
Сообщение на тему нервная система позвоночных животных
Обновлено: 25.06.2024
Нервная система позвоночных
Нервная система позвоночных построена на принципах вероятностного развития, дублирования, избыточности и индивидуальной изменчивости. Это не означает, что в мозге позвоночных нет места генетической детерминации развития или нейрогормональной регуляции. Все эти слагаемые поведения присутствуют. Однако они играют несоизмеримо меньшую роль, чем у беспозвоночных. При развитии нервной системы позвоночных действуют законы формообразования, которые детерминируют первичную региональную экспрессию генов и морфогенез (Савельев, 2001). Одновременно есть и огромная переизбыточность эмбриональных нейробластов. Строгой детерминации развития каждого отдельного нейрона, как у беспозвоночных, нет. Судьба клетки вероятностна и зависит от тех коммуникативных взаимодействий, в которые она вступает во время индивидуального развития и дифференцировки. Достаточно высокая случайность судьбы каждого нейробласта отражается в огромных масштабах гибели клеток нервной системы. При развитии головного мозга млекопитающих нормальная гибель клеток обычно составляет 0,2–0,4 %. К концу метаморфоза бесхвостых амфибий гибнут 85 % всех нейробластов спинного мозга.
Такие потери клеток были бы невосполнимы для беспозвоночных, а у позвоночных эти события составляют нормальный морфогенез.
Вероятностное развитие нервных клеток позвоночных подтверждено в многочисленных экспериментах. Достаточно упомянуть способность нервной системы позвоночных к компенсации искусственно вызванной гибели клеток. Если у эмбриона амфибии на стадии нервной пластинки случайным образом (не в одном месте) разрушить 40 % клеток, то зародыш разовьётся в нормальный организм немного меньшего размера.
Рис. I-15. Гистологическое строение нервной системы позвоночных и беспозвоночных

Ганглии нервной системы имеют общий план строения как у позвоночных, так и у беспозвоночных животных. а — подглоточный ганглий речного рака; б — головные ганглии таракана; в — срез через головной мозг хорька; г — спинной мозг домовой мыши; д — ганглии дорсальных рогов спинного мозга мышонка. Срезы а, г, д окрашены по Маллори, б, в — по Нисслю.
Рис I-16. Основные тенденции усложнения структурной организации нервной системы.

Синими стрелками обозначен условный этап появления нервныхклеток, красными — основной путь церебрализации беспозвоночных, азелёными — позвоночных животных. В обоих случаях наиболее сложная нервная система организована по ганглиозно-ядерно-корковому типу.Основными отличиями позвоночных являются больший размер мозга,наличие мозговых желудочков и замкнутое кровообращение.
Многие костистые рыбы, обладающие электрорецепцией, имеют оченьразвитый задний мозг. Именно в нём и его деривате — мозжечке происходит анализ электрорецепторных сигналов из окружающего мира.
Это приводит к появлению стратифицированных структур ассоциативно- аналитического типа уже в заднем мозге. Таким образом, как у беспозвоночных, так и у позвоночных ассоциативные центры являются своеобразной надстройкой над анализаторами и могут располагаться в различных структурных отделах центральной нервной системы. Ум позвоночных нейрогормональные клетки выделены в специализированный
отдел — промежуточный мозг. Он столь же консервативен, как нейрогемальный орган беспозвоночных, и не меняет положения в мозге (Edinger, 1911; Nieuwenhuys, 1998).
Эти данные говорят не только о глубоких различиях, но и о сходстве развития и строения нервной системы позвоночных и беспозвоночных.
Реконструируя пути усложнения морфологической организации мозга, можно предположить такую последовательность событий. На первом этапе исторического развития нервной системы из клеток эктодермального зачатка появились чувствительные элементы (см. рис. I-16).
Специализированные клетки эктодермы обладали одновременно сенсорными и эффекторными функциями. Они рецептировали сигнал, проводили его к эффекторным органам и запускали их реакцию. Эти клетки были связаны между собой и формировали непрерывную сеть,которая не имела выраженных центров (см. рис. I- 12; I-14; I-16).
Такой тип организации нервной системы мы встречаем у кишечнополостных. При появлении более сложных поведенческих задач элементы нервной системы стали объединяться в небольшие скопления. По-видимому, это происходило двумя путями. С одной стороны, формировались параллельные ганглии (см. рис. I-16) с синаптическими контактами между телами клеток. Этот примитивный тип концентрации нервных элементов
отмечен у свободноплавающих кишечнополостных. С другой стороны, появились скопления нейронов с наружным расположением тел клеток и нейропилем из переплетённых отростков внутри ганглия (см. рис. I-16).
Этот тип организации ганглиев оказался достаточно эффективным и сохранился до настоящего времени у большинства беспозвоночных. Такой ганглий обладает рядом преимуществ, которые имеют особое значение для животных с незамкнутой кровеносной системой. Тела его нейронов расположены преимущественно на наружной поверхности, что позволяет поддерживать довольно высокий уровень метаболизма. Через открытую
поверхность тел нейронов происходит снабжение питательными веществами, кислородом и отводятся токсичные продукты жизнедеятельности клеток. Нейропиль, находящийся внутри ганглия, даёт возможность формировать синаптокомплексы, обмениваться сигналами и формировать генерализованный ответ на разнообразные воздействия. По- видимому, из этой формы концентрации нервных элементов возникли головные и туловищные ганглии высших беспозвоночных, ганглии и нервная трубка позвоночных (см. рис. I-15; I-16).
В головных ганглиях беспозвоночных сложились два основных типа гистологических структур: островковые скопления тел клеток и стратифицированные грибовидные тела. Островковые скопления тел клеток беспозвоночных практически идентичны подкорковым и стволовым ядрам позвоночных. Организация грибовидных тел напоминает слоистое расположение клеток в коре млекопитающих. Однако грибовидные тела
беспозвоночных не имеют упорядоченных вертикальных связей между нейронами. Тем не менее стратификация нейронов в грибовидных телах предполагает сходство механизмов обработки информации в ассоциативных центрах позвоночных и беспозвоночных животных.
Вероятно, нервная система позвоночных возникла из ганглиевбеспозвоночных с нейропилем из переплетённых отростков (см. рис. I-16).
Трубчатая нервная система сформировалась в результате выхода отростков нейронов из внутренней полости ганглия. Это событие привело к появлению нейропиля из отростков нервных клеток наружной стороны нервной трубки. Дальнейшее формирование новых нервных центров происходило преимущественно вокруг желудочков, в толще наружного переплетения отростков. В результате возникли центральное серое вещество и окружающие его волокна (белое вещество). Часть клеток выселялась из прижелудочковой зоны и формировала структуры ядерного или стратифицированного типа во внешнем нейропиле (см. рис. I-16). У высших позвоночных центральное серое вещество практически отсутствует, а основные нервные центры мозга представлены сложными ядрами и корковыми структурами различных типов (см. рис. I-15, в). В дальнейшем цефализация позвоночных развивалась по принципу количественного наращивания анатомического представительства анализаторных систем и ассоциативных центров. В отличие от беспозвоночных, нервная система трубчатого типа при замкнутой кровеносной системе может бесконечно увеличиваться в размерах. Это позволило позвоночным достигнуть очень высокого развития умственных способностей.
Подводя итог краткому обзору основных принципов анатомической и гистологической интеграции нервных клеток у беспозвоночных и позвоночных животных, необходимо сделать несколько общих выводов.
Во- первых, нейроны в обеих группах животных имеют сходное строение, но различаются по линейным размерам. Во-вторых, интегративные взаимодействия между нейронами осуществляются в сходных гистологических образованиях: ганглиях, ядрах и стратифицированных структурах. Эти образования встречаются как у беспозвоночных, так и у позвоночных животных.
Основным морфологическим отличием позвоночных являются размер нервной системы, наличие мозговых желудочков и организация кровеносной системы. Эффективная и замкнутая система кровоснабжения позволяет поддерживать высокий уровень метаболизма. Обширная система мозговых желудочков и сосудистое сплетение обеспечивают осмотический и водный баланс головного и спинного мозга. Эти морфологические
особенности позвоночных обеспечивают лабильность, большие адаптационные возможности и размеры нервной системы. Нервная система у беспозвоночных более консервативна, имеет небольшие размеры и не обладает столь широкими адаптивными возможностями, как у позвоночных. Зато она имеет огромный набор генетически детерминированных программ поведения, чётко запрограммированное развитие и эффективную систему гормональной регуляции физиологической активности. Небольшие размеры беспозвоночных позволяют добиваться впечатляющих успехов в конкуренции с другими видами животных. Сравнение организации позвоночных и беспозвоночных отражает различные принципы развития нервной системы, что даёт представление о возможных вариантах её строения и путях эволюции.
У всех позвоночных животных общий принцип построения нервной системы резко отличен от узлового типа построения нервной системы беспозвоночных животных. Центральная нервная система позвоночных животных ссамых ранних этапов эмбрионального развития не имеет узлового строения; она закладывается в виде сплошной нервной трубки, которая затем дифференцируется на различные отделы мозга и которая в дальнейшем является также источником образования нервных узлов на периферии (в симпатической и парасимпатической нервных системах). Такие узлы являются, следовательно, вторичными образованиями и состоят из клеток, мигрировавших из первоначальной сплошной нервной трубки. Нервная трубка всегда расположена на спинной (дорсальной) стороне тела, в то время как узловая нервная цепочка сложных беспозвоночных является брюшной и расположена на вентральной стороне.
У наиболее примитивных форм хордовых (ланцетник) центральная нервная система сохраняет характер мало дифференцированной сплошной нервной трубки и во взрослом состоянии. У всех же более сложно организованных форм происходит дифференциация последней. Она теряет свою равномерную форму, и на ее головном конце возникают утолщения нервной ткани в виде пузырей. Эти пузыри являются эмбриональным зачатком головного мозга.
Рассматривая различные классы позвоночных и сравнивая у них типы строения центральной нервной системы, довольно легко убедиться в том, что у всех образование мозговых пузырей происходит по одному и тому же типу; можно проследить эволюцию каждого пузыря, начиная от самых примитивных форм позвоночных и кончая млекопитающими. Варьирует лишь степень его развития и степень дифференциации в нем нервных элементов.
Основой образования головного мозга является деление переднего отдела мозговой трубки на 3 части, дающих начало трем мозговым пузырям, которые отчетливо выделяются у самых примитивных из существующих позвоночных — у круглоротых (следует учесть, что у современных круглоротых, особенно у миксин, есть черты деградации в связи с полупаразитическим образом жизни). По расположению эти эмбриональные пузыри можно обозначить как передний, средний и задний; соответственно у взрослого животного они образуют передний, средний и задний мозг.
У круглоротых задний и средний пузыри уже достаточно хорошо развиты и имеют весьма сложную нейронную структуру. Задний пузырь у миноги образует продолговатый мозг и, хотя и небольшой по объему, но хорошо видимый мозжечок, который имеет форму поперечного валика. Средний пузырь дифференцируется в весьма сложный средний мозг, который еще не разделяется на отдельные холмы, как у более высокоорганизованных животных. Он является основным центром обработки зрительных сигналов. Передний же мозговой пузырь сохраняет малодифференцированный характер. Из него образуются обонятельные луковицы и обонятельные доли, в которых заканчиваются пути от рецепторов обонятельной ямки; они тесно прижаты к слаборазвитому промежуточному мозгу.
Для рыб характерен примерно такой же общий план строения головного мозга — сложно организованный задний и средний мозг и мало дифференцированный (хотя и больший по размерам) передний мозг, выполняющий обонятельную функцию. В связи с тем, что движения у рыб значительно более сложны, и многим из них для существования необходимы точное поддержание положения тела в воде и тонкая двигательная координация, у них очень интенсивно развиваются те участки заднего мозга, которые связаны с высшей координацией двигательных актов, в частности — мозжечок. Мозжечок у многих рыб становится доминирующей частью центральной нервной системы (рис. 1.4).
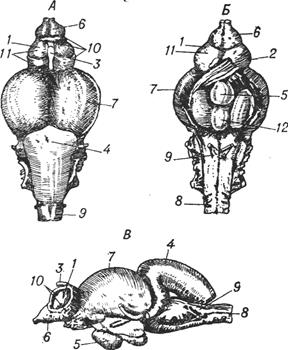
Рис. 1.4. Мозг форели. Вид сверху (А), снизу (Б) и сбоку (В);1 — базальный ганглий, 2 — хиазма, 3 — эпифиз, 4 — мозжечок, 5 — гипофиз, 6 — обонятельные доли, 7 — средний мозг, 8 — спинной мозг, 9 — задний мозг, 10 — плащ переднего мозга, 11 — передний мозг.
После выхода позвоночных на сушу происходит перераспределение функционального значения различных отделов головного мозга и соответственно изменение объема и сложности образований, возникающих из каждого мозгового пузыря. Это можно хорошо видеть, сравнивая мозг амфибий и рептилий с мозгом рыб. Задний мозг у первых занимает относительно меньший объем; мозжечок также развит значительно меньше, зато средний мозг развивается очень значительно, усложняясь в структурном отношении. Наряду с развивающимся в нем двухолмием, являющимся высшим зрительным центром, появляются дополнительные бугорки, в которые направляются пути от развившихся у этих животных органов слуха. Сравнительно простое строение мозжечка у амфибий может быть связано с упрощением этих животных, класс амфибий на данном этапе эволюционного развития находится на стадии деградации, ныне живущие земноводные отличаются весьма простыми, даже примитивными движениями.

Рис. 1.5. Мозг лягушки (по Р. Видерсгейму)- Вид сверху (А), снизу (Б) и сбоку (В); 1 — обонятельные доли, 2 — передний мозг, 3 — промежуточный мозг, 4 — эпифиз, 5 — зрительный тракт, 6 — средний мозг, 7 — мозжечок, 8 — задний мозг, 9 — воронка, 10 — гипофиз, 11 — спинной мозг.
И у амфибий, и у рептилий резко увеличивается передний мозг, который приобретает сложную анатомическую форму и не менее сложную внутреннюю структуру. Передний мозг дифференцируется на промежуточный мозг и два симметричных полушария, с которыми связаны обонятельные луковицы (рис. 1.5). Хотя передний мозг оказывается все еще осуществляющим одну из рецепторных функций, а именно — обоняние, он приобретает и более сложные координирующие функции, которые у рыб целиком связаны со средним мозгом.
С дальнейшим эволюционным прогрессом позвоночных животных связаны две различные линии развития мозговых пузырей.
Одна линия — это линия млекопитающих, у которых развитие нервной системы идет в сторону чрезвычайного увеличения и усложнения наружной части переднего мозгового пузыря, т. е. формирующей кору больших полушарий (рис. 1.6). Наряду с этим интенсивно развиваются и связанные с корой больших полушарий новые отделы мозжечка, который становится довольно объемистым, уступающим по массе нервной ткани и по сложности ее организации только структурам переднего мозгового пузыря. Остальные мозговые пузыри, в частности средний, явно отстают в развитии. Основные пути передачи зрительных сигналов, которые раньше направлялись в средний мозг, теперь через промежуточный мозг отклоняются в кору больших полушарий.
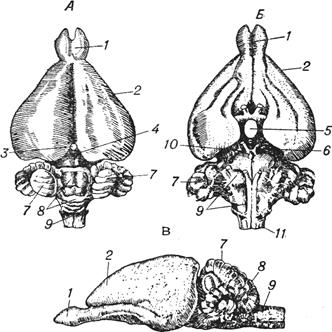
Рис. 1.6. Мозг кролика (по Р. Видерсгейму). Вид сверху (А), снизу (Б) и сбоку (В); 1 — обонятельные луковицы, 2 — передний мозг, 3 — эпифиз, 4 — средний мозг, 5 — гипофиз, 6 — хиазма, 7 — полушария мозжечка, 8 — червь мозжечка, 9 — задний мозг, 10 — ножки мозга, 11; — спинной мозг.
Вторая линия развития центральной нервной системы представлена головным мозгом птиц. Для этих животных характерно умеренное развитие из переднего мозгового пузыря коры больших полушарий; доминирующими по сложности и по объему остаются структуры, которые спрятаны в глубине полушарий (базальные ганглии и промежуточный мозг).
Функции нервной системы - восприятие внешних раздражении и передача возникающих возбуждений к клеткам, органам, тканям, а также объединение и согласование деятельности отдельных систем органов и организма в целом в единую функционирующую живую систему. Эмбрионально нервная система позвоночных возникает, так же как и у бесчерепных, в виде закладывающейся в эктодерме на спинной стороне зародыша полой трубки. В последующем происходит ее дифференцировка, приводящая . к образованию:
а) центральной нервной системы , представленной головным мозгом и спинным мозгом,
б) периферической нервной системы , состоящей из нервов; отходящих от головного и спинного мозга,
в) симпатической нервной системы , состоящей в основе из нервных узлов, расположенных около позвоночного столба и связанных продольными тяжами.
Головной мозг представлен у позвоночных животных пятью отделами: передним мозгом, промежуточным мозгом, средним мозгом, мозжечком и продолговатым мозгом. Он закладывается эмбрионально в виде вздутия переднего отдела нервной трубки, которое вскоре делится на три первичных мозговых пузыря. В дальнейшем первый мозговой пузырь дает начало спереди переднему мозгу; задняя его часть преобразуется в промежуточный мозг. Из второго мозгового пузыря формируется средний мозг. Путем выпячивания крыши третьего мозгового пузыря формируется мозжечок, под которым располагается продолговатый мозг. Передний мозг, кроме того, подразделяется на левую и правую половины. Одновременно с разрастанием и дифференцировкой головного отдела нервной трубки происходит соответственное преобразование невроцеля. Два его расширения в полушариях переднего мозга известны под названием боковых желудочков мозга. Расширенная часть невроцеля в промежуточном отделе мозга обозначается как третий желудочек, полость среднего мозга - как сильвиев водопровод, полость продолговатого мозга - как четвертый желудочек, или ромбовидная ямка. От головного мозга отходят 10 или 12 пар черепномозговых нервов. Передний мозг имеет впереди два симметрично расположенных выступа, от которых отходит первая пара головных нервов - обонятельные нервы (I). От дна промежуточного мозга отходят зрительные нервы (вторая пара головных нервов, II). На крыше промежуточного мозга развиваются два сидящих на ножках выступа: передний - теменной орган и задний - эпифиз. От дна промежуточного мозга отрастает непарный выступ - воронка, к которой прилегает сложное по строению и функции образование - гипофиз. Передний отдел гипофиза развивается из эпителия ротовой полости, задний - из мозгового вещества. Там же расположен гипоталамус. Крыша среднего мозга образует парные вздутия - зрительные доли (бугры). От среднего мозга отходит третья пара головных нервов (глазодвигательные нервы, III). Четвертая пара головных нервов (блоковые нервы, IV) отходит на границе между средним и продолговатым мозгом, все остальные головные нервы (V - Х - XII) отходят от продолговатого мозга. Спинной мозг не отграничен резко от продолговатого мозга. В центре спинного мозга (по главной оси органа) сохраняется невроцель, известный у позвоночных под названием спинномозгового канала. От спинного мозга метамерно (по числу сегментов) отходят спинномозговые нервы. Они начинаются двумя корешками: спинным - чувствующим и брюшным - двигательным. Эти корешки вскоре по выходе из спинного мозга сливаются, образуя спинномозговые нервы, которые затем вновь делятся на спинную и брюшную ветви.
Нейроэндокринная и иммунная система у всех позвоночных животных, от лососей до человека, построена по единому проекту. Несомненно, что личинка жемчужницы каким-то образом нейтрализует старческие изменения в регуляторной системе лосося "гипоталамус-гипофиз-периферические эндокринные железы-гипоталамус". Мы унаследовали от наших предков рыб принципы управления подкорковыми структурами. Если личинки жемчужницы научились отключать программу старения лосося, то почему бы и человеку не воспользоваться подобными механизмами, чтобы кардинально продлить свою жизнь.
Нервная система управляет всеми органами. Сокращение сердца, работа кишечника или мышц – все внутренние процессы находятся под строгим контролем нервной системы. Кроме того, работа нервной системы лежит в основе поведения и мышления животных.
Режим обучения доступен только авторизованным пользователям
Возможности режима обучения:
- просмотр истории в виде слайдов
- возможность прослушивания озвучки по каждому слайду
- возможность добавить свою, детскую озвучку
- тесты для детей, чтобы закрепить материал
- специально подобранные коллекции картинок и видео для улучшения восприятия
- ссылки на дополнительные обучающие курсы
Озвучка доступна в режиме обучения
Чтобы выжить, всем животным приходится решать одинаковые задачи: находить пищу, спасаться от хищника, воспроизводить и воспитывать потомство. Для успешного решения этих задач в ходе эволюции у животных сформировалось поведение. Например, хищники умеют незаметно подкрадываться к жертве, или расставлять ловушки, а жертва научилась замечать и убегать от опасности. За поведение животных отвечает нервная система. Благодаря нервной системе животное видит, чувствует, слышит и думает. Кроме того, нервная система управляет всеми органами. Сокращение сердца, работа кишечника или мышц – все внутренние процессы находятся под строгим контролем нервной системы.
Нервная система управляет животным
Как работает нервная система ?
Нервная система построена из нейронов
Нейроны в мозгу рыси общаются друг с другом
Количество нейронов у животных
Нервная система
Эволюция нервной системы
Губки и стрекающие
Впервые нервная система появляется у плоских червей. Черви уже способны осознанно перемещаться и прятаться. У них есть глаза, чувство равновесия и способность ощущать вкус. Так как основные органы чувств расположились на переднем конце тела, то здесь же начинают скапливаться нейроны. Вдоль всего тела протянуты нервные стволы. Количество нервных стволов у некоторых червей достигает 6 пар. Тем не менее, нервная система червей остаётся примитивной. Так, у круглого червя С. elegans известно расположение и происхождение всех 300 нейронов. У кольчатых червей нервные стволы слились в единственную брюшную нервную цепочку. В каждом сегменте имеется свое скопление нейронов – ганглий. Так, в теле пиявки насчитывается 32 сегмента, а значит и 32 ганглия.
Нервная система круглого червя
Членистоногие и моллюски
Нервная система позвоночных
Нервная система
На самых ранних стадиях развития мозг всех позвоночных похож на мозг ланцетника и делится на три отдела: Задний мозг. Управляет непроизвольными движениями, регулирует кровообращение и дыхание.
Средний мозг. Контролирует температуру тела, участвует в ориентации и контролирует внимание.
Передний мозг. Отвечает за сложное поведение, обучение и мышление.
Задний и средний мозг птиц и млекопитающих в целом похож на таковые у других позвоночных. Однако передний мозг претерпевает значительные изменения. Крупный, развитый передний мозг этих животных говорит о наличии продвинутого интеллекта.
Насколько сложно устроен мозг млекопитающих?
В основе поведения животных лежат рефлексы
Не следует путать инстинкты и рефлексы. Когда во время охоты кошка обучается новым приёмам – это приобретённый рефлекс. Но само желание охотиться на грызунов – это инстинкт. Инстинкты являются более сложным элементом поведения. Например, инстинкт самосохранения включает в себя потребность охотиться, защищаться от врагов и искать укрытие во время непогоды. Инстинкты являются врожденными и передаются по наследству. Какие ещё бывают инстинкты у животных? Только что вылупившиеся морские черепахи инстинктивно ползут к океану. После рождения сумчатое животное залезает в сумку матери. Пчёлы при помощи танца сообщают о местоположении нектара. Птицы инстинктивно вьют гнёзда, а белки запасают орехи на зиму.
Читайте также: