть превращается в полезную адаптацию. Возникает положительная обратная связь, в результате которой в генофонде быстро распространяются гены длинноухости и гены любви к длинноухим.
Длинноухость выгодна, потому что самки любят длинноухих, а любить длинноухих выгодно, потому что выгодна длинноухость. И никому нет дела до того, нужны ли длинные уши зачем‑то еще. В этом и состоит суть фишеровского убегания.
П.О. СПОСОБСТВУЕТ СИМПАТРИЧЕСКОМУ ВИДООБРАЗОВАНИЮ
Половой отбор может способствовать симпатрическое видообразование. Ясно, что без полового отбора и избирательного скрещивания симпатрическое видообразование едва ли возможно. До недавних пор все думали, что для симпатрического видообразования самки двух зарождающихся видов должны иметь разные половые предпочтения. И было не очень понятно, с чего бы вдруг у самок в одной популяции возникли разные вкусы. Но на самом деле, как было недавно показано, половой отбор стимулирует симпатрическое видообразование даже в том случае, если вкусы у всех самок остаются одинаковыми.
Модель ван Дорна показывает, что даже если все самки в популяции имеют одинаковые половые предпочтения, их привередливость может радикально повысить вероятность симпатрического видообразования.
Понятно, что половой отбор по индикаторам приспособленности повышает эффективность обычного отбора – как отрицательного (очищающего), так и положительного отбора, то есть фиксации полезных мутаций.
Допустим, существует популяция птиц с клювами средней длины. Местность, где они живут, состоит из участков двух типов. На одних участках преобладают растения с крупными семенами, на других — с мелкими. Для разгрызания крупных семян удобнее иметь длинный клюв, для мелких — короткий. В такой ситуации может начаться дизруптивный отбор. Это значит, что преимущество получат птицы с более длинными или более короткими клювами, но не с промежуточными. Известно, что у реальных птиц, попавших в подобные условия, может произойти симпатрическое видообразование. Остается только понять, каким образом видам удается разделиться, если их представители, перелетая с участка на участок, то и дело встречаются друг с другом и никто не мешает им образовывать смешанные пары и производить гибридное потомство с промежуточными по длине клювами.
Главными параметрами модели являются интенсивность миграций молодых птиц и ширина кривой приспособленности (величина σ на рисунке). Чем больше сигма, тем легче разгрызать семена неподходящим по размеру клювом. Вместе эти две величины задают интенсивность дизруптивного отбора.
ПРИМЕР ПАРАЛЛЕЛЬНОГО СИМПАТРИЧЕСКОГО ВИДООБРАЗОВАНИЯ НА ОСТРОВАХ ТРИСТАН-ДА-КУНЬЯ
Считалось, что вьюрки рода Nesospiza, обитающие на двух маленьких островах архипелага Тристан-да-Кунья, относятся к двум видам: один с большим клювом, другой с маленьким. Генетический анализ показал, что эти птицы произошли от одной предковой формы, которая когда-то заселила оба острова, а затем на каждом из них подразделилась на большеклювую и мелкоклювую разновидности.

Половой отбор — процесс, в основе которого лежит конкуренция за полового партнера между особями одного пола, что влечет за собой выборочное спаривание и производство потомства. Этот механизм может быть причиной эволюции некоторых характерных черт и приводить к их усилению. В пределах вида один из полов (практически всегда женский) играет роль ограниченного ресурса для другого (практически всегда мужского).
Некоторыми современными этологами высказывается мнение, что чувство любви и самопожертвования является следствием полового отбора и внутриродственной селекции на генетическом уровне [1] .
См. также
Примечания
Ссылки
Литература
- Механизмы эволюции
- Факторы эволюции
- Половой отбор
Wikimedia Foundation . 2010 .
Полезное
Смотреть что такое "Половой отбор" в других словарях:
Половой отбор — Отбор специфических характеристик, способствующих успеху животного при спаривании в брачный период. Существует два вида полового отбора: 1. Развитие характеристик одного пола (обычно самцов), позволяющее животным соперничать друг с другом за… … Большая психологическая энциклопедия
Половой отбор — * палавы адбор * sexual selection теория, развитая Ч. Дарвином, согласно которой внутри вида (см.) между самцами идет борьба за скрещивание, определяющаяся признаками, способствующими успеху в борьбе за существование вида в целом. В литературе П … Генетика. Энциклопедический словарь
ПОЛОВОЙ ОТБОР — ПОЛОВОЙ ОТБОР, процесс жизнедеятельности животных, основанный на успешном поиске пары и производстве потомства. Половой отбор сходен с ЕСТЕСТВЕННЫМ ОТБОРОМ, однако в отличие от него здесь большое значение имеют ВТОРИЧНЫЕ ПОЛОВЫЕ ПРИЗНАКИ.… … Научно-технический энциклопедический словарь
ПОЛОВОЙ ОТБОР — форма естественного отбора у животных, основанного на соперничестве особей одного пола (преимущественно мужского) за обладание особями другого пола, ведущая к уменьшению потомства у менее приспособленных. В результате полового отбора у многих… … Большой Энциклопедический словарь
ПОЛОВОЙ ОТБОР — форма естеств. отбора у ряда групп животных, основанная на соперничестве особей одного пола (чаще мужского) за спаривание с особями др. пола. В результате П. о. или при его участии у мн. видов животных в процессе эволюции возникли и развились… … Биологический энциклопедический словарь
Половой отбор — особая форма естественного отбора (См. Естественный отбор), определяющая возникновение в процессе эволюции вторичных половых признаков (См. Вторичные половые признаки). К таким признакам относятся: яркая брачная окраска оперения уток,… … Большая советская энциклопедия
Половой отбор — Тип естественного отбора, который осуществляется только в пределах одного пола внутри вида. Является результатом борьбы за партнера для спаривания и приводит к половому диморфизму с приобретением одной или более черт. отбор, происходящий в силу… … Физическая Антропология. Иллюстрированный толковый словарь.
половой отбор — форма естественного отбора у животных, основанная на соперничестве особей одного пола (преимущественного мужского) за обладание особями другого пола, ведущая к уменьшению потомства у менее приспособленных. В результате полового отбора у многих… … Энциклопедический словарь
половой отбор — форма естественного отбора, основанного на соперничестве особей одного пола (гл. обр. муж.) за обладание особями др. пола, ведущая к уменьшению потомства у менее приспособленных. (Источник: Словарь сексуальных терминов) … Сексологическая энциклопедия
ПОЛОВОЙ ОТБОР — форма естеств. отбора у ж ных, основанная на соперничестве особей одного пола (преим. муж.) за обладание особями др. пола, ведущая к уменьшению потомства у менее приспособленных. В результате П. о. у мн. видов ж ных возникли вторичные половые… … Естествознание. Энциклопедический словарь
Читайте также: