
Реферат на тему цитоплазматическая наследственность
Обновлено: 06.07.2024
Наследственность — это уникальная способность живых организмов к передаче собственных признаков каждому последующему поколению.
В какой-то момент люди стали отмечать, что в процессе размножения растения и животные в своем потомстве воспроизводят определенные качества. К тому же, еще до четкого понимания механизмов передачи наследственной информации люди активно практиковали селекцию. Человек научился получать растения разных сортов и породы животных с определенными качествами и признаками.
Развитие микробиологии дало понимание, что нуклеиновые кислоты являются хранителями и переносчиками наследственной информации.
Есть два вида наследственности:
- Ядерная наследственность. Она представляет собой совокупность наследственной информации, которая содержится в нуклеиновых кислотах ядра (а именно в хромосомах). Также такая наследственность называется хромосомной.
- Внеядерная или цитоплазматическая наследственность. Под ней понимают способность определенных структур цитоплазмы хранить, а затем передавать следующим поколениям наследственную информацию.
Виды наследственности
Цитоплазматическая наследственность
В основе внеядерной или цитоплазматической наследственности лежит способность определенных структурных компонентов цитоплазмы к сохранению и передаче от родительского организма дочернему часть наследственной информации.
В процессе наследования главная роль отводится генам хромосом — именно они обеспечивают наследование большей части признаков организма. Тем не менее внеядерная наследственность тоже важна.
С цитоплазматической наследственностью связаны два генетических явления:
- с наследованием признаков, которые обусловлены внеядерными генами, находящимися в специальных органеллах. Роль таких органелл отводится пластидам и митохондриям;
- с возникновением у потомков признаков, обусловленных ядерными генами родительских форм. На их развитие и проявление оказывает влияние цитоплазма яйцеклетки.
В митохондриях и пластидах, под которыми понимают способные к самоудвоению полуавтономные органеллы, в начале 20 века ученые обнаружили наличие генов.
Пластидное наследование
Связанная с генами пластид цитоплазматическая наследственность встречается у многих цветочных растений (львиный зев, ночная красавица и др). Некоторые из них — формы с довольно пестрыми листьями. Такой признак передается только по материнской линии.
Пестрые листья — результат того, что отдельные части пластид не могут образовывать хлорофилл. Так же в ходе образования гамет (спермиев и яйцеклеток) пластиды попадают к яйцеклеткам, а не к спермиям.
Пластиды, которые размножаются делением, характеризуются генетической непрерывностью:
- зеленые пластиды дают начало зеленым;
- бесцветные пластиды образуют бесцветные.
В процессе деления клетки разного типа пластиды распределяются случайным образом. Так происходит образование клеток с бесцветными, зелеными или пластидами обоих вариантов одновременно.
Митохондриальная наследственность
Связанную с митохондриями цитоплазматическую наследственность изучали на примере дрожжей. Их митохондрии содержат гены, влияющие на наличие или отсутствие дыхательных ферментов. Помимо этого, эти гены определяют степень устойчивость к определенным антибиотикам.
Есть способ, который помогает проследить, как влияют ядерные гены материнского организма через цитоплазму яйцеклетки на формирование некоторых состояний отдельных признаков потомков. Для этого используется пресноводный брюхоногий моллюск-прудовик.
Для него характерно наличие форм с разными состояниями наследственного признака: раковина этого моллюска может быть закручена как влево, так и вправо.
Аллель, обеспечивающая закрученность ракушки вправо, доминирует над аллелью, гарантирующей левозакрученность. При этом само направление закрученности определяют исключительно гены материнского организма.
Правозакрученную ракушку, к примеру, могут иметь гомозиготные особи по рецессивному признаку левозакрученности. Но только тогда, когда происходят от материнского организма с доминантной аллелью правозакрученности.

ГОСТ
Наследственность – это способность живых организмов передавать свои признаки каждому последующему поколению.
Люди давно стали замечать, что растения и животные при размножении воспроизводят в потомстве определенные свои качества. Задолго до выяснения механизма передачи наследственной информации человечество уже занималось селекцией. Люди научились получать растения разных сортов и породы животных, имеющие требуемые качества. С развитием микробиологии ученым удалось выяснить, что роль хранителей и переносчиков наследственной информации выполняют нуклеиновые кислоты. В настоящее время выделяют ядерную и внеядерную (цитоплазматическую) наследственность.
Ядерная наследственность – это совокупность наследственной информации, содержащаяся в нуклеиновых кислотах ядра (в хромосомах).
Ее еще называют хромосомной.
Внеядерная (цитоплазматическая) наследственность – это способность определенных структур цитоплазмы хранить и в дальнейшем передавать из поколения в поколение часть наследственной информации.
Цитоплазматическая наследственность
Явление внеядерной или цитоплазматической наследственности состоит непосредственно в способности некоторых структурных компонентов цитоплазмы сохранять и передавать от родителей к потомкам, из поколения в поколение часть наследственной информации. Конечно, ведущую роль выполняют гены хромосом в процессе наследования большей доли признаков организма, но внеядерная наследственность тоже играет достаточно значительную роль.
Готовые работы на аналогичную тему
Этот вид наследственности связан напрямую с двумя видами генетических явлений.
Первое - это с наследованием признаков, обусловленных внеядерными генами, которые содержатся в специальных органеллах. Такими органеллами в клетках являются митохондрии, пластиды.
Второе - с появлением у потомков признаков, которые обусловлены ядерными генами родительских форм, но на развитие и проявление их влияет цитоплазма яйцеклетки.
Наличие генов в полуавтономных органеллах, способных к самоудвоению - митохондриях и пластидах - было обнаружено уже в начале $XX$ века.
Пластидное наследование
Цитоплазматическая наследственность, которая связана с генами пластид, свойственна многим цветочным растениям, например, таким как львиный зев, ночная красавица. Среди них встречаются формы с пестрыми листьями. Даный признак может передаваться только по материнской линии.
Степень пестроты листьев объясняется тем, что некоторые части пластид не способны образовывать хлорофилл. Кроме того, еще во время образования гамет (яйцеклеток и спермиев) пластиды попадают к яйцеклеткам, а не к спермиям. Размножающиеся делением пластиды имеют генетическую непрерывность: от зеленых пластид образуются зеленые, а бесцветные дают начало бесцветным.
При делении клетки пластиды разных типов распределяются случайным образом, в результате этого образуются клетки с бесцветными, зелеными или пластидами обоих типов вместе.
Митохондриальная наследственность
Цитоплазматическая наследственность, которая связана с митохондриями, рассматривалась на примере дрожжей. В их митохондриях были найдены гены, от которых зависит отсутствие или наличие дыхательных ферментов, также они определяют устойчивость к определенным антибиотикам.
Влияние на формирование некоторых состояний определенных признаков потомков ядерных генов материнского организма через цитоплазму яйцеклетки можно проследить на примере пресноводного брюхоногого моллюска-прудовика. Он имеет формы с разными состояниями наследственного признака - закрученности раковины направленной влево или вправо. Аллель, которая определяет правозакрученность ракушки, доминирует над левозакрученностью, но направление закрученности определяют только гены материнского организма. Так, например, особи, которые по рецессивному признаку левозакрученности гомозиготные, могут иметь правозакрученную раковину, в том случае, если происходят от материнского организма, который и обладал доминантной аллелью правозакрученности.

Наряду с ядерными генами, локализованными в хромосомах, обнаружены факторы наследственности, расположенные в цитоплазме. Их называют плазмогенами (плазмидами). Химическую основу плазмогенов составляют молекулы ДНК. Установлено, что пластиды, митохондрии и некоторые другие органоиды содержат ДНК. В цитоплазме может находиться и чужеродная ДНК вирусов и плазмиды бактерий. Внеядерная ДНК способна реплицироваться независимо от репликации хромосом. Плазмогены находятся под контролем ядерных генов. Цитоплазматическое наследование идет по материнской линии, т.е. через цитоплазму яйцеклетки, так как сперматозоид почти не содержит ее. Критериями цитоплазматической наследственности являются: 1) отсутствие количественного менделеевского расщепления в потомстве; 2) невозможность выявить сцепление; 3) различные результаты реципрокных скрещиваний. Выделяют следующие основные виды цитоплазматической наследственности: пластидную, митохондриальную и псевдоцитоплазматическую.
| Вложение | Размер |
|---|---|
| tsitoplazmaticheskaya_nasledstvennost_i_gennaya_inzheneriya.doc | 48 КБ |
Предварительный просмотр:
ЦИТОПЛАЗМАТИЧЕСКАЯ НАСЛЕДСТВЕННОСТЬ И ГЕННАЯ ИНЖЕНЕРИЯ
1. Цитоплазматическая наследственность.
Наряду с ядерными генами, локализованными в хромосомах, обнаружены факторы наследственности, расположенные в цитоплазме. Их называют плазмогенами (плазмидами). Химическую основу плазмогенов составляют молекулы ДНК. Установлено, что пластиды, митохондрии и некоторые другие органоиды содержат ДНК. В цитоплазме может находиться и чужеродная ДНК вирусов и плазмиды бактерий. Внеядерная ДНК способна реплицироваться независимо от репликации хромосом. Плазмогены находятся под контролем ядерных генов. Цитоплазматическое наследование идет по материнской линии, т.е. через цитоплазму яйцеклетки, так как сперматозоид почти не содержит ее. Критериями цитоплазматической наследственности являются: 1) отсутствие количественного менделеевского расщепления в потомстве; 2) невозможность выявить сцепление ; 3) различные результаты реципрокных скрещиваний. Выделяют следующие основные виды цитоплазматической наследственности: пластидную, митохондриальную и псевдоцитоплазматическую.
1.1. Открытие пластидной наследственности принадлежит К. Корренсу (1908), описавшему пестролистность у ночной красавицы. У пестролистных растений часть пластид неспособна образовывать хлорофилл. Пластиды при митозе распределяются между дочерними клетками неравномерно. Часть клеток получает только нормальные пластиды (листья будут зелеными); часть клеток получает только аномальные пластиды (листья белые, без хлорофилла, растение погибает); часть клеток получает и аномальное и нормальные пластиды (пестрые листья, белые пятна на зеленых листьях).
1.2. Митохондриальная наследственность описана Б. Эфрусси (1949). Он обнаружил, что около 1% хлебных дрожжей дают карликовые колонии. Оказалось, что клетки карликовых колоний не имеют в митохондриях дыхательных ферментов вследствие мутации плазмогенов и поэтому растут очень медленно. Гены, кодирующие дыхательные ферменты, находятся в кольцевых молекулах ДНК митохондрий. Длина каждой такой молекулы примерно 15000 пар нуклеотидов. Расчеты показали, что объем собственной наследственной информации митохондрии недостаточен для воспроизведения всей совокупности РНК и белков органоида. Многие белки включаются в структуру митохондрий, будучи запрограммированными ядерными генами.
Геном митохондрий человека представлен кольцевой молекулой ДНК, содержащей 16569 пар нуклеотидов. В состав генома входят также гены р-РНК, 22 различных т-РНК, субъединиц I, II и III оксидазы цитохрома c , субъединицы 6 АТФазы, цитохрома b и девяти других пока неизвестных белков. В митохондриальной ДНК имеется очень мало не кодирующих, участков и транскрибируются обе ее цепочки. Имеются данные о том, что некоторые пороки развития человека обусловлены мутациями митохондриальных генов (митохондриальная цитопатия, не сращение верхних дуг позвонков и сращение нижних конечностей, старческое слабоумие, паркинсонизм и др.).
В цитоплазме бактерий, обнаружены автономно расположенные плазмиды, состоящие из кольцевых молекул двухцепочечной ДНК. Они обусловливают устойчивость бактерий к лекарствам (антибиотикам), программируют синтез некоторых ядов (гемолизина, энтеротоксина). Плазмиды обеспечивают также обмен генетической информацией между микроорганизмами. Внехромосомные молекулы ДНК широко используются в генной инженерии, так как они способны включить в себя генетический материал хромосом и переносить его в другие клетки.
1.3. Псевдоцитоплазматическая наследственность обусловлена попаданием в цитоплазму клеток участков чужеродной ДНК, т.е. она представляет собой разновидность внутриклеточного паразитизма. Так, у некоторых линий мух дрозофил существует повышенная чувствительность к СО 2 . Установлено, что эта способность обусловлена передачей особых вирусов через цитоплазму яйцеклетки.
2. Генная инженерия.
На основании достижений молекулярной биологии, биохимии и генетики в последние десятилетия интенсивно развивается новое направление в генетике – генная инженерия, целью которой является конструирование генетических структур по заранее намеченному плану, создание организмов с новой генетической программой путем переноса генетической информации из одного организма в другой.
2.1. Методы генной инженерии были разработаны в 60-70-х годах XX века. Они включают следующие основные этапы: 1) получение генетического материала (выделение природных генов или химический их синтез); 2) включение этих генов в автономно реплицирующуюся генетическую структуру (некоторую молекулу) и создание рекомбинантной ДНК; 3) введение рекомбинантных молекул ДНК в клетку-реципиент и включение ее в хромосомный аппарат; 4) отбор трансформированных клеток, в геном которых включен переносимый ген.
2.2. В настоящее время применяют несколько способов получения генов для пересадки. Если полностью расшифрована последовательность нуклеотидов, то ген может быть синтезирован химическим путем. Впервые искусственный ген аланиновый т-РНК, состоящий из 77 нуклеотидов, был синтезирован индийским ученым Г. Корана (1970 г.). В 1976 г. был синтезирован ген тирозиновой т-РНК, состоящий из структурной и регуляторной частей (промотор и теминатор), который при введении в бактериальную клетку нормально функционировал. Однако химическим способом удается синтезировать только небольшие по размеру гены прокариот.
Синтез сложных генов осуществляют с помощью процессов обратной транскрипции, в основе которых лежит метод ферментативного синтеза. Выделяют и-РНК, и на ней, как на матрице, с помощью фермента ревертазы (обратной транскриптазы) синтезируется комплементарная ей нить ДНК, а затем ее реплицируют (получают комплементарную цепочку). Гены, синтезированные с помощью ревертазы, не имеют регуляторной части и промотора и, вследствие этого, не могут функционировать в животных клетках. При переносе в бактерию к структурным генам присоединяют промотор микробной клетки, после чего транскриптон начинает работать.
Полученные различными способами гены соединяются с векторными молекулами, которыми чаще служат плазмиды бактерий. Кроме плазмид в качестве вектора используются фаги и вирусы. Они передают генетическую информацию посредством трансдукции. Кольцевая молекула ДНК плазмиды разрывается той же рестриктазой, что и выделенный ген. В области разрыва образуются липкие концы, комплементарные липким концам пересаживаемого гена. Фермент лигаза сшивает липкие концы гена и плазмиды. Получается рекомбинантная молекула ДНК, которая обладает способностью проникать в клетку-реципиент. Комбинируя различные рестриктазы и лигазы, можно разрезать нить ДНК в разных местах и получать рекомбинантные молекулы.
2.4. Так как не во все клетки попадут рекомбинированные молекулы ДНК, то с помощью специальных методов (чаще всего на селективных питательных средах) проводят отбор трансформированных клеток (с перенесенным геном). В дальнейшем проводят клонирование – размножение клеток с рекомбинантной ДНК – и получают клон клеток с заданными свойствами.
2.5. Методами генной инженерии получены клоны клеток кишечной палочки, способные продуцировать соматотропин и инсулин в промышленных масштабах. Обычно эти препараты получают из соответствующих желез животных. Преимущественно препаратов, полученных методами генной инженерии, заключается в возможности синтеза их в достаточных количествах, биохимически чистыми и абсолютно стерильными.
Генная инженерия относится к современным интенсивно развивающимся направлениям генетики. С использованием ее методов созданы растения, способные усваивать атмосферный азот, микроорганизмы, разрушающие углеводороды нефти и синтезирующие из них пищевые белки. Разработаны методы внесения генов патогенных вирусов в бактериальные клетки и приготовление из синтезированных ими белков противовирусных сывороток. В будущем генная инженерия поможет человечеству избавиться от ряда наследственных заболеваний, посредством пересадки в зародыш недостающих или замены мутантных генов.
В настоящее время накапливаются клонированные гены человека, некоторых животных и растений, т.е. создаются банки генов.
2.6. Объединение чужеродных генов в одной клетке чревато опасными последствиями. Плазмиды способны соединяться в любых комбинациях, независимо от видовых и иммунологических барьеров. Конструирование новых разновидностей болезнетворных бактерий, устойчивых к лекарственным препаратам, может привести к возникновению серьезных эпидемий. В 1973 г. была проведена первая международная конференция по предупреждению опасных последствий генной инженерии. Опыты на время были запрещены. В 1975 г. Р. Кертис получил мутант кишечной палочки, нежизнеспособный в естественных условиях в связи с нарушением синтеза оболочки. Неопасная для человека и животных бактерия может жить только в лабораторных условиях, и опыты по генной инженерии были продолжены. Все исследования по генной инженерии проводятся в специальных лабораториях, строжайше изолированных от окружающей среды, с обязательным соблюдением определенных мер безопасности.
2.7. Будущее генной инженерии базируется на следующих достижениях молекулярной биологии:
- возможность с помощью химических мутагенов вызывать специфические мутации в определенных генных локусах;
- возможность переноса генетической информации неполовым путем у эукариот (трансформация или трансдукция), что позволит проводить генную терапию заболеваний;
- замена дефектных генов с использованием ДНК вирусов в качестве переносчиков;
- включение в геном человека искусственно синтезированных генов.
3. Перспективы генной терапии.
Необходимо различать 2 цели генной терапии – коррекцию генетических дефектов в соматических клетках и коррекцию их в гаметах или на самых ранних стадиях развития зиготы.
В настоящее время единственными клетками человека, которые можно использовать для переноса генов, являются клетки костного мозга или фибробласты. Эти клетки можно извлечь из организма, вырастить в культуре, перенести в них нужный ген и снова ввести пациенту. Наиболее перспективным является перенос нужных генов, связанный с использованием ретровирусов. Чтобы применять на практике методы генной инженерии, нужно быть уверенным в их безопасности. Например, человеческие онкогены по структуре отчасти гомологичны ретровирусам и при заражении клеток такими вирусами возможна их модификация и превращение в онкогены.
В экспериментах на мышах проведена генная терапия на уровне зиготы: в оплодотворенные яйцеклетки мышей карликовой линии вводили гены гормона роста крыс. При этом часть потомков (6 из 41) достигли гигантских размеров. Очевидно, что вновь встроенные гены не подвергаются нормальной регуляции, так как не удается внедрить их в места обычной локализации в хромосоме. Встраивание, происходит в случайном порядке и в некоторых опытах, это вызывало у мышей-реципиентов серьезные нарушения (мутации) работы нормальных генов в участках встраивания. По мнению большинства медицинских генетиков, метод генной терапии не следует в обозримом будущем применять к оплодотворенным клеткам человека, так как слишком велика опасность изменения генетической конституции человека.

Митохондриальное (цитоплазматическое) наследование характерно для особого класса наследственной патологии – митохондриальных болезней. Каждая клетка содержит тысячи копий митохондриальной ДНК. Ряд редких болезней с необычной комбинацией неврологических и миопатических признаков, кардиомиопатии, диабет, как оказалось, возникают вследствие мутаций митохондриальных генов. Неудивительно, что головной мозг, мышцы и сердце поражаются в большей степени, поскольку эти органы наиболее энергозависимы. Митохондриальные болезни поражают оба пола, но передаются только через женщин (рис. 15). У большинства людей митохондриальная ДНК идентична во всех митохондриях (гомоплазмия). В случае мутаций ДНК в части митохондрий, у индивидуума будет две популяции митохондрий – нормальные и мутантные, т. е. гетероплазмия. Доля митохондрий с мутантной ДНК варьирует между клетками и тканями. Это является объяснением различной тяжести течения заболевания у людей с митохондриальными болезнями. Ряд митохондриальных белков кодируется ядерными генами, а мутации в них нарушают функцию митохондрий. Напрмер, мутации генов белков комплекса цитохрома С наследуются аутосомно-рецессивно, а мутации Х-сцепленного гена G4.5 (TAZ) вызывают синдром Барта (кардиоскелетную миопатию с нейтропенией и аномальными митохондриями) у мальчиков.
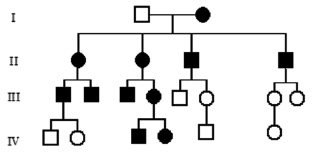
Рис. 15. Родословная при митохондриальном типе наследования
Множественные аллели и комплексные признаки
Выше рассмотрены признаки, с которыми связаны только два аллеля – нормальный и мутантный. Некоторые гены имеют более двух аллельных форм, т. е. множественные аллели. Некоторые из них могут быть доминантными, другие – рецессивными по отношению к нормальному аллелю. Пример множественных аллелей – наследование групп крови человека.
Развитие генетики сделало возможным исследование комплексных признаков, которые формируются при взаимодействии нескольких генов. На этой основе возникла концепция олигогенного (дигенного и триаллельного) наследования.
При дигенном наследовании наблюдается аддитивный эффект гетерозиготных мутаций в двух различных локусах. Например, одна из форм пигментного ретинита, приводящая к потере зрения, вызвана гетерозиготностью по мутациям двух генов (ROM1 и PRPH). Оба эти гена кодируют белки, присутствующие в фоторецепторах сетчатки глаза. Индивидуумы, гетерозиготные по мутации только одного из этих двух генов, не имеют клинических проявлений.
Триаллельное наследование можно рассмотреть на примере синдрома Барде-Бидля – редкого заболевания, характеризующегося ожирением, полидактилией, аномалиями почек, пигментным ретинитом и когнитивными нарушениями. Семь различных генных локусов, мутации в которых ведут к синдрому Барде–Бидля, были идентифицированы. До недавнего времени считалось, что заболевание наследуется аутосомно-рецессивно. Однако, сейчас известно, что есть одна форма синдрома, когда индивидуум, гомозиготный по мутациям одного локуса, является также гетерозиготным по мутации другого локуса. Таким образом, для того, чтобы заболевание проявлялось, необходимо три мутантных аллеля.
Антиципация. При некоторых аутосомно-доминантных болезнях манифестация симптомов более ранняя и течение болезни более тяжелое у потомков по сравнению с их родителями, также страдающими этим заболеванием. Феномен увеличения тяжести болезни из поколения в поколение называют антиципацией. Одним из объяснений антиципации является экспансия нестабильных триплетных повторов. В качестве примеров можно привести такие болезни экспансии триплетных повторов, как миотоническая дистрофия, хорея Гентингтона, болезнь Кеннеди.
Читайте также: