
Характеристика днк полимераз эукариот реферат
Обновлено: 07.07.2024
В полимеразы представляют собой ферменты, функция которых связана с процессами репликации и транскрипции нуклеиновых кислот. Есть два основных типа этих ферментов: ДНК-полимераза и РНК-полимераза.
ДНК-полимераза отвечает за синтез новой цепи ДНК в процессе репликации, добавляя новые нуклеотиды. Это большие сложные ферменты, различающиеся по структуре в зависимости от того, обнаружены они в эукариотическом или прокариотическом организме.
Точно так же РНК-полимераза действует во время транскрипции ДНК, синтезируя молекулу РНК. Подобно ДНК-полимеразе, он обнаружен как у эукариот, так и у прокариот, и его структура и сложность варьируются в зависимости от группы.
С эволюционной точки зрения можно предположить, что первые ферменты должны были обладать полимеразной активностью, поскольку одним из внутренних требований для развития жизни является способность генома к репликации.
Центральная догма молекулярной биологии
В этой статье мы рассмотрим два важных фермента, участвующих в первых двух упомянутых процессах.
Стоит отметить, что из центральной догмы есть исключения. Многие гены не транслируются в белки, и в некоторых случаях поток информации идет от РНК к ДНК (как в ретровирусах).
ДНК-полимераза
Характеристики
ДНК-полимераза - это фермент, отвечающий за точную репликацию генома. Работа фермента должна быть достаточно эффективной, чтобы обеспечивать сохранение генетической информации и ее передачу следующим поколениям.
Если учесть размер генома, это довольно сложная задача. Например, если мы поставим себе задачу переписать 100-страничный документ на нашем компьютере, у нас обязательно будет одна ошибка (или больше, в зависимости от нашей концентрации) на каждой странице.
Полимераза может добавлять более 700 нуклеотидов каждую секунду, а неверно только каждые 10. 9 или 10 10 включенных нуклеотидов, необычайное количество.
Полимераза должна иметь механизмы, позволяющие точно копировать информацию генома. Следовательно, существуют разные полимеразы, которые обладают способностью реплицировать и восстанавливать ДНК.
Характеристики и состав
ДНК-полимераза - это фермент, который работает в направлении 5'-3 'и работает путем добавления нуклеотидов к концевому концу со свободной группой -ОН.
Одним из непосредственных следствий этой характеристики является то, что одну из цепей можно синтезировать без каких-либо неудобств, но как насчет цепи, которую необходимо синтезировать в направлении 3'-5 '?
Эта цепь синтезируется в так называемые фрагменты Окадзаки. Таким образом, небольшие сегменты синтезируются в нормальном направлении, 5'-3 ', которые впоследствии соединяются ферментом, называемым лигазой.
Структурно ДНК-полимеразы имеют два общих активных центра, которые содержат ионы металлов. В них мы находим аспартат и другие аминокислотные остатки, которые координируют металлы.
Типы
Традиционно у прокариот выделяют три типа полимераз, названные римскими цифрами: I, II и III. У эукариот распознаются пять ферментов, которые названы буквами греческого алфавита, а именно: α, β, γ, δ и ε.
Последнее исследование выявило пять типов ДНК в Кишечная палочка, 8 в дрожжах Saccharomyces cerevisiae и более 15 у человека. В растительной линии фермент изучен меньше. Однако в модельном организме Arabidopsis thaliana Описано около 12 ферментов.
Приложения
Одним из наиболее часто используемых методов в лабораториях молекулярной биологии является ПЦР или полимеразная цепная реакция. Эта процедура использует полимеризационную способность ДНК-полимеразы для амплификации на несколько порядков молекулы ДНК, которую мы хотим изучить.
Другими словами, в конце процедуры у нас будут тысячи копий нашей целевой ДНК.Использование ПЦР очень разнообразно. Его можно применить в научных исследованиях, для диагностики некоторых заболеваний или даже в экологии.
РНК-полимераза
Характеристики
РНК-полимераза отвечает за создание молекулы РНК, начиная с матрицы ДНК. Полученный транскрипт представляет собой копию, которая дополняет сегмент ДНК, который использовался в качестве матрицы.
Информационная РНК отвечает за перенос информации в рибосому, чтобы генерировать белок. Они также участвуют в синтезе других типов РНК.
Он не может действовать в одиночку, ему нужны белки, называемые факторами транскрипции, чтобы успешно выполнять свои функции.
Характеристики и состав
РНК-полимеразы - это большие ферментные комплексы. Они более сложны в эукариотической линии, чем в прокариотической.
У эукариот есть три типа полимераз: Pol I, II и III, которые являются центральным механизмом для синтеза рибосомной, информационной и транспортной РНК соответственно. Напротив, у прокариот все их гены процессируются одним типом полимеразы.
Различия между ДНК и РНК-полимеразой
Хотя оба фермента используют отжиг ДНК, они различаются по трем ключевым причинам. Во-первых, ДНК-полимераза требует первый для начала репликации и подключения нуклеотидов. А первый или праймер представляет собой молекулу, состоящую из нескольких нуклеотидов, последовательность которых комплементарна определенному участку в ДНК.
Праймер отдает полимеразе свободный –ОН, чтобы запустить ее каталитический процесс. Напротив, РНК-полимеразы могут начать свою работу без необходимости первый.
Во-вторых, ДНК-полимераза имеет несколько участков связывания на молекуле ДНК. РНК-полимераза может связываться только с промоторными последовательностями генов.
Наконец, ДНК-полимераза - это фермент, который выполняет свою работу с высокой точностью. РНК-полимераза подвержена большему количеству ошибок, вводя неправильный нуклеотид каждые 10 4 нуклеотиды.
Механизмы репликации ДНК у эукариот менее изучены из-за их большей сложности. Основные результаты получены на модельной системе с ДНК вируса SV40, в которой процесс репликации исследовали в зараженных клетках человека, культивируемых in vitro. В этой системе вирусный белок, называемый Т-антигеном, выполняет многие функции, необходимые для репликации вирусной ДНК. Он является белком-инициатором, обладает ДНК-хеликазной активностью и необходим для правильного взаимодействия с ДНК ферментного комплекса, синтезирующего праймеры. В то же время вирус SV40 использует для репликации своей небольшой хромосомы и многие белки клетки-хозяина, что позволяет исследовать функционирование репликативного комплекса клеток человека в такой относительно простой системе.
ДНК-полимеразы эукариот. В клетках эукариот имеются по меньшей мере шесть различных ДНК-зависимых ДНК-полимераз: α, β, δ,ε,γ,ζ. Четыре из них — α, β, δ,ε — непосредственно участвуют в репликации хромосомной ДНК (табл. 18).
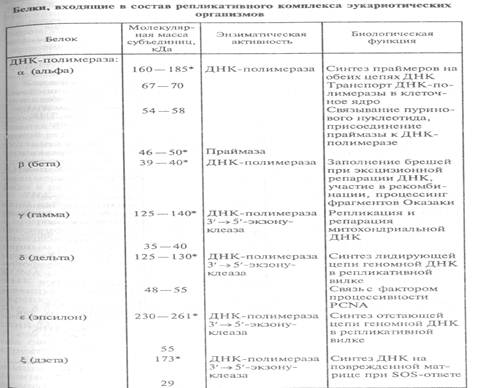
ДНК-полимераза α —первая ДНК-полимераза, обнаруженная в клетках эукариот. Она представлена в клетке в виде прочного комплекса с ДНК-праймазой — ферментом, осуществляющим синтез РНК-затравок. Комплекс ДНК-полимераза α-праймаза является единственным у эукариот ферментативным ансамблем, способным инициировать синтез ДНК de novo. В ходе репликации в клеточных ядрах ДНК полимераза α-праймаза синтезирует затравку лидирующей нити в участке ori и затравки фрагментов Оказаки запаздывающей нити. Как правило, ДНК-полимераза не обладает корректорской 3'→5'-экзонуклеазной активностью. По-видимому, в ходе эволюции экзонуклеазный центр в данном ферменте редуцировался.
ДНК-полимеразаβ является наименьшей по размеру и самой простой по строению ДНК-полимеразой в клетках эукариот. Основная функция ДНК-пол.β в клетке связана с эксцизионной репарацией ядерной ДНК (заполнение пробелов при репарации).
ДНК-полимераза ε,выделена из клеток HeLa, содержит два полипептида — каталитический 261 кДа и полипептид 55 кДа. Каталитический полипептид обладает ДНК-полимеразной и 3'→5‘-экзонуклеазной активностями. Особенностью холофермента ДНК-полимеразы ε по сравнению с ДНК-полимеразой δ является его меньшая зависимость от вспомогательных факторов (PCNA, RFС - репликативный фактор С и RPA – репликативный ядерный белок А), а также низкая (почти на порядок) скорость синтеза ДНК. Это различие, возможно, связано с разной функцией ДНК-полимераз в репликативной вилке. Один холофермент, ДНК-полимераза δ осуществляет быстрый и процессивный синтез лидирующей нити, используя для элонгации единственную
ДНК-полимераза γ локализована в митохондриях, ее функция связана с репликацией и репарацией митохондриальной ДНК, она кодируется ядерным геномом. ДНК-полимераза γ способна направлять высокопроцессивную полимеризацию на однонитевых ДНК-матрицах в отсутствие вспомогательных факторов.
Охарактеризованы также другие ДНК-полимеразы эукариот: ή, θ, REV1 и др. Все эти ферменты участвуют в репарации ДНК.
В последние годы наряду с углубленным изучением строения и свойств отдельных ДНК-полимераз эукариот большое внимание уделяется взаимодействию этих ферментов со вспомогательными факторами и механизму функционирования их в составе многокомпонентных репликативных и репаративных комплексов. Список белков, взаимодействующих с ДНК-полимеразами, постоянно растет и включает не только известные факторы PCNA, RFC и RPA, но и ключевые факторы регуляции клеточного метаболизма, такие, как белки группы МСМ (minichromosome maintenance factors), факторы узнавания участков ori репликации ORC (origin recognition complex) и др.
Таким образом, поскольку эукариотические ДНК-полимеразы α и β лишены 3′ → 5 ′ и 5 ′ → 3 ′ -экзонуклеазных активностей, становится понятным участие ДНК-полимераз δ и ε в процессе репликации ядерной ДНК в качестве корректирующих ферментов, а ДНК-полимеразе ε приписывают также функцию удаления РНК-праймеров на концах фрагментов Оказаки.
Модель работы димерной полимеразы(ДНК-пол.III); координация синтеза ДНК на комплементарных цепях
Элонгация репликации. Модель полуконсервативной прерывистой репликации ДНК в репликативной вилке представлена на рис. Дуплекс родительской молекулы ДНК расплетает АТР- зависимая хеликаза. Образующиеся одноцепочечные участки покрывает SSB-белок. ДНК-пол.III движется по одной из матричных цепей в направлении раскрывания вилки и синтезирует ведущую цепь ДНК. По другой, матричной цепи в том же направлении движется праймосома (комплекс белков, осуществляющий инициацию репликации). Время от времени, входящая в состав праймосомы праймаза (белок Dna G), Синтезирует РНК-затравки для отстающей цепи. Вторая молекула холофермента ДНК-пол. III удлиняет эти затравки до тех пор, пока не достигнет предыдущей затравки, т.е. она синтезирует фрагменты Оказаки длиной от 1000 до 2000 н. Удаление сегментов РНК с 5'-конца каждого фрагмента Оказаки осуществляет ДНК- пол.I, используя свою 5'→3'-кзонуклеазную активность. Заполнение брешей между фрагментами Оказаки катализируется той же ДНК-полимеразой I. Одноцепочечный разрыв зашивает ДНК-лигаза. Таким образом, из серии фрагментов Оказаки образуется новая цепь.
Cпособность клеток поддерживать высокую упорядоченность своей организации зависит от генетической информации, которая сохраняется в форме дезоксирибонуклеиновой кислоты (ДНК). ДНК - это вещество, из которого состоят гены. Размножение живых организмов, передача наследственных свойств из поколения в поколение и развитие многоклеточного организма из оплодотворенной яйцеклетки возможны потому, что ДНК способна к самовоспроизведению. Сам процесс самовоспроизведения ДНК называется репликацией. Иногда используют также название-синоним - редупликация.
Матричный синтез ДНК
Как известно, генетическая информация записана в цепи ДНК в виде последовательности нуклеотидных остатков, содержащих одно из четырех гетероциклических оснований: аденин (A), гуанин (G), цитозин (C) и тимин (T). Предложенная Дж. Уотсоном и Ф. Криком в 1953 году модель строения ДНК в форме регулярной двойной спирали сразу же позволила понять принцип удвоения ДНК. Информационное содержание обеих цепей ДНК идентично, так как каждая из них содержит последовательность нуклеотидов, строго соответствующую последовательности другой цепи. Это соответствие достигается благодаря наличию водородных связей между направленными навстречу друг другу основаниями двух цепей - попарно G и C или A и T. Описывая это свойство двойной спирали, молекулярные биологи говорят, что цепи ДНК комплементарны за счет образования уотсон-криковских пар GРC и AРT. Поскольку две цепи имеют противоположную направленность, их называют антипараллельными. Легко представить, что удвоение ДНК происходит вследствие того, что цепи расходятся, а потом каждая цепь служит матрицей, на которой собирается комплементарная ей новая цепь ДНК‚ результате образуются две дочерние, двуспиральные, неотличимые по строению от родительской ДНК молекулы. Каждая из них состоит из одной цепи исходной родительской молекулы ДНК и одной вновь синтезированной цепи. Такой механизм репликации ДНК, при котором от одного поколения к другому передается одна из двух цепей, составляющих родительскую молекулу ДНК, получил название полуконсервативного и был экспериментально доказан в 1958 году М. Мезельсоном и Ф. Сталь.
Кроме того, ситезу ДНК характерны такие свойства, как антипараллельность и униполярность. Каждая цепь ДНК имеет определенную ориентацию. Один конец несет гидроксильную группу (ОН), присоединенную к 3'-углероду в сахаре дезоксирибозе, на другом конце цепи находится остаток фосфорной кислоты в 5'-положении сахара. Две комплементарные цепи в молекуле ДНК ориентированы в противоположных направлениях - антипараллельно (при параллельной ориентации напротив 3'-конца одной цепи находился бы 3'-конец другой). Ферменты, синтезирующие новые нити ДНК, называемые ДНК-полимеразами, могут передвигаться вдоль матричных цепей лишь в одном направлении - от их 3'-концов к 5'-концам. Џри этом синтез комплементарных нитей всегда ведется в 5' 3' направлении, то есть униполярно. Поэтому в процессе репликации одновременный синтез новых цепей идет антипараллельно.
ДНК-полимеразы могут давать "задний ход", то есть двигаться в направлении 3' 5'. В том случае, когда последнее добавленное при синтезе нуклеотидное звено оказалось некомплементарным нуклеотиду матричной цепи, оно будет замещено комплементарным нуклеотидом. Отщепив "неправильный" нуклеотид, ДНК-полимераза продолжает синтез в 5' 3' направлении. Такая способность к исправлению ошибок получила название корректорской функции фермента (см. ниже).
В 1957 году А. Корнберг обнаружил у кишечной палочки фермент, катализирующий процесс полимеризации ДНК из нуклеотидов; он был назван ДНК-полимеразой. Затем ДНК-полимеразы выявили и в других организмах. Было показано, что субстратами всех этих ферментов служат дезоксирибонуклеозидтрифосфаты (дНТФ), полимеризующиеся на одноцепочечной ДНК-матрице. ДНК-полимеразы последовательно наращивают одноцепочечную цепь ДНК, шаг за шагом присоединяя к ней следующие звенья в направлении от 5-' к 3'-концу, причем выбор очередного дНТФ диктуется матрицей. Присоединение каждого нового нуклеотидного остатка к 3'-концу растущей цепи сопровождается гидролизом богатой энергией связи между первым и вторым фосфатными остатками в дНТФ и отщеплением пирофосфата, что делает реакцию в целом энергетически выгодной.
В клетках обычно присутствует несколько типов ДНК-полимераз, выполняющих различные функции и имеющих разное строение: они могут быть построены из различного количества белковых цепей (субъединиц), от одной до десятков. Однако все они работают на любых последовательностях нуклеотидов матрицы; задача этих ферментов- сделать точную копию каждой матрицы.
Точность синтеза ДНК и механизм коррекции
Генетический материал живых организмов имеет огромные размеры и реплицируется с высокой точностью. В среднем в процессе воспроизведения генома млекопитающего, состоящего из ДНК длиной 3 млрд пар нуклеотидов, возникает не более трех ошибок. При этом ДНК синтезируется чрезвычайно быстро (скорость ее полимеризации колеблется в пределах от 500 нуклеотидов/с у бактерий до 50 нуклеотидов/с у млекопитающих). Высокая точность репликации, наряду с ее высокой скоростью, обеспечивается наличием специальных механизмов, осуществляющих коррекцию, то есть устраняющих ошибки. Суть механизма коррекции заключается в том, что ДНК-полимеразы дважды проверяют соответствие каждого нуклеотида матрице: один раз перед включением его в состав растущей цепи и второй раз перед тем, как включить следующий нуклеотид. Очередная фосфодиэфирная связь синтезируется лишь в том случае, если последний (3'-концевой) нуклеотид растущей цепи ДНК образовал правильную уотсон-криковскую пару с соответствующим нуклеотидом матрицы. Если же на предыдущей стадии реакции произошло ошибочное спаривание оснований, то дальнейшая полимеризация останавливается до тех пор, пока ошибка не будет исправлена. Для этого фермент перемещается в обратном направлении и вырезает последнее добавленное звено, после чего его место может занять правильный нуклеотидпредшественник. Иными словами, многие (но не все) ДНК-полимеразы обладают, помимо 5'-3'-синтетической активности, еще и 3'-гидролизующей активностью, которая обеспечивает удаление ошибочно спаренных с матрицей нуклеотидов.
Основные принципы репликации
Основные правила, в соответствии с которыми происходит репликация, были выяснены в опытах с бактериями, однако они справедливы также и для высших организмов.
Инициация цепей ДНК
ДНК-полимеразы не могут начинать синтеза ДНК на матрице, а способны только добавлять новые дезоксирибонуклеотидные звенья к 3'-концу уже имеющейся полинуклеотидной цепи. Такую заранее образованную цепь, к которой добавляются нуклеотиды, называют затравкой. Короткую РНК- затравку синтезирует из рибонуклеозидтрифосфатов фермент, не обладающий корректирующей активностью и называемый ДНК-праймазой (от англ. primer - затравка). Праймазная активность может принадлежать либо отдельному ферменту, либо одной из субъединиц ДНК-полимеразы. Затравка, синтезированная этим неточным ферментом, не умеющим исправлять ошибки, отличается от остальной новосинтезированной цепи ДНК, поскольку состоит из рибонуклеотидов, и далее может быть удалена.
Размер рибонуклеотидной затравки невелик (менее 20 нуклеотидов) в сравнении с размером цепи ДНК, образуемой ДНК-полимеразой. Выполнившая свою функцию РНК-затравка удаляется специальным ферментом, а образованная при этом брешь заделывается ДНК-полимеразой, использующей в качестве затравки 3'-ОН-конец соседнего фрагмента Оказаки (см ниже). Удаление крайних РНК-праймеров, комплементарных 3'-концам обеих цепей линейной материнской молекулы ДНК, приводит к тому, что дочерние цепи оказываются короче на 10-20 нуклеотидов (у разных видов размер РНК-затравок различен). В этом заключается так называемая "проблема недорепликации концов линейных молекул". В случае репликации кольцевых бактериальных ДНК этой проблемы не существует, так как первые по времени образованиЯ РНК-затравки удаляются ферментом, который одновременно заполняет образующуюся брешь путем наращивания 3'-ОН-конца растущей цепи ДНК, направленной в "хвост" удаляемому праймеру. Проблема недорепликации 3'-концов линейных молекул ДНК решается эукариотическими клетками с помощью специального фермента - теломеразы.
В 1985 году он был обнаружен у равноресничной инфузории Tetrahymena thermophila, а впоследствии - в дрожжах, растениях и животных, в том числе в яичниках человека и иммортализованных(бессмертных) линиях раковых клеток HeLa. Теломераза является ДНК-полимеразой, достраивающей 3'-концы линейных молекул ДНК хромосом короткими (6-8 нуклеотидов) повторяющимися последовательностями (у позвоночных TTAGGG). Согласно номенклатуре, этот фермент называют ДНК- уклеотидилэкзотрансферазой или теломерной терминальной трансферазой. Помимо белковой части теломераза содержит РНК, выполняющую роль матрицы для наращивания ДНК повторами. Длина теломеразной РНК колеблется от 150 нуклеотидов у простейших до 1400 нуклеотидов у дрожжей, у человека - 450 нуклеотидов. Сам факт наличия в молекуле РНК последовательности, по которой идет матричный синтез куска ДНК, позволяет отнести теломеразу к своеобразной обратной транскриптазе, то есть ферменту, способному вести синтез ДНК по матрице РНК.
В результате того что после каждой репликации дочерние цепи ДНК оказываются короче материнских на размер первого РНК-праймера (10-20 нуклеотидов), образуются выступающие однонитевые 3'-концы материнских цепей. Они-то и узнаются теломеразой, которая последовательно наращивает материнские цепи (у человека на сотни повторов), используя 3'-ОН-концы их в качестве затравок, а РНК, входящую в состав фермента, в качестве матрицы. Образующиеся длинные одноцепочечные концы, в свою очередь, служат матрицами для синтеза дочерних цепей по традиционному репликативному механизму.
Постепенное укорочение ДНК хромосом во время репликации является одной из теорий "старения" клеточных колоний. Еще в 1971 году отечественный ученый А.М. Оловников в своей теории маргинотомии (от лат. marginalis -краевой, tome - сечение) предположил, что это явление лежит в основе ограниченного потенциала удвоения, наблюдаемого у нормальных соматических клеток, растущих в культуре in vitro, так называемого "лимита Хейфлика". Американский ученый Леонард Хейфлик в начале 60-х годов показал, что если для культивирования взять клетки новорожденных детей, то они могут пройти 80-90 делений, в то время как соматические клетки от 70-летних делятся только 20- 30 раз. Ограничение на число клеточных делений и называют лимитом Хейфлика.
Расплетание двойной спирали ДНК
Поскольку синтез ДНК происходит на одноцепочечной матрице, ему должно предшествовать обязательное разделение (хотя бы на время) двух цепей ДНК. Исследования, проведенные в начале 60-х годов на реплицирующихся хромосомах, выявили особую, четко ограниченную область репликации, перемещающуюся вдоль родительской спирали ДНК и характеризующуюся местным расхождением двух ее цепей. Эта активная область из-за своей Y-образной формы была названа репликационной вилкой. Именно в ней ДНК-полимеразы синтезируют дочерние молекулы ДНК. С помощью электронной микроскопии реплицирующейся ДНК удалосьустановить, что область, которая уже реплицирована, имеет вид глазка внутри нереплицировавшейся ДНК. Важно отметить, что репликационный глазок образуется только в тех местах молекулы, где находятся специфические нуклеотидные последовательности. Эти последовательности, получившие название точек начала репликации, состоят приблизительно из 300 нуклеотидов. В зависимости от того, в одном или в двух направлениях происходит репликация (а это зависит от природы организма), глазок содержит одну или две репликационные вилки. Последовательное движение репликационной вилки приводит к расширению глазка.
Двойная спираль ДНК весьма стабильна; для того чтобы она раскрылась, необходимы особые белки. Специальные ферменты ДНК-хеликазы быстро движутся по одиночной цепи ДНК, используя для перемещения энергию гидролиза ATФ. Встречая на пути участок двойной спирали, они разрывают водородные связи между основаниями, разделяют цепи и продвигают репликационную вилку. Вслед за этим с одиночными цепями ДНК связываются специальные дестабилизирующие спираль белки, которые не позволяют одиночным цепям ДНК сомкнуться. При этом они не закрывают оснований ДНК, оставляя их доступными для спаривания.
Не следует забывать, что комплементарные цепи ДНК закручены друг вокруг друга в спираль. Следовательно, для того чтобы репликационная вилка могла продвигаться вперед, вся еще не удвоенная часть ДНК должна была бы очень быстро вращаться. Эта топологическая проблема решается путем образования в спирали своего рода "шарниров", позволяющих цепям ДНК раскрутиться. Принадлежащие к особому классу белки, называемые ДНК-топоизомеразами, вносят в цепь ДНК одноили двухцепочечные разрывы, позволяющие цепям ДНК разделиться, а затем заделывают эти разрывы. Топоизомеразы участвуют также в расцеплении зацепленных двухцепочечных колец, образующихся при репликации кольцевых двунитевых ДНК. С помощью этих важных ферментов двойная спираль ДНК в клетке может принимать "недокрученную" форму с меньшим числом витков; в такой ДНК легче происходит расхождение двух цепей ДНК в репликационной вилке.
Прерывистый синтез ДНК
Легко вообразить, что репликация происходит путем непрерывного роста нуклеотида за нуклеотидом обеих новых цепей по мере перемещения репликационной вилки; при этом, поскольку две цепи в спирали ДНК антипараллельны, одна из дочерних цепей должна была бы расти в направлении 5'-3', а другая в направлении 3'-5'. В действительности, однако, оказалось, что дочерние цепи растут только в направлении 5'-3', то есть всегда удлиняется 3'-конец затравки, а матрица считывается ДНК-полимеразой в направлении 3'-5'. Это утверждение на первый взгляд кажется несовместимым с движением репликационной вилки в одном направлении, сопровождающемся одновременным считыванием двух антипараллельных нитей. Разгадка секрета заключается в том, что синтез ДНК происходит непрерывно только на одной из матричной цепей. На второй матричной цепи ДНК синтезируется сравнительно короткими фрагментами (длиной от 100до 1000 нуклеотидов, в зависимости от вида), названными по имени обнаружившего их ученого фрагментами Оказаки). Вновь образованная цепь, которая синтезируется непрерывно, называется ведущей, а другая, собираемая из фрагментов Оказаки, отстающей. Синтез каждого из этих фрагментов начинается с РНК-затравки. Через некоторое время РНК-затравки удаляются, бреши застраиваются ДНК-полимеразой и фрагменты сшиваются в одну непрерывную цепь ДНК специальным ферментом.
Кооперативное действие белков репликационной вилки.
До сих пор мы говорили об участии отдельных белков в репликации так, как будто бы они работают независимо друг от друга. Между тем в действительности большая часть этих белков объединена в крупный комплекс, который быстро движется вдоль ДНК и согласованно осуществляет процесс репликации с высокой точностью. Этот комплекс сравнивают с крошечной "швейной машиной" : "деталями" его служат отдельные белки, а источником энергии - реакция гидролиза нуклеозидтрифос фатов. Спираль расплетается ДНК-хеликазой; этому процессу помогают ДНК- топоизомераза, раскручивающая цепи ДНК, и множество молекул дестабилизирующего белка, связывающихся с обеими одиночными цепями ДНК. В области вилки действуют две ДНК-полимеразы - на ведущей и отстающей цепи. На ведущей цепи ДНК-полимераза работает непрерывно, а на отстающей фермент время от времени прерывает и вновь возобновляет свою работу, используя короткие РНК-затравки, синтезируемые ДНК-праймазой. Молекула ДНК-праймазы непосредственно связана с ДНК-хеликазой, образуя структуру, называемую праймосомой. Праймосома движется в направлении раскрывания репликационной вилки и по ходу движения синтезирует РНК-затравку для фрагментов Оказаки. В этом же направлении движется ДНК-полимераза ведущей цепи и, хотя на первый взгляд это трудно представить, ДНК-полимераза отстающей цепи. Для этого, как полагают, последня накладывает цепь ДНК, которая служит ей матрицей, саму на себя, что и обеспечивает разворот ДНК-полимеразы отстающей цепи на 180 градусов. Согласованное движение двух ДНК-полимераз обеспечивает координированную репликацию обеих нитей. Таким образом, в репликационной вилке одновременно работают около двадцати разных белков (из которых мы назвали только часть), осуществляя сложный, высокоупорядоченный и энергоемкий процесс.
Согласованность процессов репликации ДНК и клоеточного деления
Эукариотическая клетка перед каждым делением должна синтезировать копии всех своих хромосом. Репликация ДНК эукариотической хромосомы осуществляется посредством разделени хромосомы на множество отдельных репликонов. Такие репликоны активируются не все одновременно, однако клеточному делению должна предшествовать обязательная однократная репликация каждого из них. Из сказанного ясно, что по хромосоме эукариот в каждый момент времени может двигаться независимо друг от друга множество репликационных вилок. Остановка продвижения вилки происходит только при столкновении с другой вилкой, движущейся в противоположном направлении, или по достижении конца хромосомы. В результате вся ДНК хромосо мы в короткий срок оказывается реплицированной. После сборки на молекуле ДНК хромосомных белков каждая пара хромосом в процессе митоза упорядоченно разделяется по дочерним клеткам.
Процесс репликации ДНК согласован с клеточным делением и требуетсовместного действия многих белков. В нем участвуют:
1. ДНК-хеликаза и дестабилизирующие белки; они расплетают двойную спираль родительской ДНК и формируют репликационную вилку.
2. ДНК-полимеразы, которые катализируют синтез полинуклеотидной цепи ДНК в направлении 3'-5, копируя в репликационной вилке матрицу с высокой степенью точности. Поскольку две цепи двойной спирали ДНК антипараллельны, в направлении 5'-3' непрерывно синтезируется лишь одна из двух цепей, ведущая; другая цепь, отстающа, синтезируется в виде коротких фрагментов Оказаки. ДНК-полимераза способна к исправлению собственных ошибок, но не может самостоятельно начать синтез новой цепи.
3. ДНК-праймаза, которая катализирует короткие молекулы РНК-затравки. Впоследствии фрагменты РНК удаляются - их заменяет ДНК.
4.Теломераза, заканчивающая построение недорепликацированых 3'-концов линейных молекул ДНК.
5. ДНК-топоизомеразы, помогающие решить проблемы кручения и спутывания спирали ДНК.
6. Инициаторные белки, связывающиеся в точке начала репликации и способствующие образованию нового репликационного глазка с одной или двумя вилками. В каждой из вилок вслед за инициаторными белками к расплетенной ДНК сначала присоединяется белковый комплекс, состоящий из ДНК-хеликазы и ДНК-праймазы (праймосома).
Затем к праймосоме добавляются другие белки и возникает "репликационная машина", которая и осуществляет синтез ДНК.
1. О. О. Фаворова. Сохранение ДНК в ряду популяций: репликация ДНК. Соросовский образовательный журнал, 1996 г.
2. Г.М. Дымшиц. Проблема раепликации концов линейных молекул и теломераза. Соросовский образовательный журнал, 2000 г.

Прежде чем изложить современные представления о механизме биосинтеза ДНК, следует представить сведения о синтезе этого соединения в бесклеточной системе, которыми располагает биохимия. Известно, что для любого синтеза полимерной органической молекулы, осуществляемого in vitro или in vivo, требуется энергия. Источником энергии в реакциях полимеризации мононуклеотидов является энергия, освобождаемая всеми четырьмя типами дезоксирибонуклеозидтрифосфатов, участвующих в синтезе ДНК. Образующийся пирофосфат под действием пирофосфатазы также расщепляется на две молекулы ортофосфата, давая дополнительную энергию для биосинтеза ДНК.
Помимо энергии, биогенез ДНК требует наличия специфических ферментов, катализирующих отдельные этапы синтеза, и множества белковых факторов, абсолютно необходимых для регулирования процесса репликации и проявления каталитической активности ферментов.
Ферментные системы синтеза ДНК у про- и эукариот до конца не выяснены. По имеющимся данным, в репликации ДНК, включающей узнавание точки начала процесса, расплетение родительских цепей ДНК в репликационной вилке, инициацию биосинтеза дочерних цепей и дальнейшую их элонгацию и, наконец, окончание (терминация) процесса, участвует более 40 ферментов и белковых факторов, объединенных в единую ДНК-репликазную систему, называемую реплисомой.
1.Процесс биосинтеза ДНК.
Важную функцию соединения двух цепей ДНК или замыкания двух концов одной цепи ДНК в процессе репликации либо репарации ДНК выполняет особый фермент ДНК-лигаза, катализирующая за счет энергии АТФ образование фосфодиэфирной связи между 3'-ОН-группой дезоксирибозы одной цепи и 5'-фосфатной группой другой цепи ДНК.
Из клеток животных выделено несколько ДНК-полимераз, и в разных лабораториях они получили различные наименования.
К настоящему времени у эукариот, как и у бактерий (см. ранее), открыто несколько ДНК-полимераз. В репликации ДНК эукариот участвуют два главных типа полимераз – α и δ. Показано, что ДНК-полимераза α состоит из 4 субъединиц и является идентичной по структуре и свойствам во всех клетках млекопитающих, причем одна из субъединиц оказалась наделенной праймазной активностью. Самая крупная субъединица ДНК-полимеразы а (мол. масса 180000) катализирует реакцию полимеризации, преимущественно синтез отстающей цепи ДНК, являясь составной частью праймасомы. ДНК-полимераза δ состоит из 2 субъединиц и преимущественно катализирует синтез ведущей цепи ДНК (см. далее). Открыта также ДНК-полимераза ε, которая в ряде случаев заменяет δ-фермент, в частности при репарации ДНК (исправление нарушений ДНК, вызванных ошибками репликации или повреждающими агентами). Следует отметить, что в эукариотических клетках открыты два белковых фактора репликации, обозначаемых RFA и RFC. Фактор репликации А выполняет функцию белка – связывание одноцепочечной ДНК (наподобие белковых факторов связывания разъединенных цепей ДНК при репликации у Е. coli), фактор С – функцию стабилизатора всего реплика-ционного комплекса.
В генетической инженерии с целью получения белков в достаточных количествах и с заданными свойствами (например, для генотерапии наследственных и соматических болезней) широкое применение получили эндо-нуклеазы рестриктазы, катализирующие расщепление молекулы двух-цепочечной ДНК по специфическим нуклеотидным последовательностям внутри цепи. Рестриктазы узнают определенные 4–7-членные последовательности, вызывая, таким образом, разрывы в определенных сайтах цепи ДНК. При этом образуются не случайные последовательности, а фрагменты ДНК строго определенной структуры с липкими концами (ре-комбинантные ДНК), используемые далее для конструирования гибридных молекул и получения генно-инженерной, биотехнологической продукции (например, инсулина, гормона роста, интерферона, вакцин против вируса гепатита В, СПИДа и др.).
2. Общий механизм синтеза ДНК.

Химический смысл полимеризации состоит в том, что свободная 3'-гидроксильная группа матрицы атакует α-фосфатную группу соответствующего присоединяемого нуклеозидтрифосфата (определяется природой азотистого основания затравки), при этом происходят отщепление остатка пирофосфата и образование фосфодиэфирной связи. Далее свободный 3'-гидроксил вновь присоединенного нуклеотида атакует α-фосфатную группу следующего нуклеозидтрифосфата, и таким путем продолжается процесс полимеризации, идущий в направлении 5'–3', антипараллельно матрице, оканчивающейся 5'-фосфатом:

Реакция требует присутствия одноцепочечной ДНК или в крайнем случае небольшого полидезоксирибонуклеотида. В деталях выяснено значение предобразованной ДНК в механизмах действия ДНК-полимераз: ДНК служит не только затравкой, но и матрицей, на которой фермент комплементарно и антипараллельно синтезирует дочернюю цепь ДНК. Это можно представить в виде схемы:

Сложность процесса репликации ДНК объясняется тем, что обе цепи реплицируются одновременно, хотя имеют разное направление (5'–3' и 3'–5'); кроме того, рост дочерних цепей также должен происходить в противоположных направлениях. Элонгация каждой дочерней цепи может осуществляться только в направлении 5'–3'. Р. Оказаки высказал предположение, подтвержденное экспериментальными данными, что синтез одной из дочерних цепей осуществляется непрерывно в одном направлении, в то время как синтез другой дочерней цепи происходит прерывисто, путем соединения коротких фрагментов (в честь автора названы фрагментами Оказаки), в свою очередь синтезирующихся в противоположном направлении (рис. 13.4).
Как видно, синтез ведущей цепи ДНК идет всегда в направлении 5'–3', соответствующем направлению движения репликационной вилки. Сохраняя правило синтеза дочерних молекул ДНК 5'–3', синтез на второй цепи родительской ДНК идет в направлении, противоположном движению репликационной вилки. В зависимости от типа клетки фрагменты Оказаки имеют разные размеры – от нескольких сот до нескольких тысяч нуклеотидов (150–200 у эукариот и 1000–2000 у бактерий).

Рис. 13.1. Роль ДНК-полимеразы и ДНК-лигазы в синтезе кольцевой одноцепочеч-ной ДНК фага φХ174.
3. Особенности репликации ДНК у эукариот.

Рис. 13.3. Основные этапы репликации ДНК (схема).
Как было указано, инициация биосинтеза дочерних цепей ДНК требует предварительного синтеза на матрице ДНК необычного затравочного олигорибонуклеотида, названного праймером, со свободной гидроксильной группой у С-3' рибозы. Этот короткий олигорибонуклеотид синтезируется комплементарно на матрице ДНК при участии особого фермента – прай-мазы, наделенной РНК-полимеразной активностью.
Синтез начинается с реакции между 3'-ОН-группой концевого рибонуклеотида праймера и α-фосфатной группой первого дезоксирибонуклеотидтрифосфата в строгом соответствии с комплементарностью родительской цепи ДНК, при этом освобождается пирофосфат. В дальнейшем этот фрагмент РНК, комплементарно присоединенный к новообразованной цепи ДНК, разрушается под действием ДНК-полимеразы I, и возникшая брешь застраивается олигодезоксирибо-нуклеотидом при помощи той же ДНК-полимеразы I. Вполне допустимо предположение, что синтез праймера из олигорибонуклеотида имеет глубокий биологический смысл, поскольку в этом случае могут устраняться ошибки, неизбежно возникающие при инициации репликации ДНК.
4. Этапы биосинтеза ДНК.
Предложен ряд моделей механизма биосинтеза ДНК с участием указанных ранее ферментов и белковых факторов, однако детали некоторых этапов этого синтеза еще не выяснены. Основываясь главным образом на данных, полученных в опытах in vitro, предполагают, что условно механизм синтеза ДНК у Е. coli может быть подразделен на три этапа; инициацию, т.е. начало, элонгацию, т.е. продолжение, и терми-нацию, т.е. завершение (прекращение) синтеза. Каждый из этих этапов требует участия специфических ферментов и белковых факторов.
Этап I – инициация биосинтеза ДНК – является началом синтеза дочерних нуклеотидных цепей; в инициации участвует минимум восемь хорошо изученных и разных ферментов и белков. Первая фаза – это, как указано ранее, ферментативный биосинтез на матрице ДНК необычного затравочного олигорибонуклеотида (праймера) со свободной гидроксиль-ной группой у С-3' рибозы. При инициации к цепям ДНК последовательно присоединяются ДНК-раскручивающие и ДНК-связывающие белки, а затем комплексы ДНК-полимераз и праймаз (см. рис. 13.3). Инициация представляется единственной стадией репликации ДНК, которая весьма тонко и точно регулируется, однако детальные механизмы ее до сих пор не раскрыты и в настоящее время интенсивно исследуются.
Этап II – элонгация синтеза ДНК – включает два кажущихся одинаковыми, но резко различающихся по механизму синтеза лидирующей и отстающей цепей на обеих материнских цепях ДНК. Синтез лидирующей цепи начинается с синтеза праймера (при участии праймазы) у точки начала репликации, затем к праймеру присоединяются дезоксирибонуклеотиды под действием ДНК-полимеразы III; далее синтез протекает непрерывно, следуя шагу репликационной вилки. Синтез отстающей цепи, напротив, протекает в направлении, обратном движению репликационной вилки и начинается фрагментарно. Фрагменты всякий раз синтезируются раздельно, начиная с синтеза праймера, который может переноситься с готового фрагмента при помощи одного из белковых факторов репликации в точку старта биосинтеза последующего фрагмента противоположно направлению синтеза фрагментов. Элонгация завершается отделением олигорибонуклеотидных праймеров, объединением отдельных фрагментов ДНК при помощи ДНК-лигаз и формированием дочерней цепи ДНК. Нельзя исключить, однако, возможности сопряженного и согласованного механизма синтеза лидирующей и отстающей цепей ДНК при участии полимераз и всего комплекса праймасом.
Этап III – терминация синтеза ДНК – наступает, скорее всего, когда исчерпана ДНК-матрица и трансферазные реакции прекращаются. Точность репликации ДНК чрезвычайно высока, возможна одна ошибка на 1010 трансферазных реакций, однако подобная ошибка обычно легко исправляется за счет процессов репарации.
Основным свойством ДНК является то, что она служит матрицей и определяет порядок, в котором нуклеотиды выстраиваются в новые полинуклеотидные нити.
Собственно репликация ДНК в широком смысле – очень важный для делящейся клетки процесс. В него входит также подготовка хроматина к репликации и недопущение повторного митоза
Содержание
Прикрепленные файлы: 1 файл
репликацияшшш.docx
Генетическая программа всех живых организмов, за исключением РНК-содержащих вирусов, записана в нуклеотидной последовательности ДНК. Следовательно, для сохранения уникальных свойств организма необходимо точное воспроизведение этой последовательности в каждом последующем поколении. Е. соli, например, должна дуплицировать практически без ошибок полный геном размером 4·106 нуклеотидных пар при образовании каждого последующего поколения; точно так же должны быть скопированы почти 4·109 пар оснований в 23 парах хромосом человека при каждом акте деления клеток.
Основным свойством ДНК является то, что она служит матрицей и определяет порядок, в котором нуклеотиды выстраиваются в новые полинуклеотидные нити.
Собственно репликация ДНК в широком смысле – очень важный для делящейся клетки процесс. В него входит также подготовка хроматина к репликации и недопущение повторного митоза. Это обеспечивает однократную дупликацию ДНК в течение одного клеточного цикла, поддерживая таким образом стабильность генома.
1. Вилка репликации
Процесс репликации происходит в специальных структурах, названных вилками репликации. Схематическое устройство репликативной вилки E.coli представлено на рис. 1. То, что две нити молекулы ДНК расположены антипараллельно друг другу, создает ряд проблем для их олдновременной разнонаправленной репликации.
По мере движения вилки одновременно должны синтезироваться две дочерние цепи. Вилка движется в направлении от 5' к 3’ на одной цепи и от 3’ к 5' – на другой. Однако нуклеиновые кислоты синтезируются только от 5'– к 3'-концу. Проблема решается таким образом, что на одной из родительских нитей новая нить синтезируется непрерывно в направлении 5'-3', что совпадает с движением вилки репликации. Это называется лидирующей или ведущей. Другая нить называется отстающей или запаздывающей, так как синтез на ней идет с некоторой задержкой по сравнению с лидирующей нитью. Это связано с тем, что ДНК на этой нити синтезируется также от 5' к 3', но в направлении, противоположном движению вилки, и короткими фрагментами. Благодаря этому разнонаправленный синтез ДНК может осуществляться в рамках одной структуры – репликативной вилки.
Репликация лучше всего изучена у кишечной палочки E.coli.Реликация состоит из 3 стадий:
2.1 Инициация репликации у E.coli
Инициация репликации в оriС в системе in vitro начинается с формирования комплекса, в состав которого входят шесть белков: DnaА, DnaВ, DnaС, НU, Girase и SSВ. Сначала с девятичленной последовательностью связывается мономер DnaА, затем 20–40 мономеров этого белка формируют большой агрегат. ДНК ориджина опоясывает его, и цепи ДНК разъединяются в области трех тринадцатичленных последовательностей. На следующем этапе димер DnaВ/DпаС присоединяется к комплексу oriС/DnaА, формируя агрегат размером около 480 кДа, соответствующий сфере с радиусом 6 нм. В результате формируется вилка репликации.
Фермент ДНК-полимераза начинает синтез новых нитей ДНК с 3’ концов двух РНК-затравок . При таком синтезе дочерние молекулы ДНК на матричных нитях будут синтезироваться в противоположных направлениях. Элонгация начинается с присоединения одного нуклеотида к 3’ концу РНК-затравки. Это присоединение осуществляет фермент ДНК-полимераза.
К первому нуклеотиду ДНК-полимераза присоединяет второй, третий и т.д. нуклеотид в соответствии с принципом комплементарности к нуклеотидам, находящимся на матричной нити ДНК. Основания нуклеотидов новой нити и матричной соединяются друг с другом водородными связями. Поскольку синтез новой нити ДНК начинается с 3’ конца РНК-затравки, к которому присоединяется 5’ конецпервого нуклеотида ДНК, то принято считать, что синтез дочерней нити ДНК происходит в направлении 5’ - 3’.
На той нити, где синтез дочерней нити идёт в сторону вилки репликации, он идёт непрерывно, и по мере раскручивания нити фрагмент синтезированной ДНК будет постоянно удлиняться. Чем дальше продвигается вилка репликации, тем длиннее будет вновь синтезированная цепь ДНК. Эту нить ДНК называют непрерывной, лидирующей или ведущей. В дальнейшем никаких РНК-затравок на матричной нити ДНК, с которой реплицируется лидирующая нить, не формируется.
Синтез запаздывающей нити ДНК происходит не постоянно, а фрагментами. После окончания синтеза одного фрагмента вблизи репликационной вилки вновь происходит синтез РНК-затравки - С. С 3’ этой затравки вновь начинается синтез дочерней нити ДНК в направлении РНК-затравки – Б и по её достижению синтез вновь прекращается – сформировался второй фрагмент ДНК, который начинается от РНК-затравки – С. Фактически на отстающей цепи мы имеем два фрагмента, которые состоят из соединённых друг с другом РНК и ДНК. Один фрагмент ДНК начинается с РНК-затравки Б, второй – с РНК-зартавки С. Эти фрагменты ДНК носят название фрагмент Оказаки .Последующие раунды репликации повторяются – топоизомераза раскручивает очередной виток спирали ДНК, хелаза разрывает водородные связи между нитями ДНК и они расходятся, белки SSB фиксируют нити, лидирующая цепь продолжает удлиняться, на отстающей цепи синтезируется третий фрагмент РНК-затравки - Д и с его 3’ конца начинает синтезироваться новый фрагмент Оказаки. Затем на отстающей нити ДНК РНК-затравки разрушаются и оставшиеся фрагменты ДНК соединяются в единую цепь . Таким образом, на отстающей нити идут непрерывно 4 процесса: образования новых РНК-затравок, синтез с их 3’ конца фрагментов Оказаки, разрушение РНК-затравок и воссоединение фрагментов в единую цепь.(?)
Терминация происходит тогда, когда встречаются две репликативные вилки про удвоении кольцевых молекул ДНК. Непрерывный рост лидирующей и отстающей цепей вдоль кольцевых матрицы неизбежно приводит к совмещению 3,-гидроксильного и 5,-фосфорильного концов одной цепи ,либо в точке начала репликации(однонаправленная репликация),либо при двунаправленной репликации- в середине кольца. Кольца в этих местах встречи соединяются ДНК –лигазой, при этом обычно они оказываются попарно сцепленными, т.е. образуют катенан. ДНК-гираза может расцепить зацепленные кольца, используя свою возможность вносить временный двуцепочечный разрыв(А.С.Коничев,Г.Ф. Севастьянова,2005).
При каждом клеточном делении каждая молекула ДНК должна удваиться; т.е. на каждом oriС должен происходить в точности один акт инициации репликации. Существует тонкие механизмы регуляции репликации на уровне инициации новых раундов, позитивным регулятором этого процесса является белок Dna-A.Среди моделей ,объясняющих механизм регуляторного действия белка Dna-A, наиболее распростронение получила модель титрования Dna-A:весь вновь синтезируемый белок Dna-A9титруется) Dna-A-боксами oriС хромосомы . Как только количество молекул инициатора превышает число внутриклеточных Dna-A-боксов .инициируется синтез ДНК. После запуска инициации на одном oriС освобождаются молекулы Dna-A, резко повышаются его внутриклеточная концентрация и синхронная инициация синтеза ДНК на других доступных областях начала репликации. (А.С.Коничев,Г.Ф.Севастьянова, 2005).
3. ДНК-полимеразы прокариот
Электронная библиотека © 2006-2014 | Контакты | Авторам и правообладателям
Читайте также: