
Какие вирусы называют ретровирусами кратко
Обновлено: 07.07.2024
Основной биологической информацией, необходимой для построения и поддержания организма, является геном. В связи с этим особенно пугающим кажется тот факт, что в наших генах содержится 100 тыс. фрагментов ДНК эндогенных ретровирусов (ЭРВ), которые составляют 5–8% человеческого генома. Насколько это существенно, можно судить хотя бы по тому, что в кодировании основных белков нашего организма участвует 20 тыс. генов, что составляет только 1,2% нашей ДНК
Наши внутренние паразиты
Большинство эндогенных ретровирусов довольно старые, они встроились в геном наших предков свыше 25 млн лет назад. Это летопись следов наших встреч с вирусными инфекциями, которые заканчивались по-разному. Вирусы, которым удалось пробраться в половые клетки, получили возможность передаваться по наследству. В дальнейшем в результате мутаций эффективная экспрессия вирусных генов прекращается и ЭРВ превращаются в неактивный наследственный элемент генома. Сейчас все они находятся на той или иной стадии разрушения в разобранном состоянии.
Генетический мусор?
Ретровирусы представляют большую опасность для нашего здоровья, находясь в свободном состоянии, но даже после одомашнивания они по-прежнему могут представлять потенциальную угрозу
Герои плацентарной революции
Появление у древних организмов плаценты — важный этап их эволюционного развития в сторону усложнения. Плацента позволила предкам человека продлить внутриутробное развитие. Именно с этим сегодня связывают кардинальные изменения у млекопитающих, живших около 60 млн лет назад, — увеличение размеров мозга и постепенное развитие умственных способностей.
Роль в эволюции человеческого мозга
Ген PRODH кодирует фермент пролиндегидрогеназу, связанный с синтезом глутамата, одного из нейромедиаторов, стимулирующего передачу сигналов возбуждения в нервной системе. Есть основания полагать, что внедрение ретровируса вблизи этого гена сыграло весомую роль в развитии умственных способностей человека. Несомненно, что эта ретровирусная вставка имела большое значение в эволюции человеческого мозга. Эндогенные ретровирусы принимают активное участие в тонкой регуляции экспрессии генов мозга. Они помогают формировать наше мышление, участвуя в развитии мозга.
Онкологическая угроза
Тем не менее при упоминании о вирусах, обосновавшихся в наших генах, прежде всего предполагаются всевозможные угрозы и опасности, тем более что именно к семейству ретровирусов принадлежит патогенный вирус иммунодефицита человека (ВИЧ), вызывающий СПИД. Кроме того, некоторые из обнаруженных ретровирусов вызывают рак, но далеко не все. Ретровирусы представляют большую опасность для нашего здоровья, находясь в свободном состоянии, но даже после одомашнивания они по-прежнему могут представлять потенциальную угрозу.

Особую угрозу несут так называемые ксенотрансплантации — пересадки органов животных человеку. Как могут проявить себя ретровирусы животных, потенциально способные взаимодействовать с вирусами генома человека, трудно предсказать. Потому многие ученые выступают категорически против проведения таких трансплантаций. Кроме того, рассматривается связь эндогенных дельта-ретровирусов с рядом аутоиммунных заболеваний и, в частности, с рассеянным склерозом.
Опасная вирусная коалиция
Прогрессивное движение вспять
Ретровирусы и мобильные ретроэлементы являются действенным инструментом эволюции. Они возникли раньше нас в процессе эволюции, принимали активное участие в создании новых организмов, но при этом вполне способны стать причиной новых проблем для человечества. Иммунная система многоклеточных была создана ретроэлементами и может выполнять роль естественного резервуара для ретровирусов.
Ретровирусы представляют важную группу РНК-вирусов, которые вызывают ряд болезней нервной системы. Более того, потенциальную угрозу представляют вновь выявленные ретровирусные инфекции нервной системы и некоторые ритровирусные векторв или белки, используемые как переносчики генов. Данная статья описывает различные ретровирусы и их этиологическую роль в развитии болезней нервной системы, или, по другому, нейровирулентностъ. Специфические последовательности в пределах ретровирусных генов могут быть ответственны за нейровирулентностъ. Другой механизм заключается в активации иммунных реакций хозяина нейровирулентны-миретровирусами, что запускает нейропатогенетический каскад, медиаторами которого служат провоспалительные и нейротоксичные молекулы, а заканчивается все смертью нейрона. Таким образом, ретровирусные инфекции нервной системы иллюстрируют молекулярное взаимодействие между определенными вирусными возбудителями инфекций и патологическими реакциями хозяина; результатом такого взаимодействия является нейровирулентностъ.
Значение вирусных инфекций нервной системы все время возрастает, что видно по появлению за последние 5 лет нескольких глобальных нейрови-русных эпидемий [1]. Кроме того, повышается интерес в использовании вирусных векторов для генной экспрессии и для лечения заболеваний головного мозга [2]. Инфекцию считают нейровирусной, если она соответствует следующим критериям: нейроинвазивность, или проникновение вируса в нервную систему; нейротропизм (см. список терминов) или инфицирование вирусами клеток головного мозга, причем селективно инфицируются нейроны (это и называется нейротропизмом); и нейровирулентность или заболевание нервной системы, индуцированное вирусами. Хотя многие ретровирусы удовлетворяют этим критериям, но некоторые ретровирусные свойства усложняют их ней-робиологию, среди этих свойств: 1) предрасположенность к геномным мутациям; 2) способность вызывать как врожденный, так и адаптационный иммунный ответ в нервной системе; 3) способность вызывать патологические изменения за пределами нервной системы.
Структура и цикл жизни ретровирусов
Ретровирусы образуют большую семью оболочечных РНК-вирусов, которые паразитируют на многих видах (рис. 1а) [3]. Эти РНК-вирусы называются ретровирусами, потому что, для них характерна уникальная обратная транскрипция с РНК на вспомогательную ДНК провируса, которая затем интегрируется в хромосомную ДНК хозяина (рис. 1b). После этой интеграции провирус, путем транскрипции, синтезирует вирусные вещества, потенциально способные продлить жизнь клетке, что зависит от факторов транскрипции, продуцируемых клеткой хозяина или вирусом. Все ретровирусы, способные к репликации, имеют три главных открытых фрагментов считывания (в пределах генов gag, pol и env, - см. глоссарий) (рис. 1с). Однако, некоторые группы ретровирусов, такие как лентивирусы, содержат некоторые вспомогательные гены, которые регулируют вирусную репликацию, способность к инфицированию и транскрипцию. Непрочитанный длинный концевой повтор (long terminal repeat - LTR), который располагается на фланге открытых фрагментов считывания и действует как единица промоции, необходим для транскрипции всего вирусного генома и начала транскрипции отдельных вирусных генов. Ретровирус-ный геном представляет не что иное, как линейную молекулу РНК, размером примерно 10 kb [3], которая существует в виде дупликата в интактной вирусной частице или вирионе. Специфические ретровирусные белки, такие, как обратная транскриптаза, интеграза и протеаза кодируются в pol, и они необходимы, соответственно, для вирусной репликации, интеграции и сборки. Ген gag кодирует белки, необходимые для внутриклеточной сборки вируса и его высвобождения из клетки. Ген env кодирует структурные белки (структурная единица поверхности и трансмембранная структурная единица), которые необходимы для связывания с поверхностными молекулами клеток, служащих как рецепторы и медиирующих вход в клетку. У некоторых ретровирусов именно трансмембранные белки ответственны за нейровирулентность, убивая нейроны посредством прямых или косвенных механизмов [4], в последнем случае вначале инфицируются глиальные клетки, из которых затем высвобождаются нейротоксичные молекулы. В качестве непрямого механизма иногда действует активация (и инфицирование) клеток, ответственных за врожденный иммунитет (макрофаги, микроглия, астроциты) или адаптационный иммунитет (Т или В лимфоциты) в нервной системе.
Рис. 1. Ретровирусное филогенетическое дерево, жизненный цикл и репрезентативные структуры генома.
a) На филогенетическом дереве показано несколько нейротропных ретровирусов, в том числе - ретровирусы типа С [вирусы лейкозов грызунов (MuLV) и птиц(АLV), а также вирус, тропный к Т-лимфоцитам человека(НТLV)], лентивирусы [вирусы иммунодефицита человека (ВИЧ-1), обезьян (SIV), кошек (FIV) и вирус visna-maedi (VMV)], спумавирусы и эндогенные ретровирусы человека (HERV-K и HERV-C).
b) Жизненный цикл ретровирусов включает связывание вириона с рецептором клетки хозяина, вход в клетку, затем следует обратное транскрибирование с последующей интеграцией в геном хозяина. Вирусные транскрипты полной длины и отдельные вирусные мРНК синтезируются в ядре клетки, они служат информационным материалом для кодирования как структурных, так и регуляторных белков, которые создают предпосылки для самосборки вирусов и почкования новых вирионов.
c) В качестве наглядного примера представлены геномы ретровируса типа С (MuLV) и лентивируса (ВИЧ-1), показаны важнейшие структурные гены ретровирусов (gag, pol, env), отображена высокая степень сложности генома лентивирусов, обусловленная присутствием дополнительных генов, в том числе, -rev, tat, vif, vpu, nef, vpr.
Иммунологические нарушения, вызываемые некоторыми ретровирусами, приводят ко вторичным инфекциям за пределами нервной системы и к злокачественным процессам в нервной системе. Однако, настоящая статья рассматривает только прямую нейровирулентность ретровирусов, с особым вниманием к механизмам нейровирулентности и ней-ротропизма, кроме того, всесторонне описываются ретровирусы типа С (также именуемые онкогенными), в том числе, вирус лейкемии грызунов (MuLV) и вирусы, тропные к человеческим Т-лимфоцитам (HTLV), а также лентивирусы, такие, как вирусы иммунодефицита человека. Нейротропные
штаммы этих вирусов изучались широко. Они способны вызвать серьезные нарушения здоровья у человека.
Ретровирусы типа С (другое название - онкогенные ретровирусы)
Ретровирусы типа С (онкогенные) проявляют нейровирулентность в целом ряде биологических видов, таких, как человек, кошки, грызуны и птицы [4] (таблица 1), что иногда сопровождается параллельным поражением иммунитета. Структура генома этих вирусов проще, чем у лентивирусов, а именно, их геномы включают три структурных гена (два LTR и, в некоторых случаях, имеется добавочный ген). К нейротропным ретровирусам типа С относятся вирусы лейкоза грызунов (MuLV), кошек (FeLV) и птиц (ALV), а также вирусы, тропные к лимфоцитам Т у человека (HTVL-1 и HTVL-2).
Таблица 1. Типичные нейротропные ретровирусы: штаммы, нейропатология и клеточный тропизм.
| Группа ретровирусов | Вирус | Патоструктурные измененияв нервной системе а) | Клетки нервной системы инфицированные in vivo |
| Тип С | CasBrE | губчатость, глиоз (СМ) | эндотелий, нейроны, глия |
| NE-8 | губчатость, глиоз (СМ, СтМ, Мзж) | эндотелий, астроциты, нейроны | |
| FrCas е) | губчатость, глиоз (СМ, СтМ, Мзж) | эндотелий, микроглия, нейроны, олигодендроциты | |
| Fr98D | губчатость, глиоз (Мзж, БВ, СВ) | микроглия, нейроны (?) | |
| MoMuLV-tsl | губчатость, глиоз (СМ, СтМ, Тал, СВ,БВ) | эндотелий, астроциты, макрофаги | |
| PCV-211 | губчатость (Пол, СМ, СтМ, Мзж, БВ) | эндотелий | |
| TR1.3 | кровоизлияние (Мзж) | эндотелий | |
| LP-BM5 | глиоз (Пол, Мзж, БВ) | астроциты, микроглия, сосудистое сплетение | |
| WB91-GV | глиоз | олигодендроциты, астроциты | |
| HTLV-I/II | демилиенизация, глиоз, лимфоцитарнаяинфильтрация (СМ) | лимфоциты, астроциты | |
| Лентивирусы | вирус иммуно- дефицита человека | глиоз, микроглиальные узелки, многоядерные гигантские клетки, диффузноепобледнение миелина(СМ, Пол, Мзж, БВ, СВ, ПН) | макрофаги, микроглия, астроциты |
| вирус visna-maedi | глиоз, периваскулярное воспаление, демиелинизация (БВ) | макрофаги, микроглия, астроциты | |
| вирус иммуно- дефицита кошек | периваскулярное воспаление, глиоз, микроглиальные узелки (БВ, СВ, Мзж, СМ, ПН) | макрофаги, микроглия | |
| вирус карпального артрита/энцефалита | глиоз, демиелинизация (БВ) | макрофаги и микроглия | |
| вирус инфекционной анемии лошадей | эпендимит, периваскулрное воспаление (Мзж) | клетки периваскулярных зон и мозговых оболочек | |
| вирус иммуноде- фицита коров | периваскулярное воспаление (БВ) | глиальные клетки | |
| вирус иммуноде- фицита обезьян | периваскулярное воспаление, глиоз, многоядерные гигантские клетки (БВ, Мзж, СВ, СМ) | макрофаги, микроглия, астроциты, эндотелий |
а) Нейроструктурные изменения даны на основании гистологических заключений. Губчатые изменения и глиоз могут сопровождаться повреждением нервной системы и потерей ее структур, что может быть не заметным при гистопатологиче-ском исследовании.
Сокращения:
СтМ - ствол головного мозга; Мзж - мозжечок; Пол. - большие полушария; БВ - белое вещество; СВ - серое вещество; ПН - периферическая нейропатия; СМ - спинной мозг; Тал - таламус; БВ - белое вещество.
Вирусы MuLV
Вирусы MuLV образуют самую большую группу нейротропных ретровирусов. У пораженных грызунов развиваются разнообразные нарушения движений, - чаще всего отмечаются: тремор, паралич задних лап и отсутствие координации [5]. Некоторые штаммы MuLV, попадая в головной мозг, первично инфицируют глиальные клетки, такие, как микроглия, астроциты и олигодендроциты, хотя несколько MuLV инфицируют нейроны. Было выделено несколько рецепторов к MuLV на поверхности клеток [3], в том числе - переносчик катионных аминокислот, тСАТ-1 [6], но до сих пор не известие функционирует ли эта молекула как рецептор цен тральной нервной системы. Считается, что в естест венных условиях MuLV проникают в головно] мозг либо с помощью ими инфицированных лейко цитов, которые перемещаются по нервной системе либо путем инфицирования эндотелиальных кле ток головного мозга. Патологические изменения в нервной системе, вызываемые MuLV, также зависят от конкретного штамма, хотя чаще всего ветречаются "губчатые" изменения или потеря паренхимы ЦНС во всех секторах спинного мозга и ствола.
Следует отметить, что один вид MuLV вызывает геморрагические изменения, подобные инсульту, что отражает его селективный тропизм к эндотелиаль-ным клеткам (таблица 1).
Нейропатологические механизмы развития MuLV-инфекции являются примером взаимодействия разных последовательностей в геноме ретровируса, обычно расположенных в генах env или gag, и параллельно действует патологическая реакция организма-хозяина, определяемая генотипом мыши. Например, у штамма Fr-98 MuLV два отдельных участка гена env регулируют реализацию нейровирулентности в особых мышиных линиях [7], при этом нейровирулентность проявляется в разных нейроповеденческих нарушениях, приводящих к смерти. В отличие от этого штамма, его штамм-предшественник, хотя и имеет похожий клеточный тропизм (микроглия), но авирулентен в отношении нервной системы. У некоторых разновидностей MuLV нейровирулентности способствуют изменения во внутриклеточном цикле и в экспрессии белков MuLV, кодируемых геном env, а также изменения уровня вирусной репликации в головном мозге [7]. Патологические реакции хозяина, инициируемые инфекцией MuLV, остаются плохо изученными, но обнаруживалась корреляция между нейро-вирулентностью и повышенным про-воспалитель-ным образованием цитокинов в гиальных клетках [9]. К другим патологическим реакциям хозяина, индуцируемым инфекцией MuLV, относятся изменение проницаемости гематоэнцефалического барьера и повышение внеклеточной концентрации глутамата, что ведет к смерти нейрона посредством механизма внешней цитотоксичности [10].
Проведено картирование некоторых участков генов хозяина, ответственных за резистентность к неврологическому заболеванию, индуцируемому MuLV. К этим участкам относятся FV-1, FV-4 и Akvr-1, кодирующие эндогенные ретровирусные гены env и gag, которые мешают проявлениям нейровирулентности, так как связываются с предполагаемым рецептором к вирусу, или, может быть, блокируют этап после входа [5]. Кроме того, нейровирулентность, вызванная несколькими разновидностями MuLV, зависит от возраста хозяина, причем новорожденные имеют повышенную подверженность, ввиду слабости иммунной защиты.
Вирусы HTVL
HTVL-1 и HTVL-2 - это ретровирусы, которые вызывают неврологические нарушения или неопла-зию у человека, впрочем, одновременно эти два вида нарушений встречаются редко [11]. У небольшой (
Ретровирусы. Семейство ретровирусов. Виды ретровирусов.
Ретровирусы содержат две идентичные молекулы геномной РНК и РНК-зависимую ДНК-полимеразу (обратную транскриптазу, ревертазу). Ретровирусы выделены от множества видов животных и проявляют разнообразный спектр патогенного потенциала. Семейство включает вирусы 7 родов: альфа-, бета-, гамма-, дельта-, эпсилонретровирусы, лентивирусы и спумавирусы. Семейство включает вирусы, патогенные для человека и многих видов животных.
Большинство ретровирусов обладают выраженным тропизмом к клеткам лимфОретикулярной и гемопоэтической системы. Ретровирусы у специфических для них видов животных встречаются повсеместно. В борьбе с ретровирусными инфекциями основной мерой является предотвращение передачи вируса.
Вирионы ретровирусов представляют собой округлые оболочечные частицы диаметром 80—100 нм, обладающие уникальной трехслойной структурой. Центральная часть вириона представлена нуклеопротеиновым комплексом, который включает около 30 молекул ревертазы и имеет спиральную симметрию. Эта структура окружена икосаэдрическим капсидом диаметром около 60 нм, покрытым оболочкой, происходящей из мембраны клетки, от которой отходят гликопротеиновые пепломеры. Лентивирусы имеют на поверхности примерно 72 шишкоподобных пепломера длиной около 10 нм с яйцевидным концевым уплотнением.
Ретровирусы имеют диплоидный геном, представляющий инвертированный димер из двух молекул линейной позитивной полярности, оцРНК; каждая молекула содержит 7-11 тн и имеет полиА последовательность на З'-конце и КЭП-структуру на 5'-конце. Детальная организация геномов разных ретровирусов широко варьирует.
Геном ретровирусов уникален в следующих отношениях:
1) является единственным диплоидным;
2) вирусная РНК синтезируется и изменяется с помощью механизма, изменяющего клеточную мРНК;
3) это единственный геном, связанный со специфическим переносом функции РНК целиком к первичной репликации;
4) это единственная оц(+)РНК, которая не является мРНК вскоре после инфекции;
5) это единственный геном, кодирующий обратную транскриптазу, которая сама по себе уникальна.
Среди многих своих функций обратная транскриптаза служит в качестве РНК-зависимой ДНК полимеразы, а ДНК-зависимая ДНК полимераза, интеграза и РНКаза, каждая в меру своей определенной функции, представляют различную часть белковой молекулы. Геном недефектных ретровирусов содержит три разных гена, каждый из которых кодирует два и более белка. Gag ген кодирует вирионные коровью белки, pol ген кодирует обратную транскриптазу, a env ген кодирует вирионные пепломерные белки. Геном кодирует также несколько определенных вирионных компонентов.
Геном быстро трансформирующих ретровирусов содержит четвертый ген — вирусный онкоген (v-onc). Присутствие онкогена обычно связано с делецией в другом месте генома, обычно в env гене, так что большинство v-onc содержащих вирусов не способны синтезировать полностью оболочку и поэтому являются дефектными по репликации. Они всегда находят связь с недефектными вирусами, которые реплицируются полностью и функционируют в качестве помощников. Вирус саркомы Рауса является исключением. Его геном содержит вирусный онкоген v-src и также полные gag, pol и env гены, и поэтому является репликативнокомпетентным.
Vpu, vpr и vpx найдены в лентивирусах приматов. Vpu способствует созреванию вирусных гликопротеинов и освобождению вирионов почкованием; Vpr -слабый активатор транскрипции; Vpx, по-видимому, способствует репликации в Т-лимфоцитах и макрофагах.
При продуктивной инфекции вирионы формируются и освобождаются почкованием через плазматическую мембрану. Некоторые ретровирусы вызывают образование опухолей.
Спумавирусы являются необычными ретровирусами во многих отношениях. Они выделены от человека, обезьян, КРС, кошек, морских львов и хомяков. Они представляют проблему, когда контаминируют клеточные культуры, но их связь с заболеваниями неизвестна. Существует мнение, что пенящие вирусы ближе стоят к ДНК гепаднавирусам.
Эндогенный ретровирус свиней (PERV) найден у всех исследованных пород свиней. Известны три субгруппы этого вируса: А, В и С. Все свиньи содержат множественные провирусы (около 50) в своем геноме. PERVлатентно инфицирует многие культуры клеток свиньи и, реже, человека. Патогенность этого вируса не установлена.
Онковирусы, в отличие от лентивирусов, реплицируются в делящихся клетках. В составе ретровирусов обнаружено 11-13 белков. Часть из них локализована в сердцевине, а главный внутренний белок Р27 - Р30 - в мембране, окружающей РНК. В составе липопротеиновой оболочки 2-5 гликопротеинов, из которых более крупный образует головку шипика и является главным компонентом оболочки, а более мелкие белки образуют отросток, погруженный в билипидный слой. Различные белки ответственны за групповую и типовую специфичность. Подгрупповые и типоспецифические антигены связаны с наружными гликопротеинами и выявляются в РН, РСК, ИФ и РИД. Группоспецифические антигены связаны с белками сердцевины. Их обнаруживают в РСК, ИФ и РИД. Главный внутренний белок Р27 - РЗО - основной группоспецифический антиген, общий для всех ретровирусов птиц или млекопитающих, относящихся к одной группе.
Согласно предложенной ранее номенклатуре структурные белки ретровирусов обозначали следующим образом: МА - матриксный, СА - капсидный, NC -белок нуклеокапсида, SU - поверхностный, ТМ - трансмембранный, PR- протеаза, RT - ревертаза, IN - белок интеграции.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.

Уже много лет между эволюционистами и креационистами продолжается спор о том, как же все-таки произошел человек — путем эволюции или божественного сотворения? К счастью, древние вирусы на протяжении миллионов лет ведут летопись эволюции и записывают ее в нашу ДНК.
После расшифровки нуклеотидной последовательности ДНК многих животных, в том числе и человека, стало возможным узнать, где именно в геноме находятся эти остатки древних вирусов. И взору ученых предстала строгая упорядоченность расположения эндогенных ретровирусов — выяснилось, что все они находятся в геномах в строго определенных местах. Некоторые из них характерны лишь для человека или для кошки и не встречаются у других животных. Другие же ретровирусы можно обнаружить в одном и том же месте, к примеру, в геномах гориллы, шимпанзе, орангутанга и человека.
Почти половина генома

У разных ретротранспозонов (транспозон — мобильный элемент ДНК) процесс обратной транскрипции имеет свои особенности.
У ретротранспозонов с длинными концевыми повторами (LTR-ретропозоны) обратная транскрипция происходит не в ядре, а в цитоплазме. Так как по своему строению и механизму перемещения LTR-ретропозоны имеют большое сходство с вирусами, данный класс подвижных элементов называют ретровирусоподобными. Их содержание в геноме человека — около 8% всей последовательности нуклеотидов.
Ко второму классу ретротранспозонов, без длинных концевых повторов (non-LTR), относятся элементы LINE (Long Interspersed Elements — длинные перемежающиеся элементы) и SINE (Short Interspersed Elements — короткие перемежающиеся элементы). Перемещение и встраивание ДНК-копии этих элементов происходит не в цитоплазме, а в ядре. Элементы LINE — самые многочисленные из подвижных структур человека: они занимают в ДНК пятую часть (около 20%) от всей последовательности нуклеотидов. И они же единственные из мобильных генов человека, сохранившие до сих пор свою самостоятельную способность к перемещению.
Одомашненные вирусы
Транскрипция — это перенос генетической информации с ДНК на рибонуклеиновую кислоту (РНК), при котором ДНК используется в качестве отправной точки, матрицы. Транскрипцию можно наблюдать всякий раз, когда осуществляется синтез новых белков. До открытия американцами обратной транскрипции считалось, что движение в направлении от РНК к ДНК невозможно. Но, как оказалось, этот генетический метод активно используется в живой природе, в том числе и такими опасными ее представителями, как вирусы (среди которых и самый опасный для человека — ВИЧ).
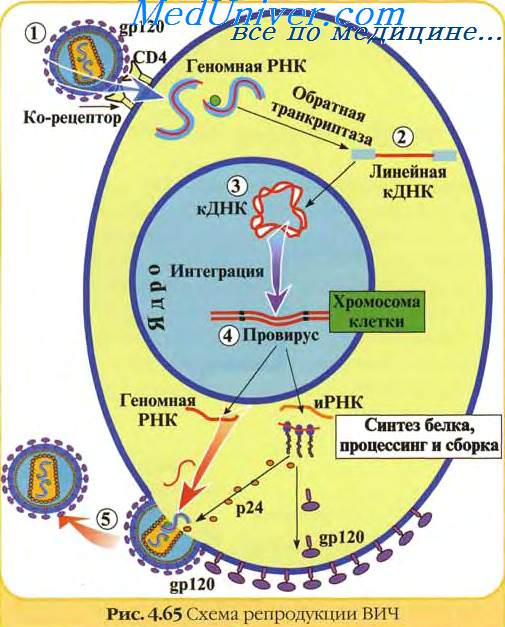
Жизненный цикл ретровируса

Вирус прикрепляется к строго определенным клеткам хозяина благодаря образованию связей белков капсида и рецепторов на поверхности клетки. После проникновения в клетку собственные ферменты или ферменты клетки хозяина разбирают капсид. Вирусная РНК высвобождается и подвергается обратной транскрипции: обратная транскриптаза формирует по матрице РНК цепочки ДНК, а интеграза инициирует проникновение провирусной ДНК в ядро и включение ее в геном хозяина. В ядре происходит процесс репликации (повторной сборки) вирусной РНК, который уже стал неотъемлемой функцией генома хозяина. В хозяйской цитоплазме вирусная РНК обзаводится капсидом. Отпочковываясь от клетки, обновленный вирус прихватывает с собой часть мембраны хозяина, используя ее в качестве собственной оболочки.
Устройство вириона

РНК ретровируса располагается в белковой оболочке под названием капсид. Наружная липидная оболочка покрыта ворсинками длиной 8−10 нм. Вирион имеет форму икосаэдра (двадцатигранника) и диаметр 80−100 нм.
Двигатель прогресса
Еще в конце 1980-х годов можно было встретить утверждение о том, что ретровирусы не способны вызывать эпидемический процесс. И отсюда — бесплодные попытки ученых создать вакцину против ВИЧ. Ретровирусы и сегодня не утратили своей способности вызвать большую пандемию. Однако ретровирусы, по мнению ученых, могут быть полезными. Предполагается, и не без оснований, что они сослужили нам хорошую службу в процессе эволюции, передав человеку и другим живым организмам свои структурные элементы, ставшие впоследствии нашими генами.

Как мы строим супервирус
Сегодня уже точно известен целый ряд важных генов, берущих свое начало от ретроэлементов. Прежде всего это некоторые гены, участвующие во внутриутробном развитии плода. Несколько лет назад появились данные, что ретровирусы могли сыграть весомую роль в эволюции плацентарных животных. Появление у древних организмов плаценты — важный этап их эволюционного развития в сторону усложнения. Плацента позволила предкам человека продлить внутриутробное развитие. Именно с этим сегодня связывают кардинальные изменения у млекопитающих, живших около 60 млн лет назад, — увеличение размеров мозга и постепенное развитие умственных способностей.
Основной целью российских биологов был поиск ретровирусов hsERV, которые осуществляют функции энхансеров (усилителей). Энхансеры — это нуклеотидная последовательность с регуляторными функциями, которая обычно находится вблизи (или внутри) генов и повышает их экспрессию. Из всех обнаруженных на сегодня hsERV лишь шесть копий находились в районах обычного расположения энхансеров. Изучив эти шесть ретровирусов, исследователи смогли выявить один hsERV, расположенный вблизи важного гена PRODH.
Ген PRODH кодирует фермент пролиндегидрогеназу, связанный с синтезом глутамата, одного из нейромедиаторов, стимулирующего передачу сигналов возбуждения в нервной системе. У шимпанзе аналогичный ген во всех местах его расположения (в гиппокампе, префронтальной коре и хвостатом ядре) не имеет рядом с собой участка ДНК с эндогенными ретровирусами и менее активен по сравнению с человеческим. Есть основания полагать, что внедрение ретровируса вблизи этого гена сыграло весомую роль в развитии умственных способностей человека.
Хозяева Земли
По мнению Супотницкого, именно ретровирусы (и ретроэлементы) — настоящие хозяева Земли. Они возникли раньше нас в процессе эволюции, принимали активное участие в создании сложных организмов и вполне способны ради большего разнообразия видов сгубить все человечество.
Автор выражает большую благодарность
д. б. н. А. А. Буздину (Институт биоорганической химии РАН)
за помощь при написании статьи.
Читайте также: