
Интеграция и взаимодействие клеток реферат
Обновлено: 05.07.2024
Дифференциация и интеграция биологических структур
в филогенезе. Сущность живого, его основные признаки. Структурные уровни организации живой материи
.
Содержание
Введение………………………………………………………………. стр.,3
1.Дифференциация и интеграция биологических структур
в филогенезе………………………………………………………….. стр.,4,5
2. Полифункциональность и количественное изменение функции биологических структур………………………………………. стр.,6,7,8,9,10
3. Соответствие структуры и функции в живых системах………. стр.,11,12
3.1Сущность живого, его основные признаки…………………..стр.,13,14,15
3.2Структурные уровни организации живой материи……….. ..стр.,16,17,18
4. Принципы активации и интенсификации функций органа…… стр.,19,20
Заключение……………………………………………………………… стр.,21
Список использованной литературы…………………………………. стр.,22
Введение
Организм, или особь - отдельное живое существо, в процессе онтогенеза проявляющее все свойства живого. Постоянное взаимодействие особи с окружающей средой в виде организованных потоков энергии и вещества поддерживает ее целостность и развитие. В структурном отношении организм представляет собой интегрированную иерархическую систему, построенную из клеток, тканей, органов и систем, обеспечивающих его жизнедеятельность. Подробнее остановимся на органах и системах жизнеобеспечения. Органом называют исторически сложившуюся специализированную систему тканей, характеризующуюся ограниченностью, постоянством формы, локализации, внутренней конструкции путей кровообращения и иннервации, развитием в онтогенезе и специфическими функциями.
Фрагмент работы для ознакомления
Список литературы
Пожалуйста, внимательно изучайте содержание и фрагменты работы. Деньги за приобретённые готовые работы по причине несоответствия данной работы вашим требованиям или её уникальности не возвращаются.
* Категория работы носит оценочный характер в соответствии с качественными и количественными параметрами предоставляемого материала. Данный материал ни целиком, ни любая из его частей не является готовым научным трудом, выпускной квалификационной работой, научным докладом или иной работой, предусмотренной государственной системой научной аттестации или необходимой для прохождения промежуточной или итоговой аттестации. Данный материал представляет собой субъективный результат обработки, структурирования и форматирования собранной его автором информации и предназначен, прежде всего, для использования в качестве источника для самостоятельной подготовки работы указанной тематики.
Межклеточные взаимодействия — это взаимодействия клеток друг с другом.Могут быть как дистантными,на расстоянии, так и контактными.Дистантные взаимодействия осуществляются при помощи растворимых веществ, секретируемых клетками в окружающую их среду и воздействующих на другие клетки. Эти вещества называются медиаторами,или посредниками. В качестве медиаторов могут выступать гормоны, биогенные амины, антитела и многие другие биологически активные вещества, эти вещества воздействуют на репепторный аппарат клеток, с которыми взаимодействует выделившая медиатор клетка. Следовательно, дистантные межклеточные взаимодействия опосредуют действие на клетки гормонов, имеют место при иммунном ответе, эмбриональном развитии (эмбриональная индукция,см. эмбриологию) и при многих других важных клеточных реакциях.
Кроме того, в многоклеточном организме все клетки связаны между собой при помощи межклеточных контактов (контактные межклеточные взаимодействия). Контактные взаимодействия состоят из нескольких фаз и включают как начальный этап дистантные взаимодействия:
1. Узнавание одной клеткой другой клетки (может быть дистантным при посредстве медиаторов и контактным при посредстве рецепторов).
2. Установление между клетками непрочных связей.
3. Формирование устойчивых межклеточных контактов. Вторая и третья фазы осуществляются при помощи молекул клеточной адгезии.
Все межклеточные контакты делятся на три основных типа (рис. 3.15, 3.16):
1. Адгезионные контакты,которые механически соединяют клетки между собой. Основной тип адгезионных контактов — десмосомы. Бывают трех типов:
— точечные десмосомы (пятно десмосомы). Они скрепляют клетки в отдельных местах. При этом с внутренней стороны клеточных мембран двух
клеток находится электрошюплотная пластинка, связанная с сетью кератиновых микрофиламент. Эти филаменты заканчиваются в пластинке или проходят мдоль ее поверхности. Прилегающие друг к другу пластинки двух клеток соединены через межклеточное пространство волокнами из белка неизвестной природы. В межклеточном пространстве есть электронноплотный материал;
— опоясывающие десмосомы (зоны десмосомы). Они идут вблизи апи-кального конца клеток по их периметру в виде полосы. Эта полоса состоит из пучков актиновых филаментов, локализующихся со стороны цитоплазмы. В межклеточном пространстве есть электронноплотный материал;
— полудесмосомы. Представляют собой как бы половинку точечной десмосомы. Прикрепляют эпителиальные клетки к базальной мембране.
| В функционировании адгезионных контактов важную роль играют адгезионные молекулы, такие, как Е-кадгерин, дссмоколлины, десмоглеины и др. |
2. Плотные контакты.Это разновидность замыкающих контактов. Данный тип контактов не только механически связывает клетки друг с другом, но и препятствует прохождению между ними молекул. В плотных контактах клеточные мембраны подходят друг к другу на расстояние до 5 нм и связываются друг с другом при помощи специальных белков.
3. Проводящие контакты.В этих контактах может осуществляться передача малых молекул из одной клетки в другую. При этом мембраны двух клеток подходят друг к другу на расстояние до 3 нм и образуют каналы — коннексоны.Через коннексоны между клетками осуществляется свободный обмен низкомолекулярными веществами (электролитами, витаминами, нуклеотидами, АТФ, сахарами, аминокислотами и др.). Таким образом, этот тип контактов играет важную роль не только в механической, но и в химической коммуникации клеток. Пример таких контактов — щелевые контакты: нексусымежду мышечными клетками в гладкой и сердечной мускулатуре. При этом возбуждение передается с одной клетки на другую. Второй пример — синапсы— контакты между нервными клетками.
Кроме этих основных видов межклеточных контактов, выделяют также интердигитации— или межпальцевые соединения, когда цитоплазма с покрывающей ее цитолеммои одной клетки в виде пальца вклинивается в цитоплазму другой клетки и наоборот. Интердигитации резко увеличивают прочность межклеточных соединений, а кроме того, увеличивают площадь межклеточных взаимодействий, благодаря чему возрастает межклеточный обмен метаболитами.
Дифференциация клеток
Описание: Роль ядра и цитоплазмы в клеточной дифференциации Как возникают разнообразные типы клеток в многоклеточном организме Известно что организм человека развившийся всего из 1 исходной клетки зиготы содержит более 100 различных типов клеток. Современная биология на базе представлений эмбриологии молекулярной биологии и генетики считает что индивидуальное развитие от одной клетки до многоклеточного зрелого организма результат последовательного избирательного включения в работу разных генных участков хромосом в различных клетках.
Дата добавления: 2015-01-12
Размер файла: 12.79 KB
Работу скачали: 62 чел.
Поделитесь работой в социальных сетях
Если эта работа Вам не подошла внизу страницы есть список похожих работ. Так же Вы можете воспользоваться кнопкой поиск
Роль ядра и цитоплазмы в клеточной дифференциации
Как возникают разнообразные типы клеток в многоклеточном организме? Известно, что организм человека, развившийся всего из 1 исходной клетки зиготы, содержит более 100 различных типов клеток. Каким образом возникает это разнообразие, сегодня до конца не ясно, так как еще мало конкретных данных, касающихся анализа путей появления тех или иных клеточных типов.
Современная биология на базе представлений эмбриологии, молекулярной биологии и генетики считает, что индивидуальное развитие от одной клетки до многоклеточного зрелого организма результат последовательного, избирательного включения в работу разных генных участков хромосом в различных клетках. Это приводит к появлению клеток со специфическими для них структурами и особыми функциями, то есть к процессу, называемому дифференциацией .
Дифференциация это возникновение из однородных клеток в течение индивидуального развития большого разнообразия клеточных форм, отличающихся по строению и функциям. Проявляющиеся в процессе дифференциации различия сохраняются клетками при размножении, то есть оказываются наследственно закрепленными (например, клетки печени при размножении дают только клетки печени, а мышечные клетки только мышечные и т.д.).
Наиболее отчетливым признаком цитодифференциации является развитие цитоплазматических структур, связанных с функцией клеток и обусловливающих их специализацию (то есть органоидов специального назначения). Например, в клетках мышечной ткани образуются миофибриллы, которые и обеспечивают функцию сокращения. В клетках кожного эпителия тонофибриллы, а затем поверхностные слои клеток ороговевают (белок кератогиалин превращается кератин) и отмирают. В эритроцитах синтезируется гемоглобин, затем клетки утрачивают ядра, а зрелые эритроциты после длительного периода функционирования погибают и заменяются новыми.
Все эти примеры указывают на конечные признаки дифференциации. Начальные же этапы проявления этих признаков удается обнаружить далеко не всегда, и состоят они в синтезе новых, ранее отсутствовавших в клетке белков. Например, специфические мышечные белки (актин и миозин) синтезируются в одноядерных клетках, которые затем сливаются, образуя симпласт, и уже в нем обнаруживаются миофибриллы. Даже используя электронный микроскоп, выявить момент начала синтеза новых белков удается не всегда.
В настоящее время доказано, что никогда в ядре не функционирует весь геном. Дифференцировка это результат избирательной активности разных генов в клетках по мере развития многоклеточного организма.
Следовательно, можно утверждать, что любая клетка многоклеточного организма обладает одинаковым полным фондом генетического материала, всеми возможностями для проявления этого материала, но в разных клетках одни и те же гены могут находиться или в активном, или в репрессированном состоянии.
Это представление базируется на большом экспериментальном материале. Доказано, что целостное растение может быть получено из одной его соматической клетки. Этот метод получил название клонирование организмов . Опыты по клонированию животных первоначально проводились на примере земноводных: ядро зиготы у лягушек разрушали ультра-фиолетовыми лучами, на его место внедряли ядро из клетки кишечника, и в результате получали новый организм, абсолютно идентичный материнскому. Чем выше уровень организации организмов, тем труднее осуществить их клонирование. У млекопитающих этот процесс находится в стадии активного изучения, проводятся успешные опыты на мышах, на некоторых сельскохозяйственных животных.
Из этого вытекает, что клетки многоклеточных организмов обладают полным набором генетической информации, свойственной для данного организма, и в этом отношении они равнозначны. В этом состоит правило генетической тождественности клеток в пределах организма .
Но, как и в любом правиле, в нем имеются исключения: иногда при дифференцировке происходит количественное изменение генетического материала. Так, при дроблении яиц аскариды клетки, дающие начало соматическим тканям, теряют часть хромосомного материала, т.е. происходит деминуция: вместо 40 хромосом остается всего 8 хромосом. Сходный процесс описан у насекомых-галлиц (отр. Двукрылые), у которых число хромосом при деминуции уменьшается вдвое (с 32-х до 16-ти).
Эти примеры наглядно иллюстрируют роль цитоплазмы при дифференциации клеток. Если в случае с аскаридой предварительно отцентрифугировать яйцеклетки, то все компоненты цитоплазмы перемешиваются и при первом делении попадают в оба бластомера. При этом деминуции хромосом не происходит, то есть исчезает ядерная дифференциация.
У насекомых-галлиц деминуция происходит во всех ядрах, кроме одного, которое попадает в собранную у нижнего полюса зиготы плазму, богатую РНК. При облучении зародышевой плазмы ультрафиолетовыми лучами происходит разрушение РНК, при этом ядро претерпевает деминуцию вместе с другими ядрами зародыша, и развивается нормальное насекомое, но только стерильное, так как половые клетки не формируются.
Однако, первостепенную роль в дифференциации играет ядро. Роль ядра в дифференциации клеток можно показать на двух примерах.
I . Гигантская морская одноклеточная водоросль ацетабулярия имеет сложное строение. Она состоит из ризоида, в котором помещается ядро, стебелька до 5 см длиной и шапочки. Есть два вида ацетабулярии, которые отличаются формой шапочки: у первого вида длинный стебелек и шапочка в виде блюдца; у другого вида короткий стебелек и розетковидная шапочка.
На ризоид второго вида был пересажен стебелек с шапочкой первого вида. Через некоторое время шапочка удалялась и регенерировала шапочка розетковидной формы, т.е. признаки ее определялись ядром.
II . Опыты Б.Л. Астаурова над тутовым шелкопрядом.
Облучая яйцеклетки большими дозами рентгеновских лучей и активируя их после оплодотворения температурным воздействием, удалось не только разрушить ядро яйцеклетки, но и индуцировать андрогенез, то есть развитие особей за счет слияния 2-х ядер сперматозоидов (для тутового шелкопряда характерна полиспермия). В результате развивались личинки, обладавшие только отцовскими признаками.
Из этих опытов, поставленных на совершенно различных организмах, следует, что общие признаки организма, в том числе и видовые, определяются ядром, и ядро содержит всю необходимую информацию, обеспечивающую развитие организма.
В общей форме, вероятно, наиболее приемлема теория Т. Моргана, согласно которой сначала ядро воздействует на цитоплазму и программирует белковый синтез, а затем цитоплазма влияет на ядро, избирательно блокируя ряд генов, которые до этого функционировали. Цитоплазма, получившая определенную информацию, репрессирует все гены, которые не должны работать в данный момент.
Второй системой (помимо генов), обеспечивающей правильное развитие организма и дифференциацию его клеток, являются индуцирующие механизмы (воздействие внешних факторов) и, прежде всего, эмбриональная индукция.
Эмбриональная индукция это взаимодействие между частями развивающегося организма у многоклеточных беспозвоночных и всех хордовых, в процессе которого одна часть индуктор, приходя в контакт с другой частью реагирующей системой , определяет направление развития последней.
Эмбриональная индукция открыта в 1901 г. Х. Шпеманом на примере развития зародыша земноводных. Он установил, что для образования у этих животных нервной пластинки из эктодермы гаструлы необходим контакт эктодермы с хордомезодермальным зачатком. Клетки этого зачатка выделяют химические вещества, которые диффундируют в клетки эктодермы и заставляют их превращаться в нервные клетки. Вопрос о химической природе индуктора окончательно не решен до сих пор. Скорее всего, это могут быть белки, РНК, рибонуклеопротеиды и т.п.
Для осуществления эмбриональной индукции необходимо:
1) чтобы клетки реагирующей системы обладали компетенцией, то есть способностью реагировать на индуктор; она сохраняется только на некоторое время;
2) индуктор должен выделяться в определенное время и распространяться на определенный участок реагирующей системы;
3) действие индуктора должно продолжаться какое-то минимальное время, чтобы реагирующая система успела отреагировать.
Действие индукторов лишено видовой специфичности, т.е. действие собственных индукторов может быть заменено в эксперименте чужеродными, при этом результат будет тот же. Например, один из индукторов белкового характера, выделенный из куриных зародышей, вызывает аналогичные изменения и в зародыше земноводных.
Старение и смерть клетки
Наиболее подходящим объектом для изучения процессов старения на клеточном уровне являются клетки, утратившие способность к делению еще в эмбриональном периоде развития организма. К такому типу клеток относятся клетки нервной системы, скелетных мышц, миокарда. Продолжительность жизни этих клеток равна продолжительности жизни организма.
При сравнении клеток молодого организма с гомологичными клетками организмов более старшего возраста обнаруживается ряд изменений, которые с основанием могут считаться признаками старения. Для удобства изучения эти признаки можно разделить на несколько групп.
I . Морфологические признаки:
1) кариопикноз , то есть уменьшение ядра в объеме и его уплотнение;
2) стирание границ между клетками;
3) вакуолизация цитоплазмы;
4) увеличение количества амитозов.
II . Физико-химические признаки:
1) уменьшение степени дисперсности коллоидов цитоплазмы и ядра;
2) увеличение вязкости цитоплазмы и кариоплазмы;
3) более легкая коагуляция внутриклеточных белков при действии на них спирта, растворов солей.
III . Биохимические признаки:
1) накопление в цитоплазме оранжево-желтого пигмента липофу-сцина (это продукт окисления ненасыщенных липидов);
2) уменьшение содержания воды в клетке;
3) снижение активности ферментов;
4) увеличение содержания холестерина;
5) уменьшение содержания белка лецитина.
IV . Функциональные признаки:
1) понижается интенсивность внутриклеточного дыхания;
2) угнетается биосинтез белка;
3) увеличивается устойчивость клеток к действию различных пов-реждающих агентов.
Смерть клетки наступает в результате действия повреждающих факторов, при старении, а также в результате накопления в цитоплазме специализированных продуктов синтеза, как это наблюдается у клеток голокриновых желез.
В некоторых случаях переход клетки от жизни к смерти происходит очень быстро, (например, при действии повреждающих факторов высокой интенсивности). Тогда структурные и метаболические изменения клетки произойти не успевают, и клетка сохраняет почти в неизменном виде свою структуру. Если же процесс умирания затягивается, наблюдается ряд изменений, которые называются некротическими:
1) происходит угнетение функций митохондрий, нарушение окислительного фосфорилирования и активация гликолиза;
2) наблюдается нарушение гомеостатических свойств клетки, т.е. рН сдвигается в кислую сторону, соли, метаболиты освобождаются и переходят из клетки в окружающую среду;
3) в результате подкисления и изменения электролитного состава клетки происходит денатурация внутриклеточных белков;
4) вследствие выше перечисленных процессов разрушаются мембраны лизосом, освобождаются гидролитические ферменты, которые начинают свою разрушительную работу; они вызывают гидролиз белков, углеводов, жиров, ДНК и разрушают внутриклеточные структуры;
5) ядро умирающей клетки распадается на отдельные фрагменты ( кариорексис ), которые затем растворяются ( кариолизис ).
Гибель организма, как правило, происходит в результате смерти некоторой небольшой группы жизненно важных клеток, и после смерти организма многие его клетки остаются еще живыми и функционально полноценными.
Нарушения дифференциации клеток, ведущие
к патологическим изменениям. Злокачественный рост
Как отдельные клетки, так и целые многоклеточные организмы могут подвергаться различным воздействиям, которые приводят к их структурно-функциональным изменениям, к нарушениям их жизненных функций, т.е. к патологии.
Изучение различных патологических изменений клетки имеет большое прикладное значение, так как прямо связано с задачами медицины. Кроме того, изучение типов клеточного поражения, процессов их развития, способности клеток к репаративным процессам имеет большое общебиологическое значение, раскрывая пути взаимосвязи и регуляции между отдельными клеточными компонентами. Современная биология рассматривает клетку как единую, комплексную интегрированную систему, где отдельные функции взаимосвязаны и сбалансированы друг с другом.
Таким образом, первичное нарушение любой общеклеточной функции непременно вызовет цепь взаимосвязанных внутриклеточных событий. Это можно показать на следующем примере. Под действием алкоголя происходит набухание митохондрий и нарушение их функций, вследствие этого наблюдается недостаток АТФ и затухание синтеза белков. Из-за недостатка ферментов и структурных белков происходит падение синтеза РНК и ДНК, нарушение проницаемости мембран. Это влечет за собой набухание клетки, а затем гибель органоидов и клетки в целом.
В зависимости от интенсивности поражения, его длительности и характера, судьба клетки может быть различна. Такие измененные клетки:
1) или адаптируются, приспосабливаются к повреждающему фактору;
2) или могут репарировать повреждения и реактивироваться после снятия повреждающего воздействия;
3) или могут измениться необратимо и погибнуть.
Опухолевые клетки характеризуются следующими свойствами:
1. Безудержность, неограниченность размножения. У них практически отсутствует ограничение числа делений, в то время как нормальные клетки ограничены в своих делениях. Скорость самого процесса деления опухолевых клеток равна скорости митоза нормальных клеток, сокращается продолжительность интерфазы.
3. Относительная автономность от регуляторных влияний со стороны организма. Эта особенность заключается в том, что опухолевые клетки не подчиняются регуляторным влияниям всего организма. В здоровом организме это влияние осуществляется на разных уровнях: межклеточном, межтканевом, гормональном, нервном. Степень опухолевой автономности может быть различна для разных опухолей. Так, рост некоторых опухолей может контролироваться со стороны эндокринной системы организма, другие опухоли растут вне зависимости от нее.
4. Способность к метастазированию. Вышеописанная автономизация опухолевых клеток позволяет им жить практически в любых участках организма. Отдельные опухолевые клетки могут с помощью тока крови или лимфы быть перенесены на новые места, там начать размножаться, давать новую колонию клеток, то есть метастазы. В этом отношении опухолевые клетки используют организм как какой-то субстрат, необходимый им для размножения и роста.
Все эти свойства клетки сохраняют из поколения к поколению, то есть свойства злокачественности являются наследственной особенностью таких клеток. Поэтому раковые клетки часто сравнивают с мутантами клетками, обладающими измененной генетической структурой. Возникновение раковой мутации объясняют по-разному.
Одни исследователи считают, что в результате мутации клетка утрачивает какие-то факторы (например, гены-регуляторы), необходимые для дифференцировки.
По другим представлениям, эти факторы не потеряны, а блокированы либо какими-то веществами, либо вирусами, материал которых остается в клетках в скрытом виде в течение многих клеточных поколений.
В любом случае для клетки результат будет один и тот же, независимо от того, утратит ли она те или иные гены-регуляторы, будут ли эти гены блокированы или клетка приобретает дополнительную генетическую информацию вирусной природы, в ней происходит изменение генома, соматическая мутация, выражающаяся в нарушении дифференцировки клетки и приобретении ею свойств злокачественности.
Клетка – элементарная единица строения и жизнедеятельности всех живых организмов (кроме вирусов), обладающая собственным обменом веществ, способная к самостоятельному существованию, самовоспроизведению и развитию. (Наука – цитология.)
Клетки могут существовать как одноклеточные организмы, так и в составе многоклеточных организмов.
Клеточная теория – одно из общепризнанных биологических обобщений, утверждающих единство принципа строения и развития мира растений и животных и остальных живых организмов с клеточным строением, в которой клетка рассматривается в качестве общего структурного элемента живых организмов.
Все организмы растений и животных состоят из клеток.
Каждая клетка функционирует независимо от других, но вместе со всеми.
Все клетки возникают из бесструктурного вещества неживой материи.
Позднее Р. Вирхов (1858) внес уточнение:
Все клетки возникают только из клеток путем их деления.
Современная клеточная теория содержит следующие положения:
1. Клеточная организация возникла на заре жизни и прошла долгий путь эволюции от безъядерных (прокариот) к ядерным (эукариотам), от предклеточных организмов к одно- и многоклеточным.
2. Новые клетки образуются путем деления ранее существующих.
3. Клетка является микроскопической живой системой, состоящей из цитоплазмы и ядра, окруженных мембраной (за исключением прокариот).
4. В клетке осуществляется:
Б) обратимые физиологические процессы – дыхание, раздражимость, движение
В) необратимые процессы – рост и развитие.
5. Клетка может быть самостоятельным организмом (прокариоты и простейшие, одноклеточные водоросли и грибы). Все многоклеточные организмы также состоят из клеток и их производных. Рост, развитие и размножение многоклеточного организма – следствие жизнедеятельности одной (зигота) или нескольких клеток (культура тканей).
Эти положения доказывают единство происхождения всех живых организмов, единство всего органического мира. Благодаря клеточной теории стало понятно, что клетка - это важнейшая составляющая часть всех живых организмов.
Типы клеточной организации: Выделяют два типа клеточной организации: 1) прокариотический, 2) эукариотический. Общим для клеток обоих типов является то, что клетки ограничены оболочкой, внутреннее содержимое представлено цитоплазмой. В цитоплазме находятся органоиды и включения. Органоиды — постоянные, обязательно присутствующие, компоненты клетки, выполняющие специфические функции. Органоиды могут быть ограничены одной или двумя мембранами (мембранные органоиды) или не ограничены мембранами (немембранные органоиды). Включения — непостоянные компоненты клетки, представляющие собой отложения веществ, временно выведенных из обмена или конечных его продуктов.
Эукариотические клетки по определению и в отличие от прокариотических имеют ядро. Ообычный линейный размер клеток прокариот 1-10 мкм, эукариот –10-100 мкм. Метаболизм прокариот как анаэробный, так и аэробный, эукариот – аэробный. Органоиды у прокариот немногочисленны или отсутствуют, мембранных органоидов - нет. ДНК прокариот кольцевая, располагается в цитоплазме, ДНК эукариот – линейная, организована в хромосомы, расположена в ядре. Цитоплазма прокариот не имеет цитоскелета, движения цитоплазмы, экзо- и эндоцитоза. Деление прокариотической клетки – бинарное, пополам, эукариотической – митоз или мейоз. Наконец, прокариотические организмы - одноклеточные, а эукариотические - преимущественно многоклеточные, с клеточной дифференциацией.
Специализация клеток многоклеточного организма - это клетки которые объединены в различные органы и ткани, специализирующиеся на выполнении определённых функций.
Интеграция клеток - это образование клеточных комплексов, в пределах которых клетки специализируются на выполнении узко ограниченной работы и действуют как единое целое, как единая система.


Поток энергии в клетке
В развитии низших и высших позвоночных отчетливо прослеживается единая общебиологическая закономерность, выражающаяся в появлении зародышевых листков и обособлении основных зачатков органов и тканей. Процесс образования тканей из материала эмбриональных зачатков составляет суть учения о гистогенезах.
Эмбриональный гистогенез, по определению А.А. Клишова (1984), — это комплекс координированных во времени и пространстве процессов пролиферации, клеточного роста, миграции, межклеточных взаимодействий, дифференциации, детерминации, программированной гибели клеток и некоторых других. Все названные процессы в той или иной мере протекают в зародыше, начиная с самых ранних стадий его развития.
Пролиферация. Основной способ деления тканевых клеток — это митоз. По мере увеличения числа клеток возникают клеточные группы, или популяции, объединенные общностью локализации в составе зародышевых листков (эмбриональных зачатков) и обладающие сходными гистогенетическими потенциями. Клеточный цикл регулируется многочисленными вне- и внутриклеточными механизмами. К внеклеточным относятся влияния на клетку цитокинов, факторов роста, гормональных и нейрогенных стимулов. Роль внутриклеточных регуляторов играют специфические белки цитоплазмы. В течение каждого клеточного цикла существуют несколько критических точек, соответствующих переходу клетки из одного периода цикла в другой. При нарушении внутренней системы контроля клетка под влиянием собственных факторов регуляции элиминируется апоптозом, либо на некоторое время задерживается в одном из периодов цикла.
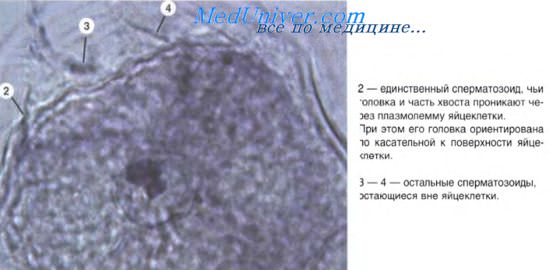
Метод радиографического анализа клеточных циклов в различных тканях выявил особенности соотношения клеточной репродукции и дифференцировки. Например, если в тканях (кроветворные ткани, эпидермис) имеется постоянный фонд пролиферирующих клеток, за счет которых обеспечивается непрерывное возникновение новых клеток взамен погибающих, то эти ткани относятся к обновляющимся. Другие ткани, например, некоторые соединительные, характеризуются тем, что в них увеличение количества клеток происходит параллельно с их дифференцировкой, клетки в этих тканях характеризуются низкой митотической активностью. Это растущие ткани. Наконец, нервная ткань характеризуется тем, что все основные процессы репродукции заканчиваются в период эмбрионального гистогенеза (когда формируется основной запас стволовых клеток, достаточный для последующего развития ткани). Поэтому она отнесена к стабильным (стационарным) тканям. Продолжительность жизни клеток в обновляющихся, растущих и стабильных тканях разная.
Наряду с обновлением клеточной популяции, в самих клетках постоянно наблюдается обновление внутриклеточных структур (внутриклеточная физиологическая регенерация).
Клеточный рост, миграция и межклеточные взаимодействия. Рост клеток проявляется в изменении их размеров и формы. При усилении функциональной активности и внутриклеточных биосинтезов наблюдается увеличение объема клетки. Если объем клетки превышает некую норму, то говорят о ее гипертрофии, и наоборот, при снижении функциональной активности происходит уменьшение объема клетки, а при переходе некоторых нормативных параметров возникает атрофия клетки. Рост клетки не беспределен и определяется оптимальным ядерно-цитоплазменным отношением.
Важное значение для гистогенеза имеют процессы перемещения клеток. Миграция клеток наиболее характерна для периода гаструляции. Однако и в период гисто- и органогенеза происходят перемещения клеточных масс (например, смещения миобластов из миотомов в места закладки скелетных мышц; движение клеток из нервного гребня с образованием спинномозговых ганглиев и нервных сплетений, миграция гоноцитов и т. д.). Миграция осуществляется с помощью нескольких механизмов. Так, различают хемотаксис — движение клеток в направлении градиента концентрации какого-либо химического агента (перемещения спермиев к яйцеклетке, предшественников Т-лимфоцитов из костного мозга в закладку тимуса).
Гаптотаксис — механизм перемещения клеток по градиенту концентрации адгезионной молекулы (движение клеток протока пронефроса у амфибий по градиенту щелочной фосфатазы на поверхности мезодермы). Контактное ориентирование — когда в какой-либо преграде остается один канал для перемещения (описан у рыб при образовании плавников).
Контактное ингибирование — этот способ перемещения наблюдается у клеток нервного гребня. Суть способа заключается в том, что при образовании ламеллоподии одной клеткой и контакта ее с другой клеткой, ламеллоподия прекращает рост и постепенно исчезает, но в другой части мигрирующей клетки при этом формируется новая ламеллоподия.
В процессе миграции клеток важную роль играют межклеточные взаимодействия. Существует несколько механизмов такого взаимодействия (контактного и дистантного). Выделяется большая группа молекул клеточной адгезии (МКА). Так, кадгерины — это Са2+-зависимые МКА, отвечают за межклеточные контакты при образовании тканей, за формообразование и др. В молекуле кадгерина различают внеклеточный, трансмембранный и внутриклеточный домены. Например, внеклеточный домен ответственен за адгезию клеток с одинаковыми кадгеринами, а внутриклеточный — за форму клетки. Другой класс МКА — это иммуноглобулиновое суперсемейство Са2+-независимых МКА, обеспечивающих, например, адгезию аксонов к сарколемме мышечных волокон, или миграцию нейробластов вдоль радиальных глиоцитов в закладке коры большого мозга и др. Следующий класс МКА — это мембранные ферменты — гликозилтранферазы. Последние по типу "ключ-замок" соединяются с углеводными субстратами — гликозаминогликанами надмембранного комплекса клетки, осуществляя таким образом прочное сцепление клеток.
Кроме механизмов межклеточного взаимодействия, существуют механизмы взаимодействия клеток с субстратом. Они включают формирование рецепторов клетки к молекулам внеклеточного матрикса. К последним относят производные клеток, среди которых наиболее изученными адгезионными молекулами являются коллаген, фибронектин, ламинин, тенасцин и некоторые другие. Коллагены, среди которых различают несколько десятков типов, входят в состав межклеточного вещества рыхлой волокнистой соединительной ткани, базальной мембраны и пр. Фибронектин, секретируемый клетками, является связывающей молекулой между мигрирующей клеткой и межклеточным матриксом. Ламинин — компонент базальной мембраны, также связывает мигрирующие клетки с межклеточным матриксом (справедливо по отношению к эпителиоцитам и нейробластам).
Для осуществления связи мигрирующих клеток с межклеточным матриксом клетки формируют специфические рецепторы. К ним относятся, например, синдекан, который обеспечивает контакт эпителиоцита с базальной мембраной за счет сцепления с молекулами фибронектина и коллагена. Интегрины клеточных поверхностей связывают с внеклеточной стороны молекулы внеклеточного матрикса, а внутри клетки — белки цитоскелета (например, актиновые микрофиламенты). Так возникает связь внутри- и внеклеточных структур, что позволяет клетке использовать для перемещения собственный сократительный аппарат. Наконец, существует большая группа молекул, формирующих клеточные контакты, осуществляющие коммуникацию между клетками (щелевые контакты), механическую связь (десмосомы, плотные контакты).
Дистантные межклеточные взаимодействия осуществляются путем секреции гормонов и факторов роста (ФР). Последние — это вещества, оказывающие стимулирующее влияние на пролиферацию и дифференцировку клеток и тканей. К ним относятся, например, ФР, полученный из тромбоцитов и влияющий на переход клеток в фазу размножения (гладких миоцитов, фибробластов, глиоцитов); эпидермальный ФР — стимулирует пролиферацию эпителиоцитов, производных эктодермы; ФР фибробластов — стимулирует пролиферацию фибробластов. Особо выделяется большая группа пептидов (соматотропины, соматомедины, инсулин, лактоген), влияющих на развитие клеток плода.
Факторы, тормозящие пролиферацию и дифференцировку клеток, также принимают кооперативное участие в процессах развития клеток и тканей. К ним относятся, например, бета-интерферон и трансформирующий ФР. Последний, однако, в отношении разных клеточных типов действует по-разному: блокирует размножение многих типов эпителиоцитов, но стимулирует размножение соединительнотканных клеток.
- Вернуться в оглавление раздела "гистология"
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Читайте также: