
Реферат на тему микроэволюция
Обновлено: 18.04.2024
Преодоление противоречий между эволюционной теорией и генетикой стало возможным с созданием синтетической теории эволюции, которая выступает основанием всей системы современной эволюционной биологии. Синтез генетики и эволюционного учения был качественным скачком в развитии как генетики, так и эволюционной теории. Он означал создание качественно нового ядра системы биологического познания, свидетельствовал о переходе биологии с классического на современный, неклассический уровень развития.
Оглавление
1.Введение…………………………………………………………. 3
2. Формирование синтетической теории эволюции…………………. 4
3. Микроэволюция – ядо СТЭ. Возникновение учения о микроэволюции…6
4. Популяция – элементарная единица эволюции……………………………7
5. Свойства и структура популяций…………………………………………..8
6. Список использованной литературы……………………………………. 15
Файлы: 1 файл
Реферат по биологии.doc
Учение о микроэволюции .
- Введение………………………………………………………… . 3
- Формирование синтетической теории эволюции…………………. 4
- Микроэволюция – ядо СТЭ. Возникновение учения о микроэволюции…6
- Популяция – элементарная единица эволюции……………………………7
- Свойства и структура популяций…………………………………………..8
- Список использованной литературы……………………………………. 15
К концу XIX в. теория Ч.Дарвина, объяснявшая эволюцию организмов воздействием естественного отбора, получила широкое признание. В этот период начались интенсивные исследования филогенеза различных групп организмов, которые привели к установлению ряда важных эмпирических закономерностей эволюционного процесса (законы необратимости эволюции, адаптивной радиации, биогенетический; принцип независимой эволюции отдельных признаков; проявления направленности эволюционного процесса, подобные параллельной эволюции близких филогенетических линий и др.). В то же время, при очевидном торжестве эволюционной идеи и всеобщем увлечении эволюционными исследованиями, многих ученых по различным причинам не удовлетворял классический дарвинизм. Неразработанными оставались проблемы наследственности. Первые эксперименты, показывавшие механизмы наследственности, были проделаны Г.Менделем еще в 1865 г., но его работы долго не привлекали внимания других исследователей. Лишь на рубеже XIX и XX вв. в изучении наследственности организмов были достигнуты первые существенные успехи .Преодоление противоречий между эволюционной теорией и генетикой стало возможным с созданием синтетической теории эволюции, которая выступает основанием всей системы современной эволюционной биологии. Синтез генетики и эволюционного учения был качественным скачком в развитии как генетики, так и эволюционной теории. Он означал создание качественно нового ядра системы биологического познания, свидетельствовал о переходе биологии с классического на современный, неклассический уровень развития.
2.Формирование синтетической теории эволюции.
В основе этой теории лежит представление о том, что элементарной “клеточкой” эволюции является не организм и не вид, а популяция. Именно популяция — та реальная целостная система взаимосвязи организмов, которая обладает всеми условиями для саморазвития, прежде всего способностью наследственного изменения в смене биологических поколений. Элементарной единицей наследственности выступает ген (участок молекулы дезоксирибонуклеиновой кислоты — ДНК, отвечающий за развитие определенных признаков организма). Наследственное изменение популяции в каком-либо определенном направлении осуществляется под воздействием ряда эволюционных факторов (изменяющих генотипический состав популяции): - мутационный процесс (поставляющий элементарный эволюционный материал), - популяционные волны (колебания численности популяции в ту или иную сторону от средней численности входящих в нее особей), - изоляция (закрепляющая различия в наборе генотипов и способствующая делению исходной популяции на несколько самостоятельных), - естественный отбор — процесс, определяющий вероятность достижения индивидами репродукционного возраста. Естественный отбор является ведущим эволюционным фактором, направляющим эволюционный процесс.
Формирование синтетической теории эволюции ознаменовало переход к популяционной концепции, сменившей организмоцентрическую, начало преодоления противопоставления исторического и структурно-инвариантного “срезов” в исследовании живого, интеграцию биологии на базе дарвинизма (в России — Н.И. Вавилов, И.И. Шмальгаузен, А.Н. Северцов, разработавший учение о главных направлениях биологического процесса — аромофозе и идиоадаптации, и др.). Это открыло качественно новый этап в развитии биологии — переход к созданию единой системы биологического знания, воспроизводящей законы развития и функционирования органического мира как целого.
Основные положения синтетической теории эволюции
в общих чертах можно выразить следующим образом:
3. Микроэволюция – ядро СТЭ. Возникновение учения о микроэволюции…
. Схема микроэволюции:
Элементарной, неделимой единицей микроэволюции является популяция.
Популяция — самая мелкая из групп особей, способная к эволюционному развитию. Популяция благодаря большой численности особей представляет собой непрерывный поток поколений и в силу мутационной изменчивости — разнородную (гетерогенную) смесь различных генотипов. Совокупность генотипов всех особей популяции — генофонд — основа микроэволюционных процессов в природе . Генетические характеристики популяции
Генофонды природных популяций отличает
наследственное разнообразие (генетическая гетерогенность, или полиморфизм), генетическое единство, динамическое равновесие доли особей с разными генотипами.
Наследственное разнообразие заключается в присутствии в генофонде одновременно различных аллелей отдельных генов. Первично оно создается мутационным процессом. Мутации, будучи обычно рецессивными и не влияя на фенотипы гетерозиготных организмов, сохраняются в генофондах популяций в скрытом от естественного отбора состоянии. Накапливаясь, они образуют резерв наследственной изменчивости. Благодаря комбинативной изменчивости этот резерв используется для создания в каждом поколении новых комбинаций аллелей. Объем такого резерва огромен. Так, при скрещивании организмов, различающихся по 1000 локусов, каждый из которых представлен десятью аллелями, количество вариантов генотипов достигает 10 1000 .
Генетическое единство популяции обусловливается достаточным уровнем панмиксии. В условиях случайного подбора скрещивающихся особей источником аллелей для генотипов организмов последовательных поколений является весь генофонд популяции. Генетическое единство проявляется также в общей генотипической изменчивости популяции при изменении условий существования, что обусловливает как выживание вида, так и образование новых видов.
Структура и свойства популяций
. Как первая надорганизменная биологическая система, популяция обладает определенной структурой и свойствами. Структуру популяции отражают такие ее показатели, как численность и распределение особей в пространстве, соотношение групп по полу и возрасту, их морфологические, поведенческие и другие особенности.
Численность — общее количество особей в популяции. Эта величина характеризуется широким диапазоном изменчивости, однако она не может быть ниже некоторых пределов. Сокращение численности по сравнению с этими пределами может привести к вымиранию популяции. Полагают, что если численность популяции меньше нескольких сотен особей, то любые случайные причины (пожар, наводнение, засуха, обильные снегопады, сильные морозы и т. д.) могут сократить ее настолько, что оставшиеся особи не смогут встречаться и оставить потомство. Рождаемость перестанет покрывать естественную убыль, и оставшиеся особи в течение сравнительно короткого времени вымрут.
Плотность — число особей на единицу площади или объема. При увеличении численности плотность популяции, как правило, возрастает; она остается прежней лишь в случае ее расселения и расширения ареала. У некоторых животных плотность популяции регулируется сложными поведенческими и физиологическими механизмами.
Пространственная структура популяции характеризуется особенностями размещения особей на занимаемой территории. Она определяется свойствами местообитания и биологическими особенностями вида. Наряду со случайным и равномерным распределением в природе наиболее часто встречается групповое распределение. Группа животных, прилагая совместные усилия, может легче защищаться от хищников, искать и добывать корм. Жизнь в семьях, стадах, колониях, гаремах приводит также к групповому распределению особей. Пространственная структура может изменяться во времени; она зависит от сезона года, от численности популяции, возрастной и половой структуры и т. д.
Половая структура отражает определенное соотношение мужских и женских особей в популяции. Генетический механизм определения пола обеспечивает расщепление потомства по полу в соотношении 1: 1. В силу разной жизнеспособности мужских и женских особей это первичное соотношение полов при оплодотворении часто заметно отличается от вторичного (при рождении — у млекопитающих) и тем более от третичного, характерного для половозрелых особей. Например, в популяциях человека вторичное соотношение полов составляет 100 девочек/106 мальчиков; к 16—18 годам это соотношение выравнивается и становится равным 1:1, к 50 годам— 100 женщин/85 мужчин, а к 80 годам соотношение по полу становится 2:1 (100 женщин/ 50 мужчин).

несколько новых. Такой процесс Дарвин называл дивергенцией или расхождением признаков.
Под этим термином он понимал возникновение разнообразных существ в потомстве от
одного предка. Из измененных форм выживают и дают потомство только наиболее
уклонившиеся разновидности, каждая из которых вновь дает веер изменившихся форм, и
опять выживают наиболее уклонившиеся и лучше приспособленные. Так, шаг за шагом
возникают все большие различия между крайними формами, перерастающими, наконец, в
различия между видами, семействами и так далее. В природе не всегда сохранялись лишь
наиболее расходящиеся, крайние формы, средние так же могли выжить и дать потомство. Из
крайних форм иногда развиваются одна, но может развиваться и более.
Если условия среды не изменяются или мало изменяются в течение длительного
времени, то вид остается почти неизмененным по сравнению с родоначальным.
С тридцатых годов текущего столетия внимание ученых привлекает популяция
как форма существования вида. Новые исследования проливают свет на самые начальные
этапы эволюционного процесса, которые протекают внутри вида и приводят к образованию
новых внутривидовых группировок – популяций и подвидов. Этот процесс называют
микроэволюцией. Она доступна непосредственному наблюдению и изучению, так как
Микроэволюция – эволюционный процесс, протекающий внутри вида, ведущий к
Сущность микроэволюционных преобразований составляет изменение генетической
структуры популяций. В результате действия элементарных эволюционных факторов
появляются новые аллели, а в результате действия отбора формируются новые адаптации.
При этом происходит замещение одного аллеля другим аллелем, одного изотопа белка
Популяции являются открытыми генетическими системами. Поэтому на
микроэволюционном уровне происходит латеральный перенос генов – обмен генетической
информацией между популяциями. Это означает, что адаптивный признак, возникший в
одной популяции, может перейти в другую популяцию. Следовательно, микроэволюцию
можно рассматривать как эволюцию открытых генетических систем, способных
В идеальных популяциях действует закон Харди-Вайнберга – закон генетического
равновесия, по которому соотношение частот встречаемости доминантного и рецессивного
генов остаётся неизменным из поколения в поколение.
Идеальная популяция отвечает следующим требованиям:
В популяциях частоты встречаемости генов А и а отвечают формуле p+q = 1, где p -
частота встречаемости гена А , q -частота встречаемости гена а . В идеальной популяции
частоты встречаемости генотипических комбинаций АА:Аа:аа остаются неизменными и
Однако в реальных популяциях условия идеальной популяции не выполняются. С.С.
Четвериков установил, что мутационный процесс в популяциях идёт постоянно, но мутации
в основном рецессивны и скрыты в гетерозиготах. При внешней фенотипической
однородности наблюдается генотипическая неоднородность популяции. С.С. Четвериков
сделал вывод о насыщенности мутациями природных популяций, что является скрытым
резервом наследственной изменчивости и приводит к нарушению генетического равновесия.
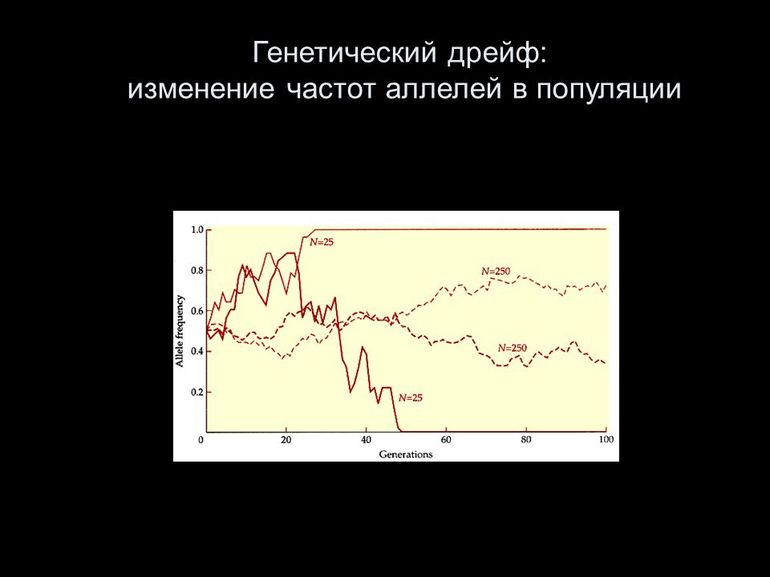
Случайное ненаправленное изменение частот аллелей в популяции получило название
дрейфа генов . Ещё в 1931 году С. Райтом была предложена концепция случайного дрейфа
генов, которая говорит об абсолютно случайном формировании генофонда дема как малой
выборки из генофонда всей популяции. Изначально дрейф генов оказался тем самым
аргументом, которого очень долго не хватало для того, чтобы объяснить происхождение
неадаптивных различий между таксонами. Поэтому идея дрейфа сразу стала близка
основал на дрейфе свою гипотезу квантовой эволюции, согласно которой популяция не
может самостоятельно выйти из зоны притяжения адаптивного пика. Поэтому, чтобы попасть
в неустойчивое промежуточное состояние, необходимо случайное, независящее от отбора
В природных условиях наблюдается периодическое колебание численности особей,
что связано с сезонными явлениями, климатическими изменениями, стихийными
бедствиями .Колебание численности особей в популяции называется популяционными
волнами . Их впервые обнаружил С.С. Четвериков. Популяционные волны – одна из причин
возрастание генетической однородности (гомозиготности) популяции;
сохранение аллелей, снижающих жизнеспособность особей;
Все эти изменения ведут к эволюционным преобразованиям генетической структуры
Всякое развитие начинается с изменения прежнего состояния объекта, поэтому в
основе процесса микроэволюции лежит изменчивость. Изменчивость генофонда популяции
может происходить в результате мутаций, их комбинаций (волны жизни), дрейфа генов, а
также переноса генетической информации из других популяций (поток генов).
Важную роль играет модификационная изменчивость , которая обеспечивает
существование особей данного вида в изменяющихся условиях среды и дает возможность
Изменение генетической структуры популяций происходит под влиянием факторов
среды. Поэтому предпосылки эволюции получили название генетико- экологических
факторов . В каждом новом поколении популяции появляется большее число потомков, чем
было родительских особей. В силу одинаковых потребностей в средствах существования
(пища, свет, влага, территория и т.п.) и в связи с ограниченностью средств существования для
удовлетворения этих потребностей между особями возникает борьба за существование,
Важным фактором эволюции является изоляция , которая приводит к расхождению
признаков одного вида и предотвращает скрещивание особей. Изоляция может быть
географической и экологической, отсюда выделяют два способа видообразования:
географическое, или аллопатическое, и экологическое, или симпатическое.
Географическое видообразование -процесс формирования и обособления
географических рас, когда популяции, дающие им начало, разделены механическими
преградами для скрещивания. Этот процесс идет очень медленно. Географическое
видообразование связанно с расширением ареала исходного вида или с расчленением ареала
на изолированные части физическими преградами (горы, реки, изменения климата). При
расширении ареала вида его популяции встречаются с новыми почвенно-климатическими
условиями, а также с новыми сообществами животных, растений и микроорганизмов. В
популяции постоянно возникают наследственные изменения, происходит борьба за
существование, действует естественный отбор. Все это со временем приводит к изменению
генного состава популяций. В дальнейшем эволюция популяции может привести к
Например, лиственница сибирская далеко продвинулась на восток; ее популяции
заселили территорию от Урала до Байкала, и оказалась в различных условиях. У особей
популяции постоянно возникали мутации, в результате скрещивания появлялись новые
комбинации генов; благодаря этим процессам популяция становилась неоднородной. В
процессе борьбы за существование и в результате действия естественного отбора выживали и
оставляли потомство особи с полезными в конкурентных условиях обитания изменениями.
Действия этих факторов на протяжении длительного времени способствовало
появлению более резких различий между популяциями и в конечном итоге возникновению
биологической изоляции – нескрещиваемости особей разных популяций одного вида .
В результате в более суровых условиях под действием движущих сил эволюции
сформировался новый вид – лиственница наурская. В условиях Крайнего Севера подобным
образом образовался особый вид мака с небольшими, сильно опущенными листьями,
быстрым развитием коротких цветоносов и ранним цветением.
Еще пример: у лесного ландыша сначала был сплошной ареал, но с оледенением он
распался на изолированные части; на этих территориях сформировались самостоятельные
популяции, признаваемые некоторыми учеными за молодые виды.
Повсеместно распространенный в Европе прострел занимает непрерывный ареал с
запада на восток. У западной формы листья тонко рассечены и разбросаны, цветы поникшие,
благодаря чему при обилии дождей на западе вода разбрызгивается и не застаивается, а
пыльца не смывается. Восточная форма в засушливых условиях характерна более грубо
расчлененными, стоячими листьями и стоячими цветками, по которым вода стекает к корням.
Опытами доказано, что листья восточной формы испаряют воду значительно меньше, чем
листья западной. Все эти отличия прострела на западе Европы от прострела на востоке носят
приспособительный характер. Между западной и восточной формами прострела существует
непрерывный ряд переходных форм. В средней полосе нашей страны произрастает белее
двадцати видов лютика. Все они произошли от одного вида. Потомки его заселили различные
места обитания – степи, леса, поля – и благодаря изоляции обособились друг от друга сначала
Вид синица большая представляет сложный комплекс популяций и подвидов,
находящихся на разных ступенях изоляции. Ареалы трех подвидов синицы большой –
евроазиатского, южно-азиатского и восточноазиатского – образуют кольцо вокруг
Подвиды занимают хорошо очерченные ареалы, но в зонах контакта южно-азиатские
синицы скрещиваются с двумя другими подвидами. Восточноазиатские и евроазиатские,
обитая совместно в долинах верхнего Амура, не скрещиваются. Обособление ареалов синиц и
образование их подвидов связано с наступлением ледника. В озере Байкал живут многие
виды и роды плоских ресничных червей, ракообразных и рыб, больше нигде не
встречающихся, так как озеро отделено от других водных бассейнов горными хребтами уже
Экологическое видообразование -способ видообразования, при котором новые
формы занимают разные экологические ниши в пределах одного ареала. Экологическое
видообразование происходит в тех случаях, когда популяции одного вида остаются в
пределах своего ареала, но условия обитания у них оказываются различными. Под влиянием
движущих сил эволюции изменяется их генный состав. Через множество поколений эти
изменения могут зайти так далеко, что особи разных популяций одного вида не будут
скрещиваться между собой, возникает биологическая изоляция, что характерно, как правило,
для разных видов. Экологическое видообразование наблюдается в пределах ареала
Например, один вид традесканции сформировался на солнечных скалистых вершинах,
а другой – в тенистых лесах. В пойме нижней Волги образовались виды житняка, костра,
щетинника (мышея), которые дают семена до разлива реки или после него. Этим они
обособлены от исходных видов, растущих на незаливаемых местах и осеменяющихся по
Естественным отбором, связанным с хозяйственной деятельностью человека, созданы
подвиды, изолированные по срокам цветения. Некоторые ученые считают их видами.
Симпатически происходит видообразование у некоторых насекомоопыляемых
растений, на основе специализации насекомых- опылителей цветка какой-либо группы
внутри данного вида. Избирательная специализация насекомых-опылителей приводит к
половой изоляции данной группы растений от находящихся рядом растений, давших начало
Образование симпатических групп возможно и на основе случайно отдаленной
гибридизации, если получающиеся гибриды оказываются плодовитыми, то, как правило, они
не скрещиваются с родительскими формами (например, из-за изменения числа хромосом).
Примером такого рода видообразования может служить рябинокизильник,
являющийся гибридом рябины и кизильником. Пять видов синиц образовались в связи с
пищевой специализацией: синица большая питается крупными насекомыми в садах, парках;
лазоревка добывает мелких насекомых в щелях коры, в почках; хохлатая синица питается
семенами хвойных деревьев; гаичка и московка питаются преимущественно насекомыми в
лесах разных типов. Популяции севанской форели различаются по срокам нереста, местам и
глубине нерестилищ (озеро, река). Видообразование продолжается и в настоящее время. Вид
черный дрозд в настоящее время распадается на две группы, еще не различимые внешне.
Одна из них обитает в глухих лесах, другая держится близ жилья человека. Это можно
Географическое и экологическое видообразование идёт по схеме: обособление
популяции накопление мутаций изоляция дивергенция признаков образование
подвида репродуктивная изоляция образование в ида.
Популяции и подвиды иногда не различимы. На разных этапах микроэволюции один
способ сменяет другой или они действуют совместно. Географическая изоляция может в
дальнейшем присовокупить действия экологической, поэтому трудно установить границы
каждого способа видообразования. Образованием нового вида завершается микроэволюция.

ГОСТ
Микроэволюция – это система эволюционных изменений, характерных для отдельных популяций и вида вплоть до образования нового вида.
Определение понятия "микроэволюция"
Следует отметить тот факт, что приведенные определения термина микроэволюция не являются исчерпывающими и могут быть дополнены в зависимости от подхода к его трактовке.
Элементарное эволюционное изменение – это необратимое, но направленное генотипическое изменение, которое касается соотношения генов в популяционном генофонде и расширяет эволюционные возможности популяции.
Таким образом, элементарные эволюционные изменения служат материалом для формирования новых видов в природе.
С течением времени генофонд внутри популяций изменяется настолько, что скрещивание между особей перестает быть возможным. Если популяции остаются относительно изолированными друг от друга, то происходит образование новых видов и наступает завершающий этап микроэволюции.
В науке различаются:
- филетическая микроэволюция;
- видообразование.
Что касается филетической микроэволюции, то ее определяют, как комплекс постепенных изменений в течение времени в пределах одного вида или группы особей. Такой процесс приводит к возрастанию степени приспособленности организмов к конкретным условиям среды обитания.
Готовые работы на аналогичную тему
Видообразование, в свою, очередь является общей системой образования нескольких видов из одного.
Факторы микроэволюционного процесса
Для того, чтобы понять, каким образом, происходит процесс микроэволюции, необходимо охарактеризовать ее факторы. К ним относят:
- мутации, дрейф генов, популяционные волны, которые поставляют изменчивость в популяционный генофонд;
- изоляция представляет собой возникающие препятствия, которые существенно уменьшают возможность размножения и обмена генетической информацией между популяциями внутри вида;
- естественный отбор позволяет закрепить те особенности, которые повышают приспособленность особей внутри популяции к различным изменениям внешней среды.
Мутации имеют ненаправленный характер. Они могут быть генными и хромосомными, геномными. К генным мутациям относят замену одного основания другим внутри триплета и др. Хромосомные мутации выражены в увеличении и уменьшении численности хромосом, а также в повреждении какого – либо участка хромосом. Такие мутации очень часто приводят к гибели организма. Любая мутация может быть вызвана целым рядом факторов, которые именуются мутагенами.
Генетические потоки представляют собой перенос мутаций из других популяций. Этот процесс зависит от широты расселения особей из соседних популяций внутри вида. Мутации наряду с генетическими потоками дают разнообразие генотипов внутри популяции. Естественный отбор, а также дрейф генов позволяют сортировать генотипы.
Сами по себе мутации без воздействия естественного отбора не могут привести к появлению новых видов. Популяционные волны отражают флуктуацию численности популяции. В этом смысле они также являются поставщиком эволюционного материала и подставляют редкие мутации под действие отбора. Другими словами, мутация и естественный отбор в комплексе может стать эволюционным способом для разграничения полезных и вредных адаптаций внутри экосистем и при анализе взаимодействия популяций между собой и внутри вида.
Изоляция является фактором накопления и закрепления изменений, которые вызываются всеми вышеописанными факторами. Изоляция бывает территориальной и биологической. Территориальная изоляция представлена разобщённостью популяции внутри конкретного ареала. Репродуктивная изоляция проявляется в несовпадении сроков размножения отдельных групп особей внутри вида.
Биологическая изоляция говорит о физической несовместимости особей и невозможности воспроизведения полноценного потомства.
Естественный отбор является вектором эволюционного процесса. Он может быть направленным и ненаправленным. Также выделяют следующие формы естественного отбора:
- движущий или смещающий значение признака от средней величины и позволяющий особям приспособиться к новым условиям обитания;
- стабилизирующий отбор проявляется в стабильных популяциях и выбраковывает из них крайние генотипы. Другими словами, при действии данной формы отбора жизнеспособность сохраняют те особи, которые имеют значение признака близкое к среднему.
- дизруптивный отбор оставляет внутри популяций особи с редкими генотипами. Иначе говоря, дизруптивный отбор является способом выявить крайние отклонения значения признаков от среднего значения.
Наконец, дрейф генов представляет собой случайное отклонение от постоянной частоты встречаемости генов. Дрейф также закрепляет редкие генотипы в популяциях.
Все вышеописанные факторы приводят к изменению генофонда популяции. Если на совокупность данных процессов накладывается изоляция, то происходит формирование новых видов или более мелких таксономических единиц.
Подводя итог всему вышесказанному, можно сделать вывод о том, что направление эволюции проходит по пути образования одного нового гена за другим при приспособлении к изменениям окружающей среды и образованию нового вида или новой экосистемы. Любое эволюционное изменение, относящееся к системе микроэволюции, так как базируется на случайных эволюционных изменениях в генофонде популяций и генетическом аппарате отдельных особей.
Микроэволюция – это процесс перестройки генофонда популяций под влиянием внешних факторов, в результате чего появляются новые виды.
Популяция и ее признаки
Популяция – это совокупность представителей одного вида, которые населяют ограниченную территорию, могут свободно скрещиваться и обмениваться генетической информацией.
Популяция – наименьшая элементарная единица эволюции, отдельный организм нельзя рассматривать как единицу эволюции, потому что его геном не изменяется с течением времени.

Виды популяций
Существует три вида популяций:
Элементарная или микропопуляция – это группа организмов, населяющих небольшую территорию. Представители имеют сходный генотип и часто скрещиваются.
Экологическая популяция – это популяции особей, которые находятся в одном ареале, тесно взаимодействуют, но сохраняют свои отличительные черты.
Географическая популяция – совокупность организмов, заселяющих относительно большую территорию с однородными географическими условиями. Такие популяции живут изолированно и редко скрещиваются со своими сородичами из соседних популяций.
Основные характеристики популяций
Плотность популяции – число особей, населяющих определенную площадь.
Численность популяции – это совокупность всех представителей популяции на изучаемой территории.
Рождаемость и смертность – указывают на способность популяции к репродукции и выживанию, и позволяет спрогнозировать конечный исход.
Формы микроэволюции
Филатическая – постепенное изменение строения и внешних признаков особей одного вида, популяции. При этом адаптационные возможности организмов возрастают.
Видообразование – заключается в эволюции отдельных популяций одного вида. В итоге исходный вид распадается на новые, или дает ответвление.
Способы видообразования
Микроэволюционные явления возможны при сочетании ряда факторов. Основной из них — изоляция, необходимая для расхождения характеристик вида и предотвращения обмена генетической информацией между его представителями. Выходя из этого, выделяют 2 способа образования вида: географическое и экологическое.
Географическое (аллопатическое) видообразование
Географическое (аллопатическое) видообразование – обусловлено наличием преград между популяциями одного вида, что не дает им свободно скрещиваться. Данный процесс довольно длительный. Аллопатическое видообразование связано с распространением живых организмов за пределы обычного ареала, при этом изоляция наступает через географические преграды – горные массивы, моря или перемены климатических зон.
Попадая в другие условия, меняется окружающий популяцию растительный и животный мир, температура окружающей среды и др., чтобы выжить необходимо преодолеть ряд адаптационных изменений. Новые поколения рождаются уже с наследственными мутациями, продолжается естественный отбор, где выживают сильнейшие. Таким образом, постепенно геном популяции меняется, что со временем может привести к формированию нового вида.
Экологическое (симпатическое) видообразование
Экологическое (симпатическое) видообразование предполагает образование видов путем изоляции особей в пределах одного ареала.
Представители вида живут в разных экологических нишах, перестают конкурировать между собой, и начинают развитие по своей линии. После генетических модификаций последующие поколения будут находиться уже в биологической изоляции, не скрещиваясь друг с другом – это последний этап формирования нового вида.
Видообразование как результат микроэволюции идет по установленной схеме:
- Выделение популяции;
- мутации и рекомбинации генома;
- изолирование;
- расхождение признаков;
- формирование подвидов;
- биологическая изоляция;
- появление нового вида.
К примеру, в начале прошлого столетия часть зайцев-русаков вывезли из обычного места обитания и поселили на юго-западной территории Сибири. От сородичей их отделяли непреодолимые горы, безграничная степная зона, что привело к полной изоляции популяции зайцев в Сибири. Спустя 20 лет сибирских зайцев пришлось выделить в отдельный вид, так как произошли значительные изменения на генетическом уровне.
Микроэволюция приводит к внутривидовым изменениям, которые возможны благодаря действию мутаций, естественного отбора, дрейфа генов. Данные факторы лежат в основе и макроэволюционных процессов, только к микроэволюции относится образование видов, а макроэволюция направлена на формирование крупных таксонов – родов, семейств и т.д.
Значение мутаций для микроэволюции
Исходным материалом для микроэволюции служат мутации. Они возникают часто, но не все имеют направленный характер и существенно не влияют на генофонд. Мутации, которые ведут к стойкому изменению генотипа, происходят реже и действуют на уровне генома или отдельных хромосом.
Мутационная изменчивость – основа для наследственных изменений, первичное звено всех дальнейших преобразований. Так, в процессе микроэволюции образуются новые виды под действием мутагенных факторов.
Естественный отбор
Естественный отбор – основная движущая сила микроэволюции. При этом в популяциях увеличивается численность организмов способных к быстрой адаптации и выживаемости, а особи с неблагоприятными характеристиками погибают.
Так, путем естественного отбора популяция избавляется от вредоносных мутаций, а полезные качества кодируются доминантными аллелями и передаются следующим поколениям. Но одна и та же мутация может быть полезной и вредной, все зависит от окружающих условий.
Например, при появлении бескрылых насекомых в обычной среде, их быстро бы вытеснили крылатые сородичи, но на островах с сильными, постоянными ветрами именно бескрылые смогли выжить и занять доминирующее положение.
Дрейф генов
Помимо мутаций в популяциях возможен дрейф генов – случайные изменения генома в небольших группах. Дрейф генов не имеет большого значения, поскольку приводит к бесцельным эволюционным сдвигам.
Проявление дрейфа генов наблюдается в родственных видах, которые населяют разные зоны земного шара. При этом представители будут иметь незначительные отличительные черты, которые не имеют существенного влияния.
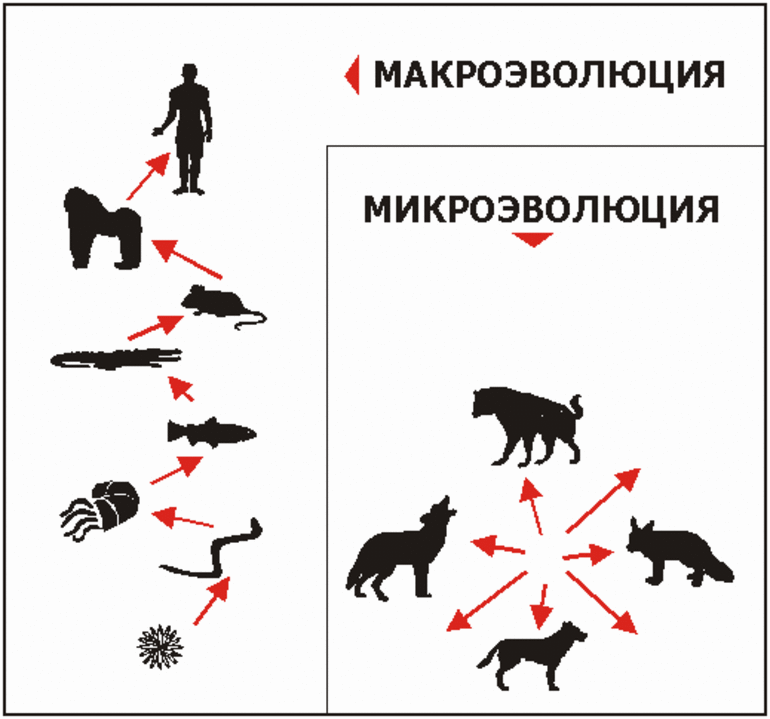
История и определение
Эволюция является результатом изменений в генетическом коде. Гены кодируют основные характеристики, которые будет иметь жизненная форма, и нет никакого известного механизма, предотвращающего небольшие изменения (микроэволюция), приводящие в итоге к макроэволюции.

Дарвин, с другой стороны, не видел принципиальных различий между микроэволюцией и макроэволюцией и считал, что они являются одним направлением биологического прогресса. Но он подтвердил, что безусловно, чёткая линия разграничения между ними не проведена.
С открытием ДНК и генов генетическая мутация получила признание как механизм вариации. Эта теория развивающейся эволюции была позже названа современным эволюционным синтезом и до сих пор сохраняет своё выдающееся положение. Синтетическая модель эволюции соответствовала микроэволюции и макроэволюции, утверждая, что единственной разницей между ними были время и масштаб.
Сегодня эти термины остаются в относительно ограниченном использовании. Их можно найти в некоторых научных текстах, но в целом для большинства биологов они не имеют значения. Для учёных нет существенной разницы между микроэволюцией и макроэволюцией. И то и другое происходит одинаково и по одним и тем же причинам, поэтому не нужно их дифференцировать.
Проблема, связанная с применением терминов микроэволюция и макроэволюция, заключается в том, что определение того, что составляет вид, не всегда устанавливается. Это может усложнить границы, которые, как утверждают некоторые, существуют между ними.
Сходство и различия
Оба процесса происходят в природе по одной схеме: наследственность, изменчивость, естественный отбор и репродуктивная изоляция. Но в то время как микроэволюция объясняет диверсификацию на индивидуальном уровне в течение относительно коротких периодов, макроэволюция определяет модифицирование в больших популяциях, часто влекущие за собой катастрофические изменения окружающей среды. Эти процессы завершаются образованием новых видов.
Макроэволяция является результатом микроэволюции — самого первого этапа. Она относится к модификации генофонда популяции с течением времени, что приводит к небольшим изменениям в организме одного и того же вида. С другой стороны, макроэволюция относится к коренному модифицированию организмов, эти изменения постепенно приводят к тому, что могут образоваться совершенно новые типы, классы, и виды организмов. То есть происходит надвидовая эволюция.
Хотя различия наблюдаются во всех формах жизни, будь то вирус, растение, бактерии, животные или люди, эта диверсификация является единственным фактором, который отличает каждого друг от друга.
Таблица сравнения микро и макроэволюции
| Признаки | Микроэволюция | Макроэволюция | |
| Сходство | |||
| Факторы эволюции по Дарвину | Изменчивость. Борьба за выживание. | ||
| Главная движущая сила эволюции | Естественный отбор | ||
| Последствия | Многообразие видов, приспособленность организмов | ||
| Различия | |||
| Процесс по отношению к виду | Внутривидовой | Надвидовой | |
| Пределы эволюции | В пределах вида | В пределах рода и выше | |
| Возникновением какой новой группы начинается этот процесс? | Популяции | Группы видов одного рода | |
| Возникновением какой новой систематической группы организмов завершается этот процесс? | Вида | Класса, типа | |
| Начало и конец процесса | Микроэволюция | Макроэволюция | |
| Доступность для наблюдения | Да | Нет | |
Критерии и структура
Микроэволюция вида — это изменение в геноме или генофонде за относительно короткий период геологического времени за счёт трансформаций в успешно воспроизводимых особях в популяции. Причиной изменений являются:
- мутация;
- генетический дрейф;
- поток генов;
- естественный отбор.
Мутации считаются одной из наиболее вероятных причин вариаций, что приводит к появлению новых аллелей. Они возникают из-за ошибок репликации, ультрафиолетового излучения, вирусов и мутагенных химических веществ. Естественный отбор занимает тысячи лет и приносит заметные изменения. Есть многочисленные случаи естественного отбора, такие как домовые воробьи, которые были введены в 1852 году в Северную Америку. С того времени они развивались с различными характеристиками, которые зависили от места их обитания. Другим примером может служить устойчивость микроорганизмов к антибиотикам.
Миграция генов — это их передача посредством физических движений аллелей в популяции, что означает — поток генов происходит, когда какие-либо особи мигрируют между популяциями.
Естественный отбор рассматривается как крупномасштабные изменения, которые наблюдаются в организме, но эти изменения занимают тысячи лет. Этот термин также следует концепции универсального общего происхождения всех живых организмов.
Современный синтез
В рамках современного синтеза начала XX века макроэволюция рассматривается как совокупный эффект микроэволюции. Таким образом, различие между микро- и макроэволюцией не является принципиальным, единственное различие между ними заключается во времени и масштабе.

Однако время не является обязательным отличительным фактором, макроэволюция может происходить без постепенного объединения небольших изменений. Дублирование всего генома приводит к видообразованию в течение одного поколения, это особенно часто встречается у растений.
Изменения в генах, регулирующих развитие, также важны для видообразования через большие и относительно внезапные изменения в морфологии животных. Есть много способов проследить макроэволюцию, наблюдая за изменениями в:
- генетике;
- морфологии;
- таксономии;
- экологии и поведении организмов.
Типы изменений
По мере увеличения таксономического разнообразия у четвероногих появились стимулы для перехода на новый образ жизни, где первоначально ресурсы могут казаться неограниченными, есть немного конкурентов и возможное убежище от опасности. И по мере увеличения экологического разнообразия таксоны диверсифицируют от их предков в гораздо большей степени среди фауны с более превосходной, инновационной или более гибкой адаптацией.
Молекулярная эволюция происходит через небольшие изменения на молекулярном или клеточном уровне. В течение длительного периода времени это может оказать большое влияние на генетику организмов. Таксономический отбор происходит за счёт небольших изменений между популяциями, а затем и видами.
Морфологическая эволюция видна через небольшие изменения в морфологии организма. Это хорошо заметно у китообразных, у которых на протяжении всей ранней эволюции группы задние конечности всё ещё присутствовали.
Однако в течение миллионов лет они регрессировали и становились внутренними.
Резкие превращения из одной биологической системы в другую, например, переход жизни из воды на землю или переход от беспозвоночных к позвоночным, редки. В течение эволюционной истории жизни появилось только несколько основных биологических типов. Когда жизненные формы совершают такие гигантские скачки, они практически не встречают конкурентов и могут использовать множество доступных ниш после адаптивного излучения. Это может привести к конвергентной эволюции, поскольку пустые ниши заполнены тем, что встречает их жизненная форма. Предметы, изучаемые в рамках макроэволюции, включают:

- Адаптивные излучения, такие как кембрийский взрыв.
- Изменения в биоразнообразии во времени.
- Эволюция генома, такая как горизонтальный перенос генов, слияния генома при эндосимбиозах и адаптивные изменения размера генома.
- Массовые вымирания.
- Оценка темпов диверсификации, включая темпы видообразования и вымирания.
- Спор между пунктуальным равновесием и постепенностью.
- Роль развития в формировании эволюции, особенно таких тем, как гетерохронность и фенотипическая пластичность.
Доказательства процесса
Палеонтология, эволюционная биология развития, сравнительная геномика и геномная биостратиграфия вносят бо́льшую часть доказательств в паттерны и процессы макроэволюции. Один из примеров эмбриологических доказательств макроэволюции — присутствие одних и тех же структур у зародышей рептилий и млекопитающих.
Найденные окаменелости устанавливают наследственную линию как растений, так и животных, и определяют периоды пунктуального равновесия в обоих. Скальные пласты можно использовать для датировки окаменелостей, потому что организмы, из которых были получены ископаемые, погибли и в конечном счёте были погребены в породе. Это делает возможным относительное датирование окаменелостей, определяя их возраст по сравнению с другими слоями породы.
Самые молодые горные породы и содержащиеся в них окаменелости находятся ближе к поверхности земли и стареют по мере того, как глубже они обнаруживаются в коре. Кроме того, слои горных пород в соседних областях могут вести себя на протяжении многих лет одинаково, если они состоят из такого же материала или минерала.

Эволюционная история области выстроена в слоях горных пород, которые вместе создают мозаику, позволяющую проследить макроэволюцию или основные события в истории жизни на Земле. Поскольку палеонтологи обнаруживают окаменелости в слое породы, они могут делать предположения на основе современных форм жизни об условиях окружающей среды, которые существовали в то время. Например, обнаружение окаменелости папоротника укажет на тёплый или умеренный климат, достаточное количество осадков, и всё это — условия, которые поддерживают рост современных папоротников.
Это может также предоставить подсказки, которые связаны с другими ископаемыми свидетельствами, чтобы пролить свет на животную жизнь, существующую в то время. Например, данная среда, которая поддерживает папоротники, также, вероятно, будет поддерживать травоядных животных, таких как улитки или пасущиеся животные, питающиеся папоротниками.
Читайте также: