
На каких уровнях возникают новые комбинации генов кратко
Обновлено: 05.07.2024
Любовь Петровна Малиновская — аспирант Новосибирского государственного университета, научный сотрудник лаборатории молекулярной цитогенетики птиц Института цитологии и генетики СО РАН. Область научных интересов — молекулярная цитогенетика птиц.
Анна Александровна Торгашёва — кандидат биологических наук, старший научный сотрудник, заведующая лабораторией молекулярной цитогенетики птиц Института цитологии и генетики СО РАН. Научные интересы связаны с молекулярной биологией мейоза, эволюционной цитогенетикой животных.
Виген Артаваздович Геодакян на съезде Вавиловского общества генетиков и селекционеров, проходившем в Московском государственном университете имени М. В. Ломоносова в июне 2009 г. Фото А. А. Торгашёвой
В этой статье мы хотим рассказать историю, которая, на первый взгляд, противоречит идеям Геодакяна. И на второй — тоже. Но как жаль, что его больше нет с нами. Почему-то нам кажется, что он и в ней нашел бы подтверждение своей теории. С теплыми воспоминаниями и светлой грустью мы посвящаем эту статью его доброй памяти.
Неправильное правило Хаксли — Холдейна
Гетерогаметный — значит, имеющий разные половые хромосомы: XY у самцов млекопитающих и двукрылых (включая комаров и муху дрозофилу) или ZW у самок птиц и бабочек. Соответственно, гомогаметным полом оказываются самки ХХ у первых и самцы ZZ у последних. т.е. правило Хаксли — Холдейна говорило не о самцах и самках, а о гетеро- и гомогаметном полах. В чем здесь фокус? Почему нужно блокировать рекомбинацию у гетерогаметного пола?
Рекомбинация наряду с мутациями служит мощным генератором генетической изменчивости. Но между ними есть большая разница. Мутационный процесс — это своего рода проявление энтропии на молекулярно-генетическом уровне. За счет неизбежных ошибок удвоения и ошибочной репарации (исправления) этих ошибок в ДНК возникают мутации — искажения генетической информации, упорядоченной предшествующим отбором, т.е. мутации — это неизбежное нарастание хаоса.
Рекомбинация — это хорошо упорядоченная, запрограммированная и затратная генерация хаоса (рис. 1). В ходе рекомбинации в ДНК запрограммированно вносится множество вполне фатальных повреждений — двунитевых разрывов. Эти разрывы затем репарируются: иногда правильно — с восстановлением исходной информации, но чаще неправильно — не по той цепи, которой нужно, а то и вовсе крест-накрест. Эти перекресты приводят к тому, что родительские комбинации аллелей, которые, между прочим, позволили родителям дожить до брака и приступить к производству потомков, разрушаются. Вместо них возникают новые, ранее не опробованные комбинации, которые вполне могут оказаться как хуже, так и лучше родительских [4].
Рис. 1. Рекомбинация гомологичных хромосом [6]. Розовым цветом обозначена ДНК материнской хромосомы, синим — отцовской, зеленым — ДНК, достроенная в ходе репарации разрывов
Таким образом, от правила Хаксли — Холдейна осталось только то, что самцы отличаются от самок. Да и то оказалось, что половые различия в уровне рекомбинации сильно варьируют и в размахе (когда сильные, когда слабые, когда и вовсе отсутствуют), и в знаке (чаще всего больше у самок, но бывает и у самцов). И если эти различия есть, то они никак не связаны с гетерогаментостью. И все же, почему рекомбинация, как правило, выше у самок?
Половой отбор как универсальное объяснение всего на свете
Половой отбор — это красивая и остроумная теория. Но при чем здесь рекомбинация? Как пониженная рекомбинация у самцов может им помочь в борьбе за самок? Как может самка определить рекомбинационные способности самца и зачем ей это нужно? Уровень рекомбинации — это такой признак самца, который можно оценить, только скрестившись с ним и посмотрев на разнообразие полученных потомков. Но если потомки уже получены, значит выбор уже сделан и оценка рекомбинации уже никому не нужна. Но даже если бы самка могла определить способности самца заранее, почему ей должны нравиться мало рекомбинирующие самцы?
Рис. 2. Половой отбор убегающего признака и рекомбинация. А — аллель, контролирующий у самок предпочтение самцов с длинным хвостом; а — нет предпочтений у самок; Т — аллель, контролирующий у самцов развитие длинного хвоста; t — нормальный хвост у самцов
От моделей к фактам
Теорий, которые объясняют половые различия в рекомбинации (гетерохиазмию), более чем достаточно. С фактами ситуация гораздо хуже. Данных мало, и они довольно противоречивы. Теоретики по определению не занимаются экспериментами, а используют в своих моделях данные, полученные другими людьми и для других целей. В итоге довольно часто им приходится сравнивать, фигурально выражаясь, ужа и ежа просто потому, что других данных нет.
С птицами, которые интересны тем, что у них гетерогаметный пол — самки, ситуация не лучше. Данные по рекомбинации у обоих полов получены для девяти видов, которых тоже разделяют десятки миллионов лет независимой эволюции и отличают друг от друга множество особенностей их биологии. Половые различия по рекомбинации у птиц непредсказуемо варьируют: у близких видов они могут сильно различаться по знаку и размаху [10]. Поэтому мы решили получить собственные данные по рекомбинации для двух видов птиц, которые разошлись относительно недавно в ходе эволюции и сходны по всем особенностям биологии, кроме одной: у одного вида нет полового отбора, а у другого он доказан наблюдениями и экспериментами.
Две ласточки
Мы сравнили два вида ласточек — деревенскую ласточку (Hirundo rustica) и бледную береговушку (Riparia diluta) по параметрам рекомбинации у самцов и самок [11]. Современные цитологические методы позволяют выявлять точки рекомбинации прямо на препаратах хромосом (рис. 3).
Рис. 3. Микрофотография сперматоцита деревенской ласточки после иммуноокрашивания белков осей хромосом (красный), центромер (синий) и рекомбинационных узелков (зеленый) [10]. Цифрами обозначены макрохромосомы, буквами — половые хромосомы. Фото Л. П. Малиновской
Почему мы выбрали именно эти виды? Потому что они состоят в относительно близком родстве (около 16 млн лет дивергенции) и довольно похожи в отношении экологии, образа жизни и систем размножения. У них сходны районы миграции и гнездования. Оба вида образуют большие колонии. Оба практикуют относительно строгую моногамию, совместное высиживание яиц и выкармливание потомков.
Различия между ними состоят в том, что у береговушек практически невозможно отличить самцов от самок, в то время как самцы европейского подвида деревенской ласточки (ареал которого доходит до Байкала) отличаются от самок большей яркостью оперения и, главное, большей длиной хвостовых направляющих перьев (косиц). По этому признаку у деревенской ласточки идет довольно интенсивный половой отбор. Самки отдают безусловное предпочтение самцам с более длинными косицами. Такие самцы оставляют больше потомков. Они раньше других находят себе пару, чаще других вступают во внебрачные отношения с соседними самками, а их собственные самки чаще производят две кладки в сезон. Эти различия в репродуктивном успехе были подтверждены как прямыми наблюдениями, так и экспериментами с искусственным удлинением или укорочением косиц [11].
Брачные пары деревенской ласточки (слева) и бледной береговушки. Фото Е. Шнайдер
Рис. 4. Половые различия по числу (вверху) и распределению (внизу) рекомбинационных узелков на хромосомах деревенской ласточки и бледной береговушки [10]
Рекомбинация на концах хромосом важна и полезна для правильного их расхождения в мейозе, но мало что дает для перетасовки аллелей. В то время как рекомбинация в средних частях хромосом дает широкий и разнообразный набор новых аллельных комбинаций в потомстве. За счет этих двух особенностей — более высокой частоты рекомбинации вообще и в средних частях хромосом в частности — самки деревенской ласточки разрушают старые комбинации аллелей и создают новые гораздо быстрее, чем самцы того же вида. При этом у бледной береговушки — морфологически мономорфного вида — мы не обнаружили значительных половых различий ни по числу рекомбинационных узелков, ни по их распределению. И, что особенно интересно, они очень похожи по этим признакам на самцов деревенской ласточки.
Логично допустить, что у общего предка наших двух видов ласточек самцы и самки были похожи друг на друга и по морфологии, и по уровню рекомбинации. Затем половой отбор у деревенских ласточек привел к возникновению полового диморфизма по обоим этим признакам. Но почему это произошло не за счет снижения рекомбинации у самцов, а за счет ее повышения у самок? Можно думать, исходно низкий уровень рекомбинации у обоих полов позволил возникнуть и закрепиться неравновесию по сцеплению между аллелями, контролирующими проявление вторичных половых признаков у самцов, и предпочтение к этим признакам у самок. Поскольку половая разница в рекомбинации у деревенской ласточки наиболее выражена в макрохромосомах, можно предположить, что именно там находятся гены, законсервированные половым отбором. Сильный отбор по тем районам хромосом, где они находятся, должен вести к накоплению вредных мутаций в этих и соседних районах. Увеличение частоты рекомбинации в макрохромосомах в женском мейозе может быть направлено на очищение от этих мутаций. И кроме того, вспомните, что самки любят самцов не просто с длинным хвостом, но с самым длинным. Отсюда следует, что самки должны интенсивно тасовать свои аллели, надеясь снабдить своих сыновей более длинными хвостами. Так что наши данные не так уж и противоречат идеям Геодакяна. Да, сами самки, по его теории, должны быть более мономорфными, чем самцы. Но при этом они могут генерировать более разнообразное потомство, чтобы их дочерям в следующем поколении было, из кого выбирать.
Конечно, эта гипотеза весьма умозрительна. Нам нужно больше данных о величине и направлении гетерохиазмии среди близких видов птиц с явным половым отбором и без него. Нам нужны данные о локализации генов, контролирующих вторичные половые признаки самцов и особенности половых предпочтений у самок. И нам очень не хватает Вигена Артаваздовича, чтобы обсудить с ним эти данные. Он бы обязательно придумал им правильное объяснение.
Работа выполнена при поддержке Российского научного фонда (проект 20-64-46021).
Литература
1. Геодакян В. А. Эволюционная роль половых хромосом (новая концепция) // Генетика. 1998; 34(8): 1171–1184.
2. Haldane J. B. S. Sex-ratio and unisexual sterility in hybrid animals // J. Genet. 1922; 12: 101–109. DOI: 10.1007/BF02983075.
3. Huxley J. S. Sexual difference of linkage in Gammarus chevreuxi // J. Genet. 1928; 20: 145–156. DOI: 10.1007/BF02983136.
4. Bishop D. K., Zickler D. Early decision; meiotic crossover interference prior to stable strand exchange and synapsis // Cell. 2004; 117(1): 9–15. DOI: 10.1016/s0092-8674(04)00297-1.
5. Sardell J. M., Kirkpatrick M. Sex differences in the recombination landscape // Am. Nat. 2020; 195: 361–379. DOI: 10.1086/704943.
6. Миллер Дж. Соблазняющий разум: Как выбор сексуального партнера повлиял на эволюцию человеческой природы. М., 2020.
7. Fisher R. A. The Genetical Theory of Natural Selection. Oxford, 1930.
8. Trivers R. Sex Differences in Rates of Recombination and Sexual Selection // The Evolution of Sex: an Examination of Current Ideas / Michod R. E. and B. R. Levin (eds). Sunderland, 1988; 270–286.
9. Mank J. E. The evolution of heterochiasmy: The role of sexual selection and sperm competition in determining sex-specific recombination rates in eutherian mammals // Genet. Res. 2009; 91: 355–363. DOI: 10.1017/S0016672309990255.
10. Malinovskaya L. P., Tishakova K., Shnaider E. P., Borodin P. M., Torgasheva A. A. Heterochiasmy and sexual dimorphism: The case of the barn swallow (Hirundo rustica, Hirundinidae, aves) // Genes (Basel). 2020; 11(10): 1119. DOI: 10.3390/genes11101119.
11. Møller A. P. Sexual Selection and the Barn Swallow. Oxford, 1994.
** См. например: Бородин П. М. Генетическая рекомбинация в свете эволюции // Природа. 2007. № 1. С. 14–22.
Изменчивость — это возникновение индивидуальных различий. На основе изменчивости организмов появляется генетическое разнообразие форм, которые в результате действия естественного отбора преобразуются в новые подвиды и виды. Различают изменчивость модификационную, или фенотипическую, и мутационную, или генотипическую.
Вопрос 2. Приведите классификацию мутаций по уровню изменений наследственного материала.
По уровню изменений наследственного материала выделяют следующие виды мутаций:
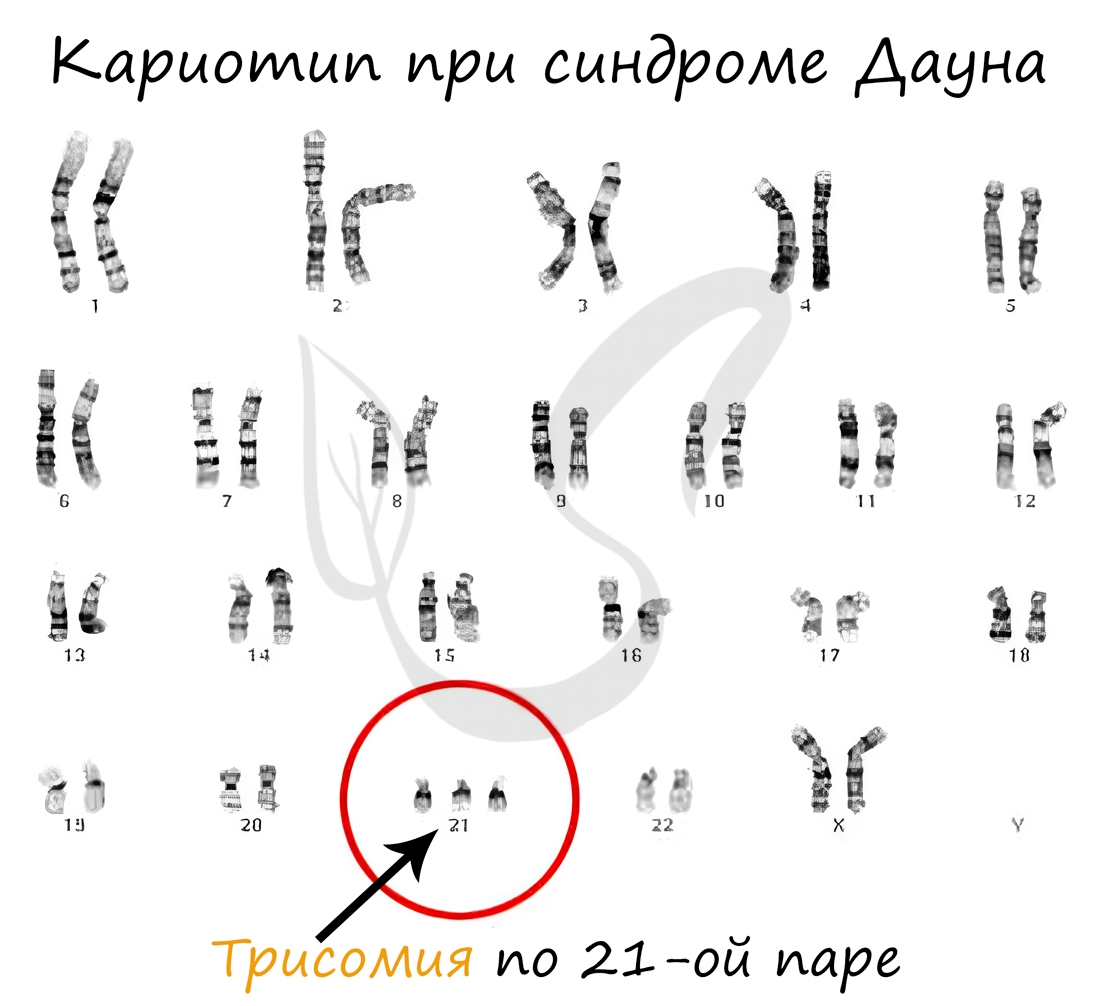
1. Геномные мутации - изменения кариотипа, кратные (3п, 4п, 8п. ) и некратные (2п ± 1; 2п 2. ) гаплоидному числу хромосом. Например, при болезни Дауна в кариотипе присутствуют три хромосомы 21-й пары.
2. Генные, или точковые, мутации - изменения, обусловленные заменой, выпадением или вставкой одного или нескольких нуклеотидов в пределах одного гена. Они влекут за собой изменение структуры белков, заключающееся в появлении новой последовательности аминокислот в полипептидной цепи.
3. Хромосомные мутации - изменение Структуры хромосом. Эти мутации могут возникать вследствие утраты хромосомой своей части или приобретение нового, нехарактерного для нее участка, что может принести к гибели организма.
Вопрос 3. Что такое полиплоидия и каково её значение?
Полиплоидия - увеличение числа хромосом, кратное гаплоидному набору. Часто подобное явление встречается в простейших и растений. Полиплоидия позволяет повысить надежность генетической системы, уменьшает опасность снижения жизнеспособности в случае возникновения мутаций, повышает жизнеспособность, плодовитость и другие свойства. В растениеводстве этим пользуются, искусственно получая полиплоидные сорта культурных растений, которые отличаются высокой урожайностью и жизнестойкостью.
Вопрос 4. Перечислите свойства мутаций.
Мутации передаются по наследству, чем обусловлена их роль в эволюции: только наследственные изменения могут стать достоянием последующих поколений при условии успешного размножения и выживания особей с этими мутациями.
Мутации вызываются различными внешними и внутренними факторами. Ультрафиолетовые лучи, колебания температуры, изменение химических реакций в клетке в связи с ее старением, действие различных химических веществ могут привести к изменениям структуры ДНК и целых хромосом.
Возникают мутации внезапно, скачкообразно, у отдельных особей вида и в большинстве случаев вредны для организма, так как расшатывают исторически сложившийся генотип. Одни и те же мутации могут возникать повторно.
Мутации ненаправленны: мутировать может любой ген, вызывая изменения как незначительных, так и жизненно важных признаков. При этом один и тот же фактор, например рентгеновское излучение, действуя на клетки, может вызвать самые разные мутации, которые трудно предвидеть.
Вопрос 5. На каких уровнях возникают новые комбинации генов?
Новые комбинации генов возникают на различных уровнях:
1. перекомбинация генов в группе сцепления вследствие кроссинговера в профазе I мейотического деления;
2. расхождение гомологичных хромосом из бивалентов в I мейотическом делении;
3. расхождение дочерних хромосом во II мейотическом делении. Всего во время мейоза образуется 2^46 комбинаций.;
4. слияние половых клеток, принадлежащих разным организмам (292 комбинаций).
ВОПРОСЫ И ЗАДАНИЯ ДЛЯ ОБСУЖДЕНИЯ
Вопрос 1. Благодаря чему формируется резерв наследственной изменчивости? Каково его значение?
Вопрос 2. С помощью каких воздействий можно повысить частоту мутаций?
Чтобы повысить частоту мутаций необходимо воздействовать на клетки различными мутагенными факторами, такими как:
1. Ультрафиолетовое излучение;
2. Химическими соединениями не встречающимися в природе (пестициды, некоторые лекарственные препараты и др.)
3. Органическими и неорганическими соединениями естественного происхождения (окислы азота, нитраты, радиоактивные соединения, алкалоиды).
Вопрос 3. Приведите примеры комбинативной изменчивости, вытекающие из закономерностей наследования признаков, выявленных Г. Менделем.
В основе комбинативной изменчивости лежит половое размножение организмов, вследствие которого возникает огромное разнообразие генотипов. Практически неограниченными источниками генетической изменчивости служат три процесса: независимое гомологичных хромосом, Взаимный обмен участками гомологичных хромосом, или кроссинговер, Случайное сочетание гамет при оплодотворении.
У цветка ночная красавица есть ген красного цвета лепестков А, и ген белого цвета а. Организм Аа имеет розовый цвет лепестков. Таким образом, у ночной красавицы нет гена розового цвета, розовый цвет возникает при сочетании (комбинации) красного и белого гена.
2. Исторически сложилось так, что наследственная изменчивость имеет несколько названий. Перечислите их.
1. генотипическая
2. неопределенная
3. индивидуальная.
3. Укажите два источника (механизма) наследственной изменчивости.
1. мутации
2. комбинации хромосом при половом размножении и кроссинговер.
4. Повторив материал § 31 главы 12, ответьте, какой внутриклеточный процесс обеспечивает появление новых комбинаций наследственных задатков (генов), содержащихся в гомологичных хромосомах.
Перегруппировка хромосом при половом размножении, конъюгация и кроссинговер.
5. Основываясь на знаниях, полученных при изучении § 31 главы 12, вставьте в предложение недостающие слова.
Перегруппировка хромосом, приводящая к появлению в гаметах новых комбинаций негомологичных хромосом, происходит в результате кроссинговера в профазе 1 мейотического деления.
6. Укажите, как классифицируются мутации.
По уровню возникновения: генные, хромосомные и геномные.
По месту возникновения: генеративные и соматические.
По характеру появления: доминантные и рецессивные.
7. Приведите определение мутационной изменчивости. Перечислите основные свойства мутаций.
Мутационная изменчивость – это изменчивость, при которой происходят наследственные изменения генетического материала.
Свойства мутаций:
1. наследуются
2. вызываются различными внешними и внутренними факторами
3. возникают внезапно; ненаправленны (мутировать может любой ген).
8. Выберите из приведенных в скобках и впишите правильные варианты пропущенных слов и словосочетаний.
Большое эволюционное значение имеют рецессивные мутации, так как сохраняются у гетерозиготных организмов и составляют необходимый резерв наследственной изменчивости для эффективного протекания естественного отбора. (Доминантные, рецессивные, генофонд вида, гомозиготные, гетерозиготные, резерв наследственной изменчивости, генетический груз, гомогаметные, гетерогаментные, гомологичные, аналогичные, аллельные, неаллельные.)
9. Объясните, что такое полиплоидия, каковы ее фенотипические последствия.
Полиплоидия – изменение хромосомного набора, при котором происходит увеличение числа хромосом, кратное гаплоидному набору.
Фенотипические последствия полиплоидии: увеличение размеров организма (а также плодов, семян у растений), повышение плодовитости и продуктивности, повышение жизнеспособности.
10. Дайте определение хромосомных мутаций. Приведите примеры.
Хромосомные мутации – изменения структуры хромосом.
Примеры: утрата жизненно важного участка хромосомы и гибель организма, потеря части 21-й хромосомы – острый лейкоз у новорожденных и др.
11. Объясните, что такое генные мутации, и к каким последствиям на молекулярном уровне они приводят.
Генные мутации – изменения, обусловленные заменой одного или нескольких нуклеотидов в пределах одного гена.
Последствия генных мутаций: они влекут за собой изменение строения белков, заключающееся в появлении новой последовательности аминокислот в полипептидной цепи и, как следствие, изменение функциональной активности белковой молекулы.
Под изменчивостью понимают способность организмов приобретать признаки и свойства, отличные от родительских, характерных для данного вида. Изменчивость является общим свойством всех живых систем и может выражаться в изменении как генотипа, так и фенотипа.
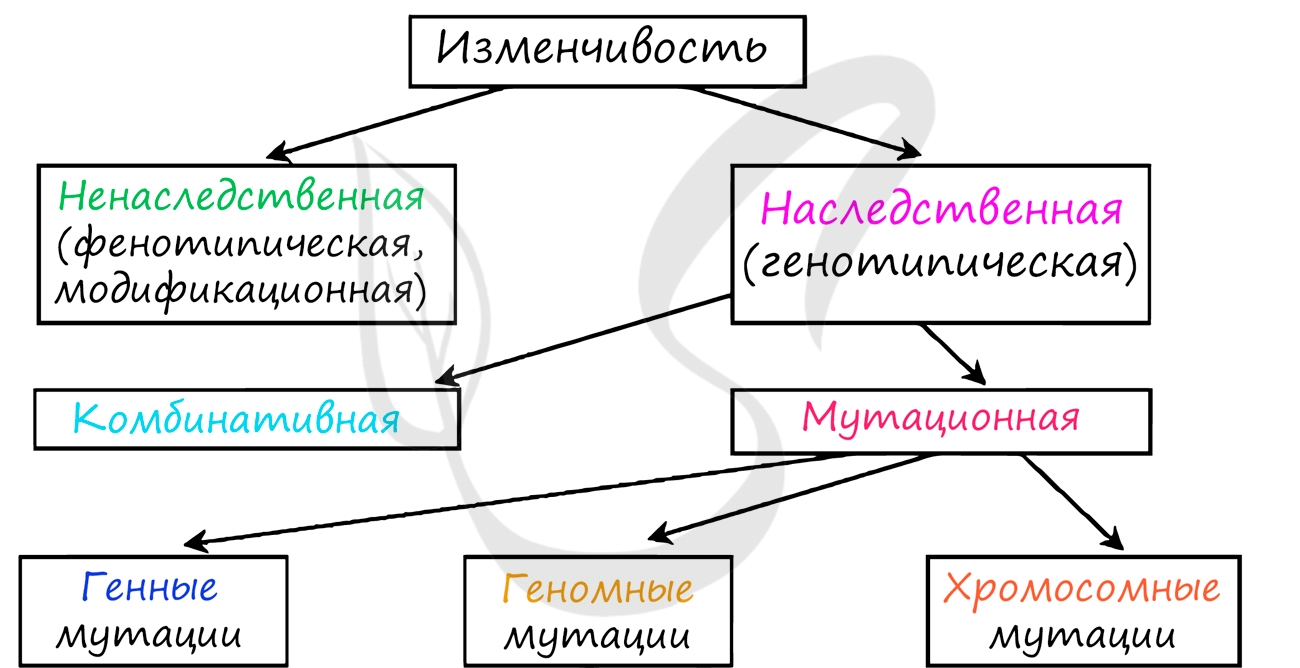
Традиционно различают ненаследственную и наследственную изменчивость.

Модификационная изменчивость
Модификационная (фенотипическая) изменчивость - изменения фенотипа организма, обусловленные влиянием факторов внешней среды. Данный вид изменчивости не приводит к изменениям генотипа особи - все изменения касаются только фенотипа.
Напомню, что генотипом называют генетическую конституцию - совокупность генов одного организма, полученных от родителей. Фенотип (греч. phаino - обнаруживаю) - совокупность наблюдаемых характеристик организма (любой морфологический, гистологический, биохимический, поведенческий признак).
Для модификационной изменчивости характерен групповой характер, она часто (но не всегда) служит приспособлением к условиям внешней среды. Известным примером модификационной изменчивости является изменение окраски шерсти у зайца-беляка в зависимости от сезона года.

Такое изменение окраски делает их более приспособленными, повышает выживаемость: заяц сливается с внешней средой и становится незаметен для хищников.
Однако не стоит забывать об относительности любой приспособленности: если среда резко изменится, то белый заяц на фоне темной земли станет легкой добычей для хищников.

Еще одним примером модификационной изменчивости служит изменение окраски шерсти у гималайских кроликов. Они рождаются полностью белыми, так как их эмбриональное развитие протекает в условиях повышенной температуры.
Однако в результате воздействия холода на разные участки их тела, шерсть начинает темнеть. В естественных условиях шерсть темная на ушах, носе, лапах и хвосте.
В эксперименте лед привязывают к спине, и через некоторое время шерсть на этом месте начинает темнеть. Это наглядно демонстрирует влияние внешней среды на проявление признака.

Вам известно, что человек, побывавший на солнце, получает его "отпечаток" - загар. Потемнение цвета кожи в данном случае связано с активной выработкой пигмента меланина, который защищает кожу и внутренние органы от УФ излучения.
Загар также является типичным примером модификационной изменчивости. Одни люди загорают быстро, у других этот процесс занимает гораздо больше времени - все дело в норме реакции.
Норма реакции
Нормой реакции называют генетически (наследственно) закрепленные пределы (границы) изменчивости признака. Принято говорить, что у каждого признака существует определенная норма реакции: она может быть узкой или широкой.
Узкая норма реакции характерна для признаков, которые относятся к качественным: форма глаза, желудка, сердца, размеры головного мозга, рост.
Количественные признаки имеют широкую норму реакцию и достаточно вариабельны в течение жизни: яйценоскость кур, удойность коров, вес, размер листьев.
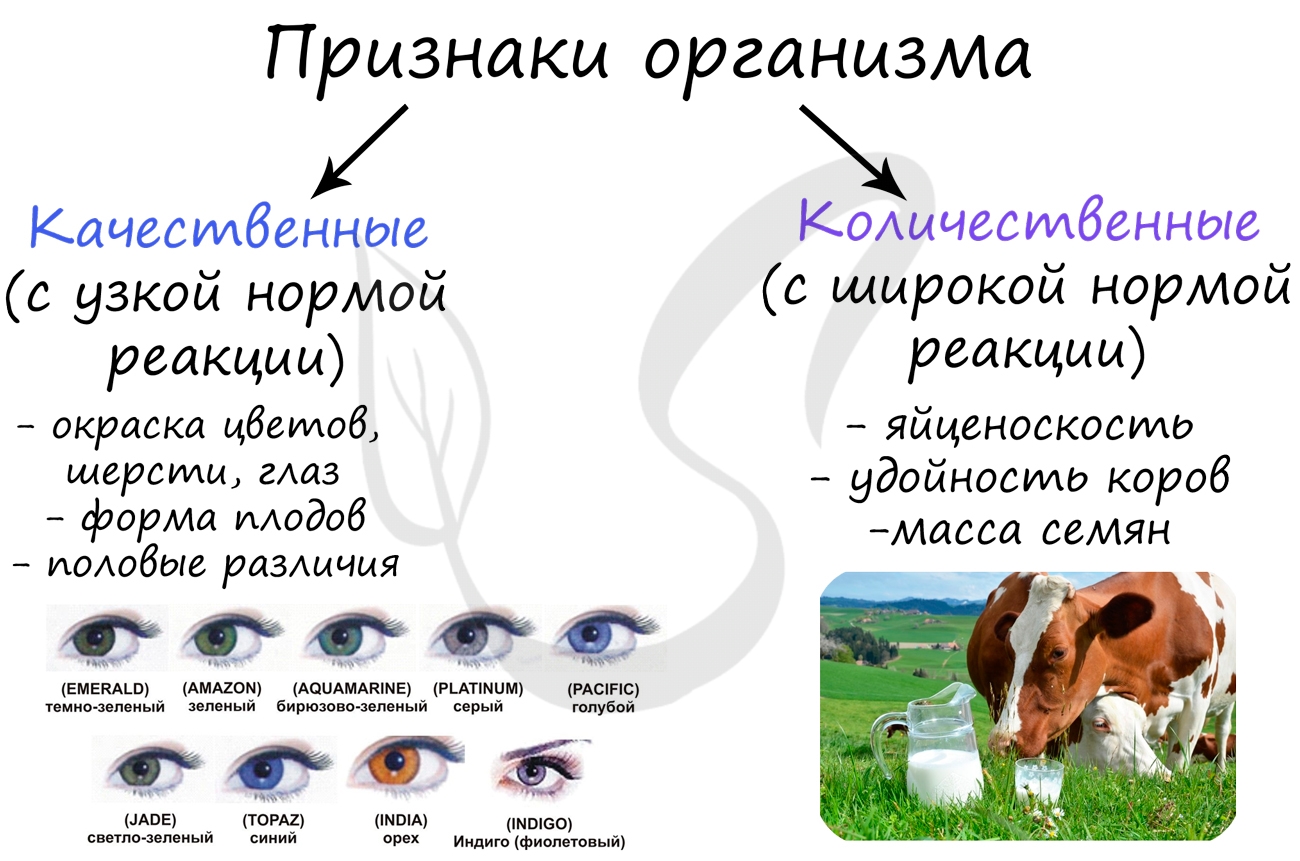
- Причина изменения - влияние факторов внешней среды
- Изменения признаков организма не затрагивают генотип, происходят в соматических клетках и не передаются потомкам
- Изменение признаков ограничено в пределах нормы реакции, которая определяется генотипом
- Изменчивость носит групповой характер, характерна для многих особей (к примеру, сезонная изменчивость)
Наследственная изменчивость
Наследственная изменчивость (неопределенная, индивидуальная, генотипическая) - форма изменчивости, вызванная изменениями генотипа организма, которые могут быть связаны с мутационной или комбинативной изменчивостью.
В отличие от модификационной изменчивости, где затрагивается только фенотип (внешние проявления), генотипическая изменчивость затрагивает генотип, а это означает, что генетические изменения затрагивают и половые клетки, которые передаются потомству. Поэтому и называется она - наследственная.
Комбинативная изменчивость
Комбинативная изменчивость возникает в результате появления у потомков новых сочетаний генов (комбинаций). Эти комбинации возникают во время мейоза в результате хорошо вам знакомого (я надеюсь!) кроссинговера - обмена участками между гомологичными хромосомами.
- Случайная комбинация генов в ходе кроссинговера
- Независимое расхождение хромосом в мейозе
- Случайная встреча гамет при оплодотворении
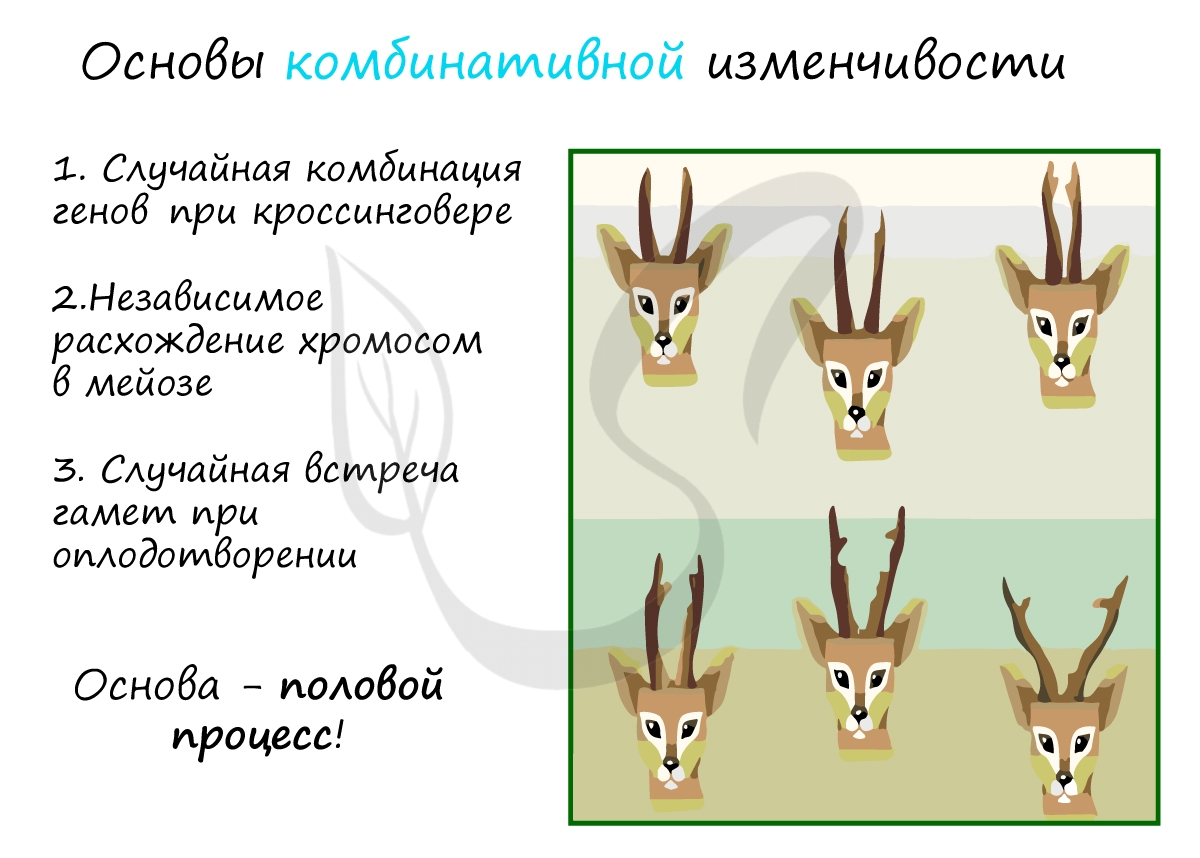
Я всегда говорю ученикам, что комбинативная изменчивость - это полная неопределенность: мы не знаем, какие комбинации возникнут между генами при кроссинговере, не знаем, какие хромосомы образуются и в какие гаметы они разойдутся, и, наконец, не знаем какие половые клетки (гаметы) встретятся при оплодотворении.
То, что мы отличаемся от своих родителей, и есть результат этих неопределенностей.

Мутационная изменчивость
Мутационная изменчивость связана с возникновением мутаций. Мутации (лат. mutatio - изменение) - внезапные, возникающие спонтанно или вызванные мутагенами наследуемые изменения генетического материала, приводящие к изменению тех или иных признаков организма.
- Мутации - резкие спонтанные изменения генотипа
- Стойкие, передаются потомкам через половые клетки (гаметы)
- Ненаправленные. Большинство мутаций - вредные (часть из них летальные), лишь очень небольшая часть носит полезный приспособительный характер, мутации также могут быть безразличными (нейтральными) для организма
- Носят индивидуальный характер

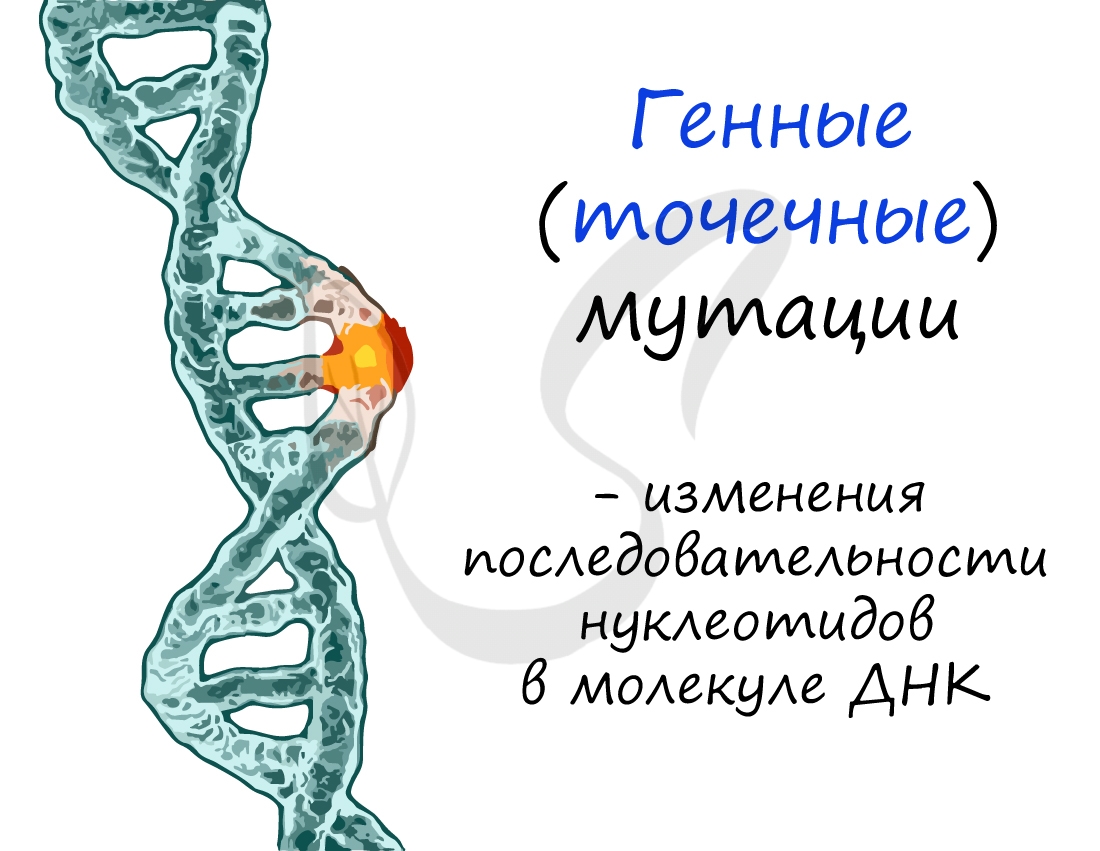
Изменения при генных мутациях происходят в последовательности нуклеотидов молекулы ДНК. Может случаться такое, что один или несколько нуклеотидов выпадают из ДНК (делеция), вставляются новые нуклеотиды, удваиваются имеющиеся нуклеотиды (дупликация).
Изменения ДНК ведут к тому, что в результате на рибосомах синтезируется белок с иной аминокислотной последовательностью. К примеру: изначально триплет ДНК "ТАЦ" кодировал аминокислоту "Мет", нуклеотид "Т" выпал из триплета произошла вставка нуклеотида "Г". В результате вместо аминокислоты "Мет" теперь синтезируется аминокислота Вал.
Новые аминокислоты могут поменять свойства белка, так что признак, за который он отвечает, будет меняться. Только что вы узнали об универсальной схеме - изменении фенотипа в результате изменений генотипа.
В результате хромосомных мутаций происходят структурные изменения хромосом (не следует путать с кроссинговером, который происходит в норме и подразумевает обмен участками между гомологичными хромосомами). Последствия хромосомных мутаций часто оказываются летальны.
В результате таких мутаций может происходить утрата (делеция) участка хромосомы, его удвоение (дупликация), поворот на 180° (инверсия), перенос участка одной хромосомы на другую (транслокация), перенос участка внутри одной хромосомы (транспозиция).

-
Автополиплоидию - кратное увеличение числа наборов хромосом
В результате таких мутаций количество хромосом увеличивается в кратное количество раз (2,3,4 и т.д.). В результате получаются организмы триплоиды, тетраплоиды и т.д. Иногда такие мутации вызывают искусственно, к примеру, в селекции растений. Известно, что у полиплоидов более крупные и сочные плоды.
В селекции полиплоидию у растений вызывают добавлением специального химического вещества - колхицина, который блокирует образование нитей веретена деления. Вследствие этого хромосомы не расходятся и остаются в одной клетке - набор хромосом увеличивается в 2 раза.

Имеет значение в процессе видообразования. Примером данной мутации может послужить отдаленная гибридизация (аутбридинг) пшеницы и ржи. Их генотип состоит из гаплоидного набора пшеницы (n) и гаплоидного набора ржи (m).
В результате такого скрещивания в 1875 году в Шотландии был получен первый искусственный стерильный гибрид - тритикале. Тритикале дает отличный урожай, в дальнейшем путем полиплоидии стерильность данного гибрида была преодолена.
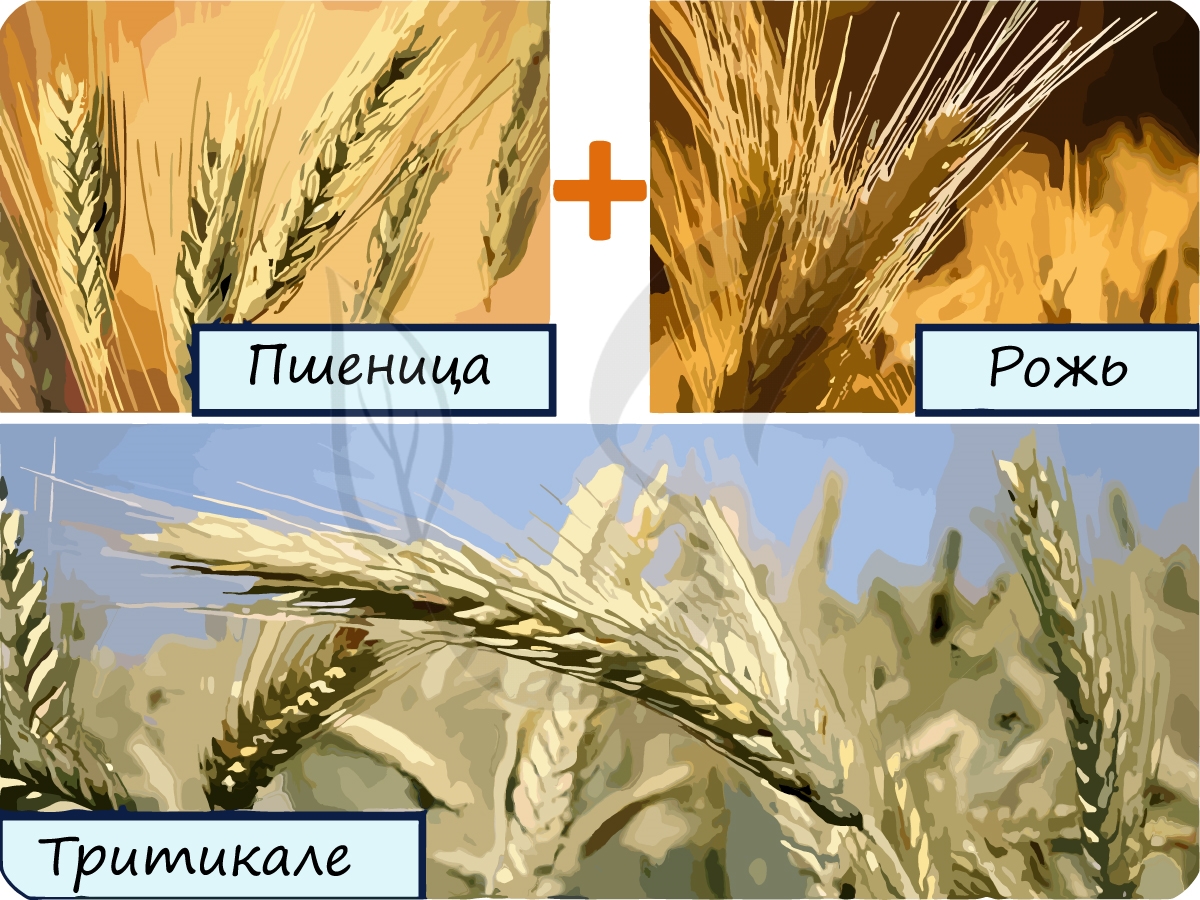
Также примером отдаленной гибридизации, соответственно и аллополиплоидии, является гибрид осла (самца) и лошади (самки) - мул. Это животное отличается большой выносливостью, но опять-таки бесплодное вследствие геномной мутации.

Анеуплоидия - изменение кариотипа (совокупность признаков хромосом), при котором число хромосом в клетках не кратно гаплоидному набору (n). Таким образом, в результате анеуплоидии отсутствует одна (или несколько) хромосом, либо же хромосомы имеются в избытке ("лишние" хромосомы).
В случае отсутствия в хромосомном наборе одной хромосомы говорят о моносомии, двух хромосом - нуллисомии. Если к паре хромосом добавляется одна лишняя, говорят о трисомии.
Наследственные болезни, в том числе связанные с геномными мутациями: синдром Шерешевского-Тёрнера, Дауна - мы более детально обсудим в следующей статье, которая посвящена наследственным заболеваниям.
Раз уж мы затронули аутбридинг, то следует коснуться явления инбридинга и гетерозиса для их полного понимания.
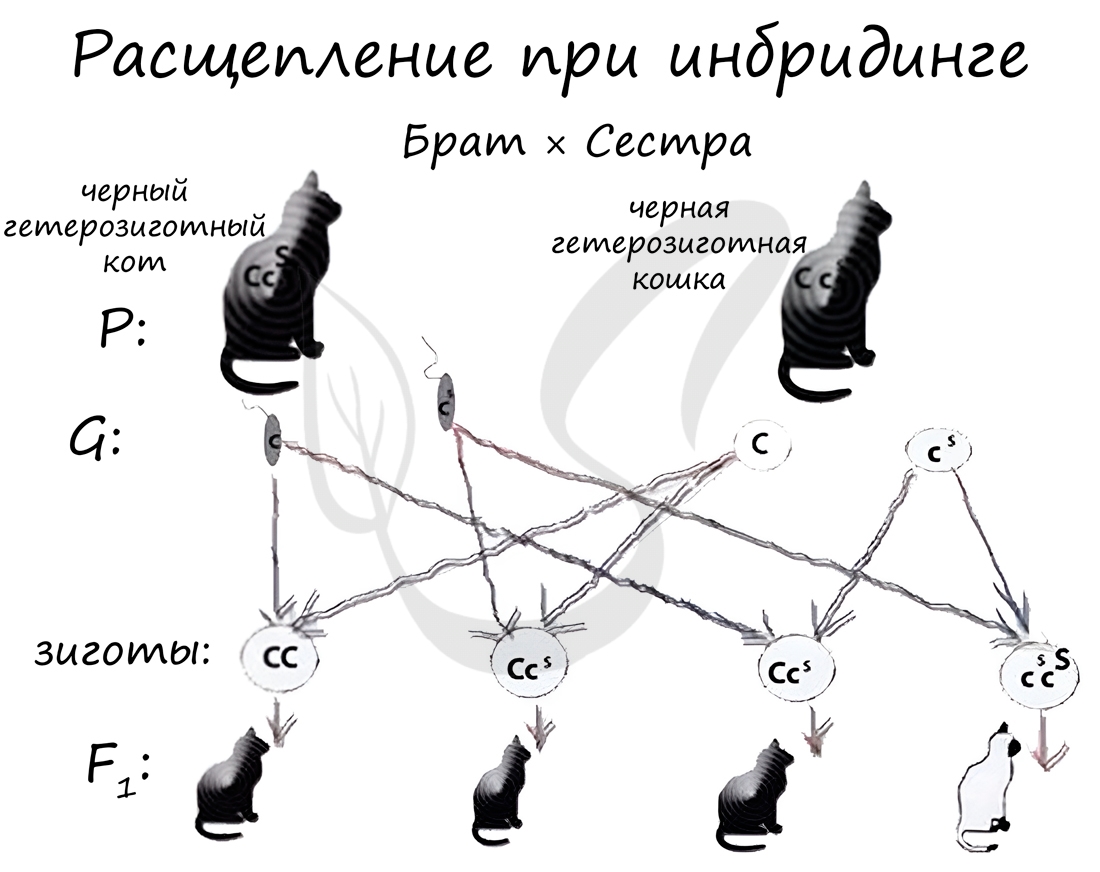
Инбридинг (англ. in — в, внутри + breeding — разведение) - скрещивание близкородственных форм, в результате которого в ряду поколений увеличивается гомозиготность. С помощью инбридинга выводят чистые линии (AA, aa, BB, bb). Однако известно, что близкородственное скрещивание может приводить к проявлению рецессивных генов заболеваний и ослаблению потомства.
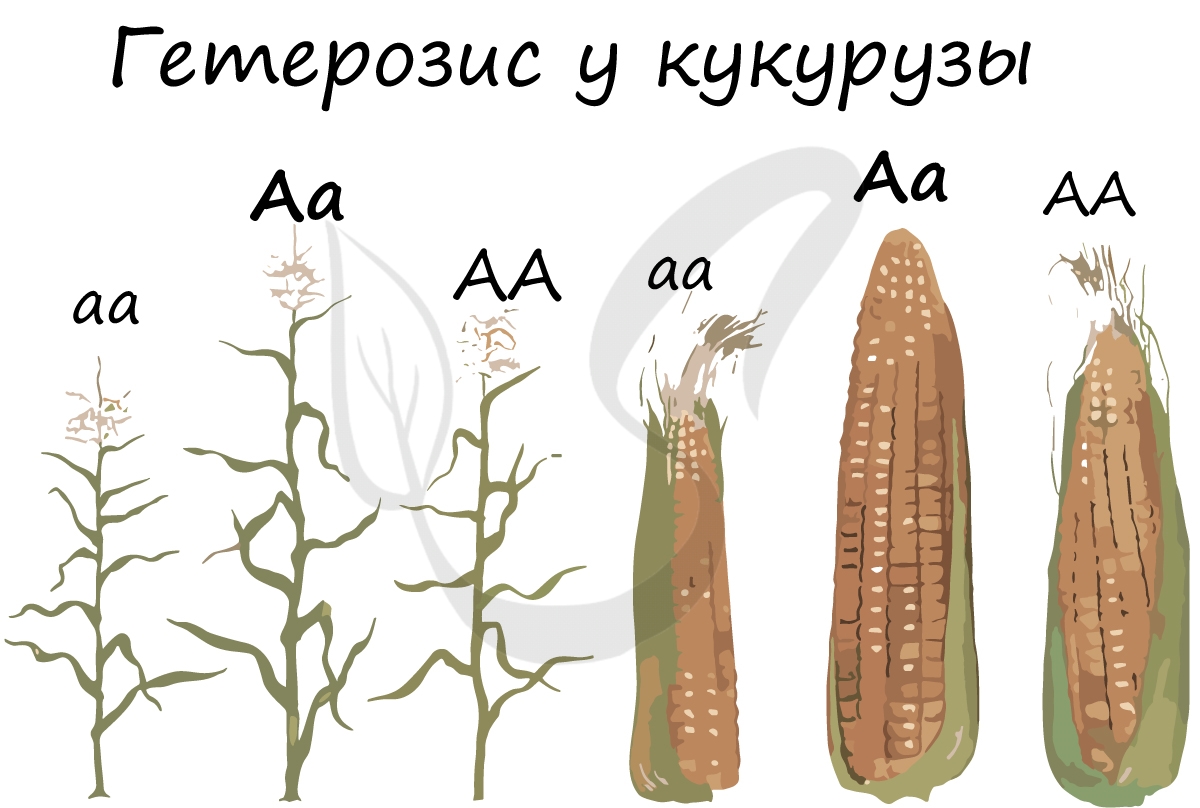
Гетерозис (греч. ἕτερος - другой + -ωσις - состояние) - явление увеличения жизнеспособности гибридов, вследствие унаследования ими различных вариантов аллельных генов от своих разнородных родителей. Увеличение жизнеспособности связывают с переходом генов в гетерозиготное состояние.
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Читайте также: