
Как осуществляется движение у бактерий кратко
Обновлено: 05.07.2024
Среди бактерий встречаются подвижные и неподвижные формы. Движение бактерий происходит обычно при помощи так называемых жгутиков. Некоторые извитые бактерии, не имеющие жгутиков, передвигаются путем изгибания тела. Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие начало от цитоплазматической мембраны, имеют большую длину, чем сама клетка. Толщина жгутиков 12-20 нм, длина 3-15 мкм. Жгутики состоят из белка - флагеллина (от flagellum – жгутик).
Расположение жгутиков на теле бактерии может быть различным: один жгутик на конце тела (бактерии-монотрихи), пучок жгутиков на конце тела (бактерии-лофотрихи), жгутики расположены по всей поверхности тела (бактерии-перитрихи) у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков на одном из концов клетки. Амфитрихи имеют по одному жгутику или пучку жгутиков на противоположных концах клетки.
Характер движения определяется характером жгутования. Бактерии с полярно расположенными жгутиками движутся по прямой, только иногда делая отклонения в сторону в виде легких колебательных движений. Движение бактерий, имеющих жгутики по всему телу, носит беспорядочный характер, происходит в виде оживленного кувыркания.
Скорость перемещения у разных бактерий различна. Наиболее подвижным считается холерный вибрион, который за 1 секунду проходит расстояние, в 15 раз превышающее длину его тела. Большая же часть бактерий за 1 секунду перемещается на расстояние, близкое длине их тела. На подвижность бактерий очень сильное влияние оказывают условия внешней среды.
По способности передвигаться все бактерии делятся на две группы – неподвижные и подвижные (примерно 1/20 часть). Броуновское движение присуще Неподвижным бактериям размером менее 4 мкм присуще броуновское движение.
Движение с помощью жгутиков. Это наиболее распространенный тип движения. По количеству и характеру расположения различают жгутики:полюсные, когда один, два и более жгутиков расположены на одном (монополярно) или обоих (биполярно) полюсах клетки и основание жгутика обычно параллельно длинной оси клетки; подполюсные жгутики (или субполярные), когда один, два и более жгутиков расположены в месте перехода боковой поверхности в полюс клетки на одном или двух концах ее, и основание жгутика обычно составляет прямой угол с длинной осью клетки; боковые (или латеральные) жгутики, когда один, два и более жгутиков в виде кустика, расположены в средней точке одной из половин клетки; перитрихальные жгутики, которые расположены по всей поверхности клетки по одному или пучками (обычно полюса клеток лишены их); смешанныежгутики, когда два или несколько жгутиков расположены в различных точках клетки.
Количество жгутиков зависит от вида микроорганизма и условий культивирования. Жгутики очень тонкие образования, состоящие в основном (95%) из белка флагеллина (лат. flagellum– жгутик).
Бактерии, имеющие один жгутик, называются монотрихами, пучок жгутиков – лофотрихами (политрихами). Биполярные монотрихи или лофотрихи называются амфитрихами.
Большинство бактерий за секунду проходит расстояние, близкое длине их тела, но есть бактерии, движущиеся с большой скоростью (до 30 мкм/с). Скорость движения не зависит от количества жгутиков, но зависит от характера расположения жгутиков (с терминальным расположением движутся быстрее, чем с перитрихальным), зависит также от свойств среды (вязкости, температуры, рН, осмотического давления и пр.). Направленность движения обуславливается характером расположения жгутиков: моно-и лофотрихи движутся прямолинейно, перитрихи – беспорядочно, кувыркаются.
Скользящий тип движения менее распространен у бактерий. Он присущ миксобактериям, некоторым микоплазмам, цитофагам. Скорость низкая (2-11 мкм/с). большинство скользящих бактерий выделяет слизь.
Вращательный тип движения присущ извитым формам, например, спирохетам. Они передвигаются за счет колебания собственного тела, очень подвижны.
Таксическое движение. Подвижные бактерии могут осуществлять направленные передвижения – таксисы (гр.taxis – расположение), обусловленные различными внешними факторами – стимулами. Таксисы бывают положительные или отрицательные в зависимости от движения бактерий к фактору или от него. Различают несколько видов таксисов.
Хемотаксис – движение, вызываемое химическими веществами. К хемотаксису относится аэротаксис, обусловленный кислородом, и осмотаксис, обусловленный концентрацией солей. По способности индуцировать положительный или отрицательный хемотаксис различают две группы веществ: аттрактанты– вещества, вызывающие скопления клеток в области более высокой концентрации соединения, и репелленты – вызывающие скопления клеток в области наименьшей концентрации вещества.
Вискозитаксис – движение бактерий в направлении увеличения или уменьшения вязкости раствора. Термотаксис – движение бактерий, вызванное источником тепла. Фототаксис– движение бактерий, обусловленное световой энергией.
Магнетотаксис – движение бактерий по силовым линиям магнитного поля Земли или магнита. Магнетотаксис обусловлен наличием у бактерий магнитосом(частички феррина – Fe304, окруженные мембраной), выполняющих роль магнитной стрелки. Ферромагнитные микрокристаллы образуются в клетках бактерий в количестве 10-20 размером 40-90 нм. Они свойственны магнитотропным бактериям, обитающим в донных осадках морских и пресноводных водоемов.
По способности передвигаться все бактерии делятся на две группы – неподвижные и подвижные (примерно 1/20 часть). Броуновское движение присуще Неподвижным бактериям размером менее 4 мкм присуще броуновское движение.
Движение с помощью жгутиков. Это наиболее распространенный тип движения. По количеству и характеру расположения различают жгутики:полюсные, когда один, два и более жгутиков расположены на одном (монополярно) или обоих (биполярно) полюсах клетки и основание жгутика обычно параллельно длинной оси клетки; подполюсные жгутики (или субполярные), когда один, два и более жгутиков расположены в месте перехода боковой поверхности в полюс клетки на одном или двух концах ее, и основание жгутика обычно составляет прямой угол с длинной осью клетки; боковые (или латеральные) жгутики, когда один, два и более жгутиков в виде кустика, расположены в средней точке одной из половин клетки; перитрихальные жгутики, которые расположены по всей поверхности клетки по одному или пучками (обычно полюса клеток лишены их); смешанныежгутики, когда два или несколько жгутиков расположены в различных точках клетки.

Количество жгутиков зависит от вида микроорганизма и условий культивирования. Жгутики очень тонкие образования, состоящие в основном (95%) из белка флагеллина (лат. flagellum– жгутик).
Бактерии, имеющие один жгутик, называются монотрихами, пучок жгутиков – лофотрихами (политрихами). Биполярные монотрихи или лофотрихи называются амфитрихами.
Большинство бактерий за секунду проходит расстояние, близкое длине их тела, но есть бактерии, движущиеся с большой скоростью (до 30 мкм/с). Скорость движения не зависит от количества жгутиков, но зависит от характера расположения жгутиков (с терминальным расположением движутся быстрее, чем с перитрихальным), зависит также от свойств среды (вязкости, температуры, рН, осмотического давления и пр.). Направленность движения обуславливается характером расположения жгутиков: моно-и лофотрихи движутся прямолинейно, перитрихи – беспорядочно, кувыркаются.
Скользящий тип движения менее распространен у бактерий. Он присущ миксобактериям, некоторым микоплазмам, цитофагам. Скорость низкая (2-11 мкм/с). большинство скользящих бактерий выделяет слизь.
Вращательный тип движения присущ извитым формам, например, спирохетам. Они передвигаются за счет колебания собственного тела, очень подвижны.
Таксическое движение. Подвижные бактерии могут осуществлять направленные передвижения – таксисы (гр.taxis – расположение), обусловленные различными внешними факторами – стимулами. Таксисы бывают положительные или отрицательные в зависимости от движения бактерий к фактору или от него. Различают несколько видов таксисов.
Хемотаксис – движение, вызываемое химическими веществами. К хемотаксису относится аэротаксис, обусловленный кислородом, и осмотаксис, обусловленный концентрацией солей. По способности индуцировать положительный или отрицательный хемотаксис различают две группы веществ: аттрактанты– вещества, вызывающие скопления клеток в области более высокой концентрации соединения, и репелленты – вызывающие скопления клеток в области наименьшей концентрации вещества.
Вискозитаксис – движение бактерий в направлении увеличения или уменьшения вязкости раствора. Термотаксис – движение бактерий, вызванное источником тепла. Фототаксис– движение бактерий, обусловленное световой энергией.
Магнетотаксис – движение бактерий по силовым линиям магнитного поля Земли или магнита. Магнетотаксис обусловлен наличием у бактерий магнитосом(частички феррина – Fe304, окруженные мембраной), выполняющих роль магнитной стрелки. Ферромагнитные микрокристаллы образуются в клетках бактерий в количестве 10-20 размером 40-90 нм. Они свойственны магнитотропным бактериям, обитающим в донных осадках морских и пресноводных водоемов.
Бактерии активно передвигаются в направлении, определяемом теми или иными раздражителями. Это явление получило название таксис.
4. Метаболизм
Конструктивный метаболизм

Рис. 4.1. Pseudomonas fluorescens За исключением некоторых специфических моментов биохимические пути, по которым осуществляется синтез белков, жиров, углеводов и нуклеотидов, у бактерий схожи с таковыми у других организмов. Однако по числу возможных вариантов этих путей и, соответственно, по степени зависимости от поступления органических веществ извне они различаются. Часть из них может синтезировать все необходимые им органические молекулы из неорганических соединений (автотрофы), другие же требуют готовых органических соединений, которые они способны лишь трансформировать (гетеротрофы).
Наибольшей степенью гетеротрофности отличаются внутриклеточные паразиты. Если при этом они способны существовать на богатых искусственных средах, они называются факультативными (факультативными иногда также называют паразитов, способных проделывать весь свой жизненный цикл во внешней среде, без участия хозяина). Некоторые облигатные (обязательные) внутриклеточные паразиты утратили часть путей биосинтеза и получают многие органические вещества, вплоть до АТФ, из клеток хозяина. Велика степень зависимости от хозяев также многих бактерий-эндосимбионтов. Большинство бактерий принадлежит к сапрофитам: они не питаются непосредственно веществами других организмов, но используют синтезированные ими органические вещества после их смерти. Существует также ряд бактерий, требующих наличия в среде небольшого круга определённых органических веществ (аминокислот, витаминов), которых они не могут синтезировать самостоятельно и, наконец, гетеротрофы, которые нуждаются лишь в одном довольно низкомолекулярном источнике углерода (сахар, спирт, кислота). Некоторые из них отличаются высокой специализацией (Bacillus fastidiosus может использовать только мочевую кислоту), другие в качестве единственного источника углерода и энергии могут использовать сотни различных соединений (многие Pseudomonas (рис. 4.1)).
Удовлетворять потребности в азоте бактерии могут как за счёт его органических соединений (подобно гетеротрофным эукариотам), так и за счёт молекулярного азота (как и некоторые археи). Большинство бактерий используют для синтеза аминокислот и других азотсодержащих органических веществ неорганические соединения азота: аммиак (поступающий в клетки в виде ионов аммония), нитриты и нитраты (которые предварительно восстанавливаются до ионов аммония). Фосфор они способны усваивать в виде фосфата, серу — в виде сульфата или реже сульфида.
Энергетический метаболизм
Способы же получения энергии у бактерий отличаются своеобразием. Существует три вида получения энергии (и все три известны у бактерий): брожение, дыхание и фотосинтез.
Брожение — серия окислительно-восстановительных реакций, в ходе которых образуются нестабильные молекулы, с которых остаток фосфорной кислоты переносится на АДФ с образованием АТФ (субстратное фосфорилирование). При этом возможно внутримолекулярное окисление и восстановление.
Дыхание — окисление восстановленных соединений с переносом электрона через локализованную в мембране дыхательную электронтранспортную цепь , создающую трансмембранный градиент протонов, при использовании которого синтезируется АТФ (окислительное фосфорилирование). В то время как эукариоты в конечном итоге почти всегда "сбрасывают" электрон на кислород (лишь в редких случаях акцептором электронов могут служить нитраты), бактерии могут использовать вместо него окисленные органические и минеральные соединения (фумарат, углекислый газ, сульфат-анион, нитрат- анион и др.; см. анаэробное дыхание), а вместо окисляемого органического субстрата использовать минеральный (водород, аммиак, сероводород и др.), что часто бывает сопряжено с автотрофной фиксацией CO2.
Фотосинтез бактерий может быть двух типов — бескислородный, с использованием бактериохлорофилла (зелёные, пурпурные и гелиобактерии) и кислородный с использованием хлорофилла (цианобактерии (хлорофилл a), прохлорофиты (a и b)). Цианобактерии (рис.4.2), глаукоцистофитовые, красные и криптофитовые водоросли — единственные фотосинтезирующие организмы, содержащие фикобилипротеины. У архей встречается бесхлорофилльный фотосинтез с участием бактериородопсина (правда, энергия света используется при этом не для фиксации CO2 , а непосредственно для синтеза АТФ, так что в строгом смысле это не фотосинтез, а фотофосфорилирование).
5. Размножение и устройство генетического аппарата бактерий
Размножение бактерий

Рис.5.1. Электронограммы ультратонких срезов делящихся бактерий: а — стафилококка, делящегося путем образования перегородки деления (указана стрелками), 32 000; б — кишечной палочки — в результате формирования перетяжки деления (указана стрелками); 1 — клеточная стенка, 2 — цитоплазматическая мембрана, 3 — нуклеоид; ´ 22 000. Некоторые бактерии не имеют полового процесса и размножаются лишь равновеликим бинарным поперечным делением (рис.5.1) или почкованием. Для одной группы одноклеточных цианобактерий описано множественное деление (ряд быстрых последовательных бинарных делений, приводящий к образованию от 4 до 1024 новых клеток). Для обеспечения необходимой для эволюции и приспособления к изменчивой окружающей среде пластичности генотипа у них существуют иные механизмы.
При делении большинство грамположительных бактерий и нитчатых цианобактерий синтезируют поперечную перегородку от периферии к центру при участии мезосом. Грамотрицательные бактерии делятся путём перетяжки: на месте деления обнаруживается постепенно увеличивающееся искривление ЦПМ и клеточной стенки внутрь. При почковании на одном из полюсов материнской клетки формируется и растёт почка, материнская клетка проявляет признаки старения и обычно не может дать более 4 дочерних. Почкование имеется у разных групп бактерий и, предположительно, возникало несколько раз в процессе эволюции.

Рис. 5.2. Половое размножение бактерий У бактерий наблюдается и половое размножение (рис.5.2), но в самой примитивной форме. Половое размножение бактерий отличается от полового размножения эукариот тем, что у бактерий не образуются гаметы и не происходит слияния клеток. Однако главнейшее событие полового размножения, а именно обмен генетическим материалом, происходит и в этом случае. Этот процесс называется генетической рекомбинацией. Часть ДНК (очень редко вся ДНК) клетки-донора переносится в клетку-реципиент, ДНК которой генетически отличается от ДНК донора. При этом перенесённая ДНК замещает часть ДНК реципиента. В процессе замещения ДНК участвуют ферменты, расщепляющие и вновь соединяющие цепи ДНК. При этом образуется ДНК, которая содержит гены обеих родительских клеток. Такую ДНК называют рекомбинантной. У потомства или рекомбинантов, наблюдается заметное разнообразие признаков, вызванное смещением генов. Такое разнообразие признаков очень важно для эволюции и является главным преимуществом полового размножения. Известны 3 способа получения рекомбинантов. Это — в порядке их открытия — трансформация , конъюгация и трансдукция .
Генетический аппарат (рис. 5.3)

Рис. 5.3. Генетический аппарат бактерии Гены, необходимые для жизнедеятельности и определяющие видовую специфичность, расположены у бактерий чаще всего в единственной ковалентно замкнутой молекуле ДНК — хромосоме (иногда для обозначения бактериальных хромосом, чтобы подчеркнуть их отличия от эукариотических, используют термин генофор (англ. genophore)). Область, где локализована хромосома, называется нуклеоид и не окружена мембраной. В связи с этим новосинтезированная мРНК сразу доступна для связывания с рибосомами, а транскрипция и трансляция сопряжены.
Помимо хромосомы, в клетках бактерий часто находятся плазмиды — также замкнутые в кольцо ДНК, способные к независимой репликации . Они могут быть настолько велики, что становятся неотличимы от хромосомы, но содержат дополнительные гены, необходимые лишь в специфических условиях. Специальные механизмы распределения обеспечивают сохранение плазмиды в дочерних клетках так что они теряются с частотой менее 10 -7 в пересчёте на клеточный цикл. Специфичность плазмид может быть весьма разнообразной: от одного вида-хозяина до плазмиды RP4, встречающейся почти у всех грамотрицательных бактерий. В них кодируются механизмы устойчивости к антибиотикам, разрушения специфических веществ и т. д., nif-гены, необходимые для азотфиксации также находятся в плазмидах. Ген плазмиды может включаться в хромосому с частотой около 10 -4 — 10 -7 .
В ДНК бактерий, как и в ДНК других организмов, выделяются транспозоны — мобильные сегменты, способные перемещаться из одной части хромосомы к другой, или во внехромосомные ДНК (в том числе в другие клетки). В отличии от плазмид, они неспособны к автономной репликации, и содержат IS-сегменты — участки, которые кодируют свой перенос внутри клетки. IS-сегмент может выступать в роли отдельной транспозоны.
Горизонтальный перенос генов
У прокариот может происходить частичное объединение геномов. При конъюгации клетка-донор в ходе непосредственного контакта передаёт клетке-реципиенту часть своего генома (в некоторых случаях весь). Участки ДНК донора могут обмениваться на гомологичные участки ДНК реципиента. Вероятность такого обмена значима только для бактерий одного вида.
Аналогично бактериальная клетка может поглощать и свободно находящуюся в среде ДНК, включая её в свой геном в случае высокой степени гомологии с собственной ДНК. Данный процесс носит название трансформация. В природных условиях протекает обмен генетической информацией при помощи умеренных фагов (трансдукция). Кроме этого, возможен перенос нехромосомных генов при помощи плазмид определённого типа, кодирующих этот процесс, процесс обмена другими плазмидами и передачи транспозон.
При горизонтальном переносе новых генов не образуется (как то имеет место при мутациях), однако осуществляется создание разных генных сочетаний. Это важно по той причине, что естественный отбор действует на всю совокупность признаков организма.
6. Клеточная дифференциация

Рис.6.1 Формирование эндоспоры спорообразующими бактериями: I - вегетативная клетка; II - инвагинация ЦПМ ; III - образование споровой перегородки (септы); IV - формирование двойной мембранной системы образующейся проспоры; V - сформированная проспора; VI - формирование кортекса; VII - формирование покровов споры; VIII - лизис материнской клетки; IX - свободная зрелая спора; X - прорастание споры Клеточная дифференциация — изменение набора белков (обычно также проявляющееся в изменении морфологии) при неизменном генотипе.
Образование покоящихся форм
Образование особо устойчивых форм с замедленным метаболизмом, служащих для сохранения в неблагоприятных условиях и распространения (реже для размножения) является наиболее распространённым видом дифференциации у бактерий. Наиболее устойчивыми из них являются эндоспоры (рис.6.1), формируемые представителями Bacillus, Clostridium, Sporohalobacter, Anaerobacter (образует 7 эндоспор из одной клетки и может размножаться с их помощью) и Heliobacterium. Образование этих структур начинается как обычное деление и на первых стадиях может быть превращено в него некоторыми антибиотиками. Эндоспоры многих бактерий способны выдерживать 10-минутное кипячение при 100 °C, высушивание в течение 1000 лет и, по некоторым данным, сохраняются в почвах и горных породах в жизнеспособном состоянии миллионы лет.
Менее устойчивыми являются экзоспоры, цисты (Azotobacter, скользящие бактерии и др.), акинеты (цианобактерии) и миксоспоры (миксобактерии).
Другие типы морфологически дифференцированных клеток
Актиномицеты и цианобактерии образуют дифференцированные клетки, служащие для размножения (споры, а также гормогонии и баеоциты соответственно). Необходимо также отметить структуры, подобные бактероидам клубеньковых бактерий и гетероцистам цианобактерий, служащие для защиты нитрогеназы от воздействия молекулярного кислорода.
Жгутики бактерий. Расположение жгутиков. Перитрихи. Монотрихи. Политрихи. Лофотрихи. Амфитрихи. Феномен роения. Диагностика подвижности бактерий.
По характеру движения подвижные бактерии разделяют на плавающие и скользящие(ползающие). Орган движения плавающих бактерий — жгутики; подвижность скользящих бактерий обеспечивают волнообразные сокращения тела.
Расположение жгутиков — характерный признак, имеющий таксономическое значение. Варианты расположения жгутиков приведены на рис. 4-1. У некоторых бактерий жгутики расположены по всей поверхности клеточной стенки (например, у бактерий рода Proteus), такие бактерии известны как перитрихи [от греч. peri-, вокруг, + trichos, волос]. Некоторые бактерии снабжены только одним толстым жгутиком (например, представители рода Vibrio), они известны как монотрихи. Политрихи — бактерии, имеющие одиночный по виду жгутик, образованный пучком из 2-50 жгутиков. Полярные жгутики прикреплены к одному или обоим концам бактерии. Монополярно-политрихиальное расположение жгутиков имеют лофотрихи [от греч. lophos, пучок, + trichos, волос], к ним, например, относят представителей рода Pseudomonas. Биполярно-политрихиальное жгутикование имеют амфитрихи [от греч. amphi-, двусторонний, + trichos, волос] (например, бактерии рода Spirillum).
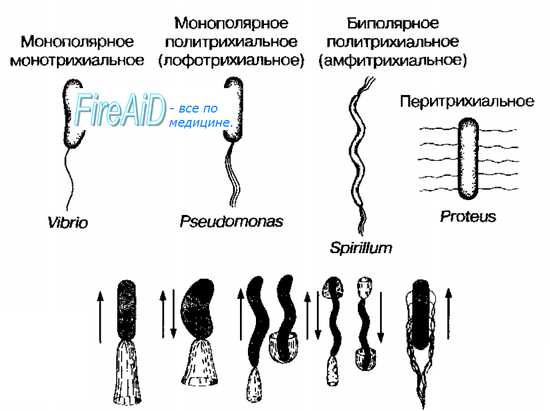
Рис. 4-1. Варианты расположения жгутиков (вверху) и движений бактерий (внизу).
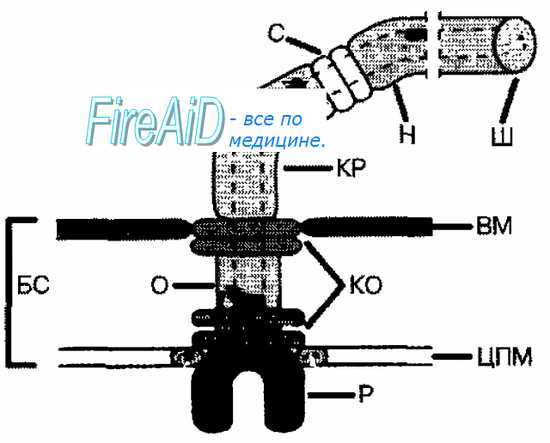
Рис. 4-2. Схема строения бактериального жгутика. БС — базальная структура, ВМ — внешняя мембрана, ЦПМ — цитоплазматическая мембрана, Р — ротор, О — ось, КО — кольцо жгутикового мотора, КР — крюк, С — цилиндрики-соединители, Н — нить жгутика, Ш — шапочка.
Способность бактерий к целенаправленному движению генетически обусловлена. Например, у Escherichia coli в регуляцию этого процесса вовлечено 3% генома (приблизительно 50 генов). Эти гены кодируют белки, образующие локомоторный аппарат, а также белки и ферменты, участвующие в преобразовании сигналов. Для жгутикового аппарата характерна периодическая изменчивость. Во многом этот процесс носит адаптивный характер и наиболее выражен у патогенных микроорганизмов. В частности, некоторые бактерии выработали систему вариабельности антигенных характеристик жгутиков, позволяющую им на какое-то время избегать направленных эффектов защитных иммунных механизмов.
Лабораторная диагностика подвижности бактерий
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Читайте также: