
Окислительное фосфорилирование биохимия реферат
Обновлено: 05.07.2024
По современным представлениям внутренняя митохондриальная мембрана содержит ряд мультиферментных комплексов, включающих множество ферментов. Эти ферменты называют дыхательными ферментами, а последовательность их расположения в мембране — цепью тканевого дыхания или цепью переноса электронов (англ. electron transport chain). Молекулы НАДН и ФАДН2, образуемые в реакциях окисления углеводов, жирных… Читать ещё >
Окислительное фосфорилирование. Энергетический обмен ( реферат , курсовая , диплом , контрольная )
Молекулы НАДН и ФАДН2, образуемые в реакциях окисления углеводов, жирных кислот, спиртов и аминокислот, далее поступают в митохондрии, где ферментами дыхательной цепи (цепи переноса электронов) осуществляется процесс окислительного фосфорилирования.
Окислительное фосфорилирование — это многоэтапный процесс, происходящий во внутренней мембране митохондрий и заключающийся в окислении восстановленных эквивалентов (НАДН и ФАДН2) ферментами дыхательной цепи и сопровождающийся синтезом АТФ.
Впервые в 1961 году механизм окислительного фосфорилирования был предложен Питером Митчеллом в виде хемиосмотической теории. Согласно этой гипотезе перенос электронов, происходящий на внутренней митохондриальной мембране, сопровождается переносом ионов Н + из матрикса митохондрий в межмембранное пространство (протонный насос), что создаёт трансмембранный градиент концентрации ионов Н + между цитозолем и замкнутым внутримитохондриальным пространством. Ионы водорода в норме способны возвращаться в матрикс митохондрий только одним способом — через специальный фермент, образующий АТФ — АТФ-синтазу.
По современным представлениям внутренняя митохондриальная мембрана содержит ряд мультиферментных комплексов, включающих множество ферментов. Эти ферменты называют дыхательными ферментами, а последовательность их расположения в мембране — цепью тканевого дыхания или цепью переноса электронов (англ. electron transport chain).

Волков Н.И., Несен Э.Н., Осипенко А.А., Корсун С.Н. Биохимия мышечной деятельности
- формат djvu
- размер 8.53 МБ
- добавлен 16 октября 2010 г.
Колесниченко А.В., Войников В.К. Белки низкотемпературного стресса растений
- формат pdf
- размер 3.36 МБ
- добавлен 05 ноября 2010 г.
Иркутск: Арт-Пресс, 2003. – 196 с. Монография посвящена обзору исследований по одной из проблем современной физиологии растений – выяснению физиологических и биохимических механизмов адаптации растений к низким температурам. В последние годы установлено, что в ответ на низкотемпературный стресс в растениях происходит синтез специфических, стрессовых белков. Среди них антифризные белки, регулирующие и предохраняющие клетки растений от повреждения.
Контрольная работа по биохимии
- формат doc
- размер 76.5 КБ
- добавлен 17 апреля 2011 г.
1. Что такое микроэлементы. Привести примеры, описать роль железа в составе гемоглобина крови, миоглобина мышц и цитохромов митохондрий 2. Что такое гормоны. Гормоны каких эндокринных желез регулирует обмен углеводов в организме 3. Что такое окислительное фосфорилирование. Охарактеризовать аэробный ресинтез АТФ по кинетическим показателям (быстроте развертывания, метаболической мощности, емкости и эффективности). В каких видах спорта и почему пр.
Курсовая работа - Биологическое окисление
- формат doc
- размер 63.1 КБ
- добавлен 16 декабря 2009 г.
Уральская государственная медицинская академия. кафедра биоорганической и биологической химии. студентки педиатрического факультета. Содержание. Введение. Общие представления о биологическом окислении. Окислительно-восстановительные системы и потенциалы. Пути использования кислорода в клетке. Оксидазный путь использования кислорода. Митохондрии. Ферменты, их локализация и значение в процессах окисления. Этапы утилизации энергии питательных вещест.
Лекции- Основы биохимии
- формат doc
- размер 2.56 МБ
- добавлен 11 ноября 2010 г.
Курс лекций Кемеровский технологический институт пищевой промшленности, 2002 г. 119 стр. Оглавление введение Общий химический состав живых организмов Белковые вещества Общая характеристика белков. Аминокислоты - структурные элементы белков. Строение и пространственная структура белковой молекулы. Физико-химические свойства белков. Выделение белков из биологических объектов. Номенклатура и классификация белков. Нуклеиновые кислоты Химический со.
Ленинджер А. Основы биохимии. Том 2
- формат djvu
- размер 6.03 МБ
- добавлен 28 февраля 2010 г.
В 3-х т. Т. 2. Пер. с англ. - М.: Мир, 1985. - 368 с., ил. Во второй том вошли материалы по биоэнергетике и метаболизму клетки. Рассмотрены роль глюкозы в биоэнергетических процессах, цикл лимонной кислоты, электронный транспорт, окислительное фосфорилирование, регуляция образования АТФ, окисление жирных кислот в тканях животных, окислительный распад аминокислот, биосинтез углеводов, аминокислот, липидов, нуклеотидов; фотосинтез. Для биологов ра.
Реферат-Биохимия гормонов гипофиза и гипоталамуса
- формат docx
- размер 60.72 КБ
- добавлен 30 сентября 2011 г.
ВГМУ,2011 год.10 стр.Реферат содержит краткую характеристику эндокринной системы и гормонов,а так же более подробное описание гормонов гипофиза и гипоталамуса.
Реферат-Интеграция обмена углеводов, белков и жиров в организме. Транспортные системы в организме человека
- формат rtf
- размер 156.21 КБ
- добавлен 27 февраля 2011 г.
Реферат, 20 стр. Раскрывается вопрос интеграции метаболизма в человеческом организме; системы, осуществляющие интеграцию, а также роль транспортных систем в осуществлении метаболических процессов.
Таганович А.Д. и др. Биологическая химия
- формат doc
- размер 5.6 МБ
- добавлен 16 июня 2010 г.
Минск: БГМУ, 2007 г. , 118 с. Необходимость в составлении данного учебно-методического пособия вызвана стрем-лением преподавателей кафедры облегчить усвоение лекционного материала по биологической химии. С этой целью в данном учебно-методическом пособии было решено сконцентрировать рисунки, схемы, графики, основные понятия, определения, изложенный простым языком краткий текст лекций, то есть все то, что призвано максимально облегчить понимание и.
Шпоры по углеводам
- формат docx
- размер 5.84 МБ
- добавлен 18 сентября 2011 г.
Химия углеводов: биологическая роль углеводов классификация углеводов моносахариды основные реакции моносахаридов, продукты реакций и их свойства олигосахариды полисахариды гомополисахариды гетерополисахариды метаболизм углеводов переваривание и всасывание углеводов синтез и распад гликогена синтез гликогена (гликогенез) распад гликогена (гликогенолиз) гликолиз спиртовое брожение включение других углеводов в процесс гликолиза глюконеогенез аэроб.
ЗАКЛЮЧЕНИЕ
Таким образом, окислительное фосфорилирование – это катаболический процесс синтеза АТФ, являющийся завершающим этапов дыхания. При этом НАДН и ФАДН2 окисляются молекулярным кислородом с постепенным переносом электроном, насчитывающим четыре этапа. Весь процесс в совокупности называется дыхательной цепью переноса электронов.
Процесс окисления сопряжен с процессом фосфорилирования вероятно благодаря протонному потенциалу, возникающему на внутренней мембране митохондрий в процессе транспорта электронов.
Исходя из всего вышесказанного, можно заключить, что окислительное фосфорилирование – это сложнейший процесс, который охватывает десятки реакций и задействует сотни органических и неорганических веществ.
Содержание
СПИСОК СОКРАЩЕНИЙ 3
ВВЕДЕНИЕ 4
Глава 1. Место окислительного фосфорилирования в метаболизме 5
Глава 2. Общие сведения об организации дыхательной цепи транспорта электронов. Дыхательные комплексы I, II, III, IV 7
Глава 3. Окислительное фосфорилирование 10
ЗАКЛЮЧЕНИЕ 14
СПИСОК ЛИТЕРАТУРЫ 15
Введение
ВВЕДЕНИЕ
Живой организм – это открытая система, в которую непрерывно поступает вещества и энергия и из которой выводятся продукты обмена веществ. При этом в клетках происходят сотни реакций, формирующих метаболические пути (последовательное превращение одного вещества в другое), совокупность которых называется метаболизмом
Глобально все метаболические пути можно объединить в две взаимосвязанные системы, а именно:
1. Катаболизм, т.е. энергетический обмен, или диссимиляция, сутью которого является распад поступивших питательных веществ на более простые и окисление различных веществ, сопровождающееся выделением энергии. Конечными продуктами катаболизма у животных и человека является вода, углекислый газ и мочевина.
2. Анаболизм, т.е. пластический обмен, или ассимиляция, представляющий со бой совокупность биосинтетических процессов, происходящих с затратой энергии [3, с. 108, 264].
При катаболизме происходит расщепление полимерных органических веществ (белки, жиры, сложные углеводы) до мономеров, и их последующее окисление с выделением энергии, которая запасается в молекулах АТФ. Один из вариантов образования молекул АТФ, который осуществляется посредством переноса протонов и электронов с помощью окислительно-восстановительных ферментов называется окислительным фосфорилированием [9, с. 216].
Целью данной работы является систематизация знаний об окислительном фосфорилировании.
Для достижения поставленной цели необходимо решить ряд задач, а именно:
1. Определить роль и место окислительного фосфорилирования в метаболическом процессе;
2. Изучить этапы окислительного фосфорилирования.
Фрагмент работы для ознакомления
Список литературы
СПИСОК ЛИТЕРАТУРЫ
1. Березов Т.Т., Коровкин Б.Ф. Биологическая химия: Учебник. – 3-е изд., перераб. и доп. – М.: Медицина, 1998. – 704 с.
2. Биологическая химия: Учебник / В.К. Кухта, Т.Е. Морозкина, З.И. Олецкий, А.Д. Таганович; Под ред. А.Д. Тагановича. – Минск: Асар, М.: Издательство БИНОМ, 2008. – 688 с.
3. Биохимия: Учебник/ Под ред. Е.С. Северина. – 2-е изд., испр. – М.: ГЭОТАР-Медиа, 2004. – 784 с.
4. Василенко Ю.К. Биологическая химия: Учебное пособие / Ю.К. Василенко. – Пятигорск: ГОУ ВПО Пятигорская ГФА Росздрава, 2005. – 418 с.
5. Жеребцов Н.А., Попова Т.Н., Артюхов В.Г. Биохимия: Учебник. – Воронеж: издательство Воронежского государственного университета, 2002. – 696 с.
6. Кнорре Д.Г., Мызина С.Д. Биологическая химия: Учебник для хим., биол. и мед. спец. Вузов. – 3-е изд., испр. – М.: Выысш. Школа, 2000. – 479 с.
7. Комов В.П. Биохимич: Учебник для вухов/ В.П. Комов, В.Н. Шведова. – М.: Дрофа, 2004. – 640 с.
8. Николаев А.Я. Биологическая химия. – 3-е изд., перераб. и доп. – М.: Медицинское информационное агентство, 2004. – 566 с.
9. Новакшанова А.Л. Биохимия для технологов: Учебник и практикум для академического бакалавриата/ А.Л. Новокшанова. – М.: издательство Юрайт, 2015. – 508 с.
Пожалуйста, внимательно изучайте содержание и фрагменты работы. Деньги за приобретённые готовые работы по причине несоответствия данной работы вашим требованиям или её уникальности не возвращаются.
* Категория работы носит оценочный характер в соответствии с качественными и количественными параметрами предоставляемого материала. Данный материал ни целиком, ни любая из его частей не является готовым научным трудом, выпускной квалификационной работой, научным докладом или иной работой, предусмотренной государственной системой научной аттестации или необходимой для прохождения промежуточной или итоговой аттестации. Данный материал представляет собой субъективный результат обработки, структурирования и форматирования собранной его автором информации и предназначен, прежде всего, для использования в качестве источника для самостоятельной подготовки работы указанной тематики.
ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ, синтез АТФ из аденозиндифосфата и неорг. фосфата, осуществляющийся в живых клетках, благодаря энергии, выделяющейся при окислении орг. в-в в процессе клеточного дыхания. В общем виде окислительное фосфорилирование и его место в обмене в-в можно представить схемой:
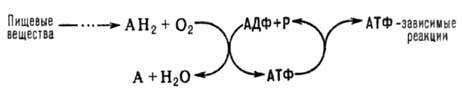
АН 2 -орг. в-ва, окисляемые в дыхат. цепи (т. наз. субстраты окисления, или дыхания), АДФ-аденозиндифосфат, Р-неорг. фосфат.
Поскольку АТФ необходим для осуществления мн. процессов, требующих затраты энергии (биосинтез, совершение мех. работы, транспорт в-в и др.), Окислительное фосфорилирование играет важнейшую роль в жизнедеятельности аэробных организмов. Образование АТФ в клетке происходит также благодаря др. процессам, напр. в ходе гликолиза и разл. типов брожения, протекающих без участия кислорода. Их вклад в синтез АТФ в условиях аэробного дыхания составляет незначит. часть от вклада окислительного фосфорилирования (ок. 5%).
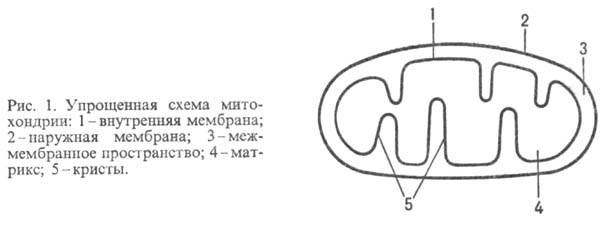
У животных, растений и грибов окислительное фосфорилирование протекает в спе-циализир. субклеточных структурах-митохондриях (рис. 1); у бактерий ферментные системы, осуществляющие этот процесс, находятся в клеточной мембране.
Митохондрии окружены белково-фосфолипидной мембраной. Внутри митохондрий (в т. наз. матриксе) идет ряд метаболич. процессов распада пищ. в-в, поставляющих субстраты окисления АН 2 для окислительного фосфорилирования. Наиб. важные из этих процессов -трикарбоновых кислот цикл и т. наз.-окисление жирных к-т (окислит. расщепление жирной к-ты с образованием ацетил-кофермента А и к-ты, содержащей на 2 атома С меньше, чем исходная; вновь образующаяся жирная к-та также может подвергаться -окислению). Интермедиаты этих процессов подвергаются дегидрированию (окислению) при участии ферментов дегидрогеназ; затем электроны передаются в дыхат. цепь митохондрий-ансамбль окислит.-восстановит. ферментов, встроенных во внутр. митохонд-риальную мембрану. Дыхат. цепь осуществляет многоступенчатый экзэргонич. перенос электронов (сопровождается уменьшением своб. энергии) от субстратов к кислороду, а высвобождающаяся энергия используется расположенным в той же мембране ферментом АТФ-синтетазой, для фос-форилирования АДФ до АТФ. В интактной (неповрежденной) митохондриальной мембране перенос электронов в дыхат. цепи и фосфорилирование тесно сопряжены между собой. Так, напр., выключение фосфорилирования по исчерпании АДФ либо неорг. фосфата сопровождается торможением дыхания (эффект дыхат. контроля). Большое число повреждающих митохондриальную мембрану воздействий нарушает сопряжение между окислением и фосфори-лированием, разрешая идти переносу электронов и в отсутствие синтеза АТФ (эффект разобщения).
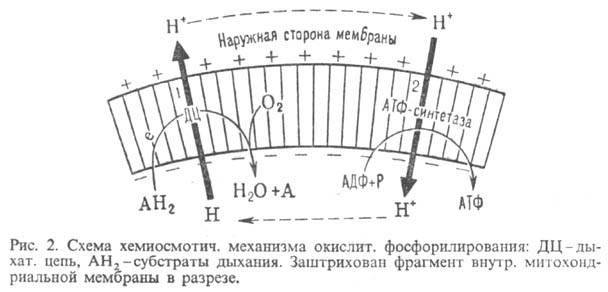
Механизм окислительного фосфорилирования можно представить схемой: Перенос электронов (дыхание) А ~ ВАТФ А ~ В-высокоэнергетич. интермедиат Предполагалось, что А ~ В - хим. соед. с макроэргич. связью, напр. фосфорилир. фермент дыхат. цепи (хим. гипотеза сопряжения), или напряженная конформация к.-л. белка, участвующего в окислительном фосфорилировании (конформац. гипотеза сопряжения). Однако эти гипотезы не получили эксперим. подтверждения. Наиб. признанием пользуется хемиосмотич. концепция сопряжения, предложенная в 1961 П. Митчеллом (за развитие этой концепции в 1979 ему присуждена Нобелевская премия). Согласно этой теории, своб. энергия транспорта электронов в дыхат. цепи затрачивается на перенос из митохондрий через митохондриальную мембрану на ее наружную сторону ионов Н + (рис. 2, процесс 1). В результате на мембране возникает разность электрич. потенциалов и разность хим. активностей ионов Н + (внутри митохондрий рН выше, чем снаружи). В сумме эти компоненты дают трансмембранную разность электрохим. потенциалов ионов водорода между матриксом митохондрий и внеш. водной фазой, разделенными мембраной:
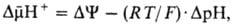
где R-универсальная газовая постоянная, T-абс. т-ра, F- число Фарадея. Величина обычно составляет ок. 0,25 В, причем осн. часть (0,15-0,20 В) представлена электрич. составляющей . Энергия , выделяющаяся при движении протонов внутрь митохондрий по электрич. полю в сторону меньшей их концентрации (рис. 2, процесс 2), используется АТФ-синтетазой для синтеза АТФ. Т. обр., схему окислительного фосфорилирования, согласно этой концепции, можно представить в след. виде:
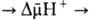
Перенос электронов (дыхание) АТФ
Сопряжение окисления и фосфорилирования через позволяет объяснить, почему окислительное фосфорилирование, в отличие от гликолитич. ("субстратного") фосфорилирования, протекающего в р-ре, возможно лишь в замкнутых мембранных структурах, а также почему все воздействия, снижающие электрич. сопротивление и увеличивающие протонную проводимость мембраны, подавляют ("разобщают") окислительное фосфорилирование. Энергия , помимо синтеза АТФ, может непосредственно использоваться клеткой для др. целей - транспорта метаболитов, движения (у бактерий), восстановления нико-тинамидных коферментов и др.
В дыхат. цепи имеется неск. участков, к-рые характеризуются значит. перепадом окислит.-восстановит. потенциала и сопряжены с запасанием энергии (генерацией ). Таких участков, наз. пунктами или точками сопряжения, обычно три: НАДН: убихинон-редуктазное звено ( 0,35-0,4 В), убихинол: цитохром-c-редуктазное звено (~ ~ 0,25 В) и цитохром-с-оксидазный комплекс (~ 0,6 В)-пункты сопряжения 1, 2 и 3 соотв. (рис. 3). Каждый из пунктов сопряжения дыхат. цепи м.б. выделен из мембраны в виде индивидуального ферментного комплекса, обладающего окислит.-восстановит. активностью. Такой комплекс, встроенный в фосфолипидную мембрану, способен функционировать как протонный насос.
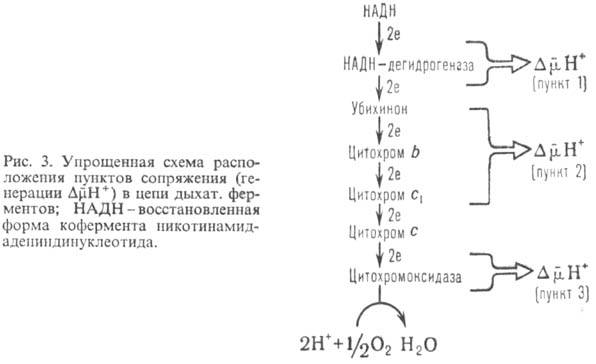
Обычно для характеристики эффективности окислительного фосфорилирования используют величины Н + /2е или q/2e, указывающие сколько протонов (либо электрич. зарядов) переносится через мембрану при транспорте пары электронов через данный участок дыхат. цепи, а также отношение Н + /АТФ, показывающее, сколько протонов нужно перенести снаружи внутрь митохондрий через АТФ-синтетазу для синтеза 1 молекулы АТФ. Величина q/2e составляет для пунктов сопряжения 1, 2 и 3 соотв. 3-4, 2 и 4. Величина Н + /АТФ при синтезе АТФ внутри митохондрий равна 2; однако еще один Н + может тратиться на вынос синтезированного АТФ 4- из матрикса в цитоплазму переносчиком адениновых нуклеотидов в обмен на АДФ -3 . Поэтому кажущаяся величина Н + / АТФ наружн равна 3.
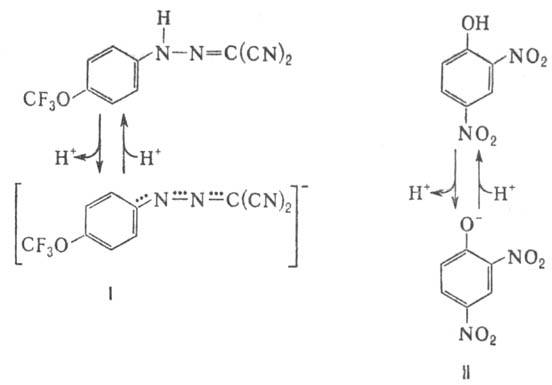
В организме окислительное фосфорилирование подавляется мн. токсичными в-вами, к-рые по месту их действия можно разделить на три группы: 1) ингибиторы дыхат. цепи, или т. наз. дыхат. яды (см. Дыхание). 2) Ингибиторы АТФ-синтетазы. Наиб. распространенные ингибиторы этого класса, употребляемые в лаб. исследованиях, - антибиотик олигомицин и модификатор карбоксильных групп белка дициклогексилкарбодиимид. 3) Т. наз. разобщители окислительного фосфорилирования. Они не подавляют ни перенос электронов, ни собственно фосфорилирование АДФ, но обладают способностью уменьшать величину на мембране, благодаря чему нарушается энергетич. сопряжение между дыханием и синтезом АТФ. Разобщающее действие проявляет большое число соед. самой разнообразной хим. структуры. Классич. разобщители - в-ва, обладающие слабыми кислотными св-вами, способные проникать через мембрану как в ионизованной (депротонированной), так и в нейтральной (протонированной) формах. К таким в-вам относят, напр., 1-(2-дицианометилен)гидразино-4-трифтор-метоксибензол, или карбонилцианид-n-трифторметокси-фенилгидразон, и 2,4-динитрофенол (соотв. ф-лы I и II; показаны протонир. и депротонир. формы).
Двигаясь через мембрану в электрич. поле в ионизованной форме, разобщитель уменьшает; возвращаясь обратно в протонир. состоянии, разобщитель понижает (рис. 4). Т. обр., такой "челночный" тип действия разобщителя приводит к уменьшению
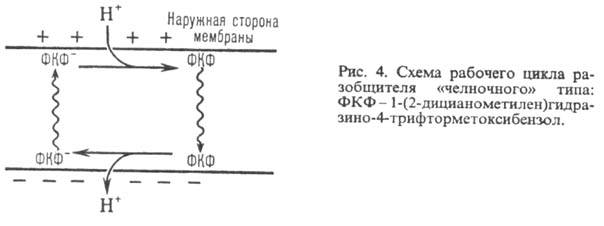
Разобщающим действием обладают также ионофоры (напр., грамицидин), повышающие электропроводность мембраны в результате образования ионных каналов или в-ва, разрушающие мембрану (напр., детергенты).
О кислительное фосфорилирование открыто В. А. Энгельгардтом в 1930 при работе с эритроцитами птиц. В 1939 В. А. Белицер и Е. Т. Цыбакова показали, что окислительное фосфорилирование сопряжено с переносом электронов в процессе дыхания; к такому же заключению несколько позднее пришел Г. М. Калькар.
Читайте также: