
Трнк кратко и понятно
Обновлено: 05.07.2024
Выделяют три вида РНК: 1) информационная (матричная) РНК — иРНК (мРНК), 2) транспортная РНК — тРНК, 3) рибосомная РНК — рРНК.
Все виды РНК представляют собой неразветвленные полинуклеотиды, имеют специфическую пространственную конформацию и принимают участие в процессах синтеза белка. Информация о строении всех видов РНК хранится в ДНК. Процесс синтеза РНК на матрице ДНК называется транскрипцией.
Транспортные РНК содержат обычно 76 (от 75 до 95) нуклеотидов; молекулярная масса — 25 000–30 000. На долю тРНК приходится около 10% от общего содержания РНК в клетке.
Функции тРНК
1) транспорт аминокислот к месту синтеза белка, к рибосомам, 2) трансляционный посредник. В клетке встречается около 40 видов тРНК, каждый из них имеет характерную только для него последовательность нуклеотидов. Однако у всех тРНК имеется несколько внутримолекулярных комплементарных участков, из-за которых тРНК приобретают конформацию, напоминающую по форме лист клевера.
Следует подчеркнуть, что конкретная тРНК может транспортировать строго определенную аминокислоту, соответствующую ее антикодону. Специфичность соединения аминокислоты и тРНК достигается благодаря свойствам фермента аминоацил-тРНК-синтетаза.
Рибосомные РНК содержат 3000–5000 нуклеотидов; молекулярная масса — 1 000 000–1 500 000. На долю рРНК приходится 80–85% от общего содержания РНК в клетке. В комплексе с рибосомными белками рРНК образует рибосомы — органоиды, осуществляющие синтез белка. В эукариотических клетках синтез рРНК происходит в ядрышках.
Функции рРНК
1) необходимый структурный компонент рибосом и, таким образом, обеспечение функционирования рибосом; 2) обеспечение взаимодействия рибосомы и тРНК; 3) первоначальное связывание рибосомы и кодона-инициатора иРНК и определение рамки считывания, 4) формирование активного центра рибосомы.
Информационные РНК разнообразны по содержанию нуклеотидов и молекулярной массе (от 50 000 до 4 000 000). На долю иРНК приходится до 5% от общего содержания РНК в клетке.
Функции РНК
1) перенос генетической информации от ДНК к рибосомам, 2) матрица для синтеза молекулы белка, 3) определение аминокислотной последовательности первичной структуры белковой молекулы. РНК хранится в ДНК.
Синтез РНК
Транскрипция – синтез РНК на ДНК, то есть синтез комплементарной нити РНК на молекуле ДНК осуществляется ферментом РНК-полимеразой. У бактерий, например, кишечной палочки – одна РНК-полимераза, и все бактериальные ферменты очень похожи друг на друга; у высших организмов (эукариотов) – несколько ферментов, они называются РНК-полимераза I, РНК-полимераза II, РНК-полимераза III, они также имеют сходство с бактериальными ферментами, но устроены сложнее, в их состав входит больше белков.
Первый этап — инициация транскрипции – начало синтеза нити РНК, образуется первая связь между нуклеотидами. Затем идет наращивание нити, ее удлинение – элонгация, и, когда синтез завершен, происходит терминация, освобождение синтезированной РНК. РНК-полимераза при этом готова к новому циклу транскрипции.
Процесс изменения нуклеотидной последовательности РНК после синтеза носит название процессинга или редактирования РНК. После завершения транскрипции РНК часто подвергается модификациям, которые зависят от функции, выполняемой данной молекулой.
Эпистатическое взаимодействие генов
Эпистаз (от греч. epistasis — остановка, препятствие), один из типов взаимодействия генов, при котором аллели одного гена подавляют (эпистатируют) проявление аллелей других генов. Ген-подавитель действует на подавляемый ген по принципу, близкому к доминантности — рецессивности. Разница состоит в том, что эти гены не являются аллельными, т. е. занимают различные локусы в гомологичных или негомологичных хромосомах.
Гены, подавляющие действие других, неаллельных им генов, называются эпистатичными, а подавляемые — гипостатичными. Эпистатическое взаимодействие генов по своему характеру противоположно комплементарному взаимодействию. При эпистазе фермент, образующийся под контролем одного гена, полностью подавляет или нейтрализует действие фермента, контролируемого другим геном.
1. Рассмотрим эпистатическое взаимодействие генов на примере наследования окраски зерна у овса. У этой культуры установлены гены, определяющие черную и серую окраску. Оба гена являются доминантными: ген А определяет черную окраску, ген В — серую. В первом поколении в генотипе растений содержатся доминантные гены как черной окраски А, так и серой окраски В.
Так как ген А эпистатичен по отношению к гену В, он не дает ему проявиться, поэтому все гибриды в F1 будут черносеменными. В F2 происходит расщепление в соотношении 12 черных : 3 серых : 1 белый: 9 (А — В) : 3 (А — вв) : 3 (а — ВВ) : 1 (аа вв). В девяти типах зигот присутствуют оба доминантных гена А и В, но ген черной окраски А подавляет ген серой — В, вследствие чего семена имеют черную окраску.
В трех сочетаниях (АА вв, Аа вв, Аа вв) присутствует только один доминантный ген А, что также обуславливает развитие черносеменных растений. Эта группа по фенотипу совершенно сходна с первой, поэтому 12 растений из 16 будут иметь черные семена.
В трех сочетаниях (аа ВВ, аа Вв, аа Вв) присутствует только доминантный ген В, следовательно, все растения с такими генотипами будут серыми. Один генотип (аа вв) представляет собой новую комбинацию, в которой проявится белая окраска зерна, т. к. отсутствуют оба доминантных гена.
Этот тип взаимодействия неаллельных генов называют доминантным эпистазом. В отличие от него, при рецессивном эпистазе рецессивная аллель одного гена, будучи в гомозиготном состоянии, подавляет проявление доминантной или рецессивной аллели другого гена.
В этом случае вместо ожидаемого при дигибридном скрещивании расщепления (по Менделю) 9 : 3 : 3 : 1,получается соотношение 9:3:4. Эпистатическое взаимодействие генов на примере наследования окраски зерна у овса.
А – черная окраска, В – серая окраска
F2: 9/16 А_В_ — черная окраска
3/16 А_вв – черная окраска
3/16 ввВ_ — серая окраска
1/16 аавв – белая окраска
Типы РНК и функции различных типов РНК
Эпистатическое взаимодействие генов
Эпистаз (от греч. epistasis — остановка, препятствие), один из типов взаимодействия генов, при котором аллели одного гена подавляют (эпистатируют) проявление аллелей других генов. Ген-подавитель действует на подавляемый ген по принципу, близкому к доминантности — рецессивности.
Гены, подавляющие действие других, неаллельных им генов, называются эпистатичными, а подавляемые — гипостатичными. Эпистатическое взаимодействие генов по своему характеру противоположно комплементарному взаимодействию.
При эпистазе фермент, образующийся под контролем одного гена, полностью подавляет или нейтрализует действие фермента, контролируемого другим геном.
1. Рассмотрим эпистатическое взаимодействие генов на примере наследования окраски зерна у овса. У этой культуры установлены гены, определяющие черную и серую окраску. Оба гена являются доминантными: ген А определяет черную окраску, ген В — серую. В первом поколении в генотипе растений содержатся доминантные гены как черной окраски А, так и серой окраски В.
Так как ген А эпистатичен по отношению к гену В, он не дает ему проявиться, поэтому все гибриды в F1 будут черносеменными. В F2 происходит расщепление в соотношении 12 черных : 3 серых : 1 белый: 9 (А — В) : 3 (А — вв) : 3 (а — ВВ) : 1 (аа вв). В девяти типах зигот присутствуют оба доминантных гена А и В, но ген черной окраски А подавляет ген серой — В, вследствие чего семена имеют черную окраску.
В трех сочетаниях (АА вв, Аа вв, Аа вв) присутствует только один доминантный ген А, что также обуславливает развитие черносеменных растений. Эта группа по фенотипу совершенно сходна с первой, поэтому 12 растений из 16 будут иметь черные семена. В трех сочетаниях (аа ВВ, аа Вв, аа Вв) присутствует только доминантный ген В, следовательно, все растения с такими генотипами будут серыми.
Один генотип (аа вв) представляет собой новую комбинацию, в которой проявится белая окраска зерна, т. к. отсутствуют оба доминантных гена. Этот тип взаимодействия неаллельных генов называют доминантным эпистазом. В отличие от него, при рецессивном эпистазе рецессивная аллель одного гена, будучи в гомозиготном состоянии, подавляет проявление доминантной или рецессивной аллели другого гена.
В этом случае вместо ожидаемого при дигибридном скрещивании расщепления (по Менделю) 9 : 3 : 3 : 1,получается соотношение 9:3:4. Эпистатическое взаимодействие генов на примере наследования окраски зерна у овса.
А – черная окраска, В – серая окраска
F2: 9/16 А_В_ — черная окраска
3/16 А_вв – черная окраска
3/16 ввВ_ — серая окраска
1/16 аавв – белая окраска
Оба гена являются доминантными: ген А определяет черную окраску, ген В — серую. В первом поколении в генотипе растений содержатся доминантные гены как черной окраски А, так и серой окраски В. Так как ген А эпистатичен по отношению к гену В, он не дает ему проявиться, поэтому все гибриды в F1 будут черносеменными. В F2 происходит расщепление в соотношении 12 черных : 3 серых : 1 белый: 9 (А — В) : 3 (А — вв) : 3 (а — ВВ) : 1 (аа вв).
В девяти типах зигот присутствуют оба доминантных гена А и В, но ген черной окраски А подавляет ген серой — В, вследствие чего семена имеют черную окраску. В трех сочетаниях (АА вв, Аа вв, Аа вв) присутствует только один доминантный ген А, что также обуславливает развитие черносеменных растений.
Эта группа по фенотипу совершенно сходна с первой, поэтому 12 растений из 16 будут иметь черные семена. В трех сочетаниях (аа ВВ, аа Вв, аа Вв) присутствует только доминантный ген В, следовательно, все растения с такими генотипами будут серыми.
Один генотип (аа вв) представляет собой новую комбинацию, в которой проявится белая окраска зерна, т. к. отсутствуют оба доминантных гена. Этот тип взаимодействия неаллельных генов называют доминантным эпистазом.
В отличие от него, при рецессивном эпистазе рецессивная аллель одного гена, будучи в гомозиготном состоянии, подавляет проявление доминантной или рецессивной аллели другого гена. В этом случае вместо ожидаемого при дигибридном скрещивании расщепления (по Менделю) 9 : 3 : 3 : 1,получается соотношение 9:3:4. Эпистатическое взаимодействие генов на примере наследования окраски зерна у овса.
А – черная окраска, В – серая окраска
F2: 9/16 А_В_ — черная окраска
3/16 А_вв – черная окраска
3/16 ввВ_ — серая окраска
1/16 аавв – белая окраска
РНК (рибонуклеиновая кислота)
РНК (рибонуклеиновая кислота), так же как и ДНК, относится к нуклеиновым кислотам. Молекулы-полимеры РНК намного меньше, чем у ДНК. Однако в зависимости от типа РНК количество входящих в них нуклеотидов-мономеров различается.
В состав нуклеотида РНК в качестве сахара входит рибоза, в качестве азотистого основания — аденит, гуанин, урацил, цитозин.
Урацил по строению и химическим свойствам близок к тимину, который обычен для ДНК. В зрелых молекулах РНК многие азотистые основания модифицированы, поэтому в реальности разновидностей азотистых оснований в составе РНК намного больше.
Рибоза в отличие от дезоксирибозы имеет дополнительную -ОН-группу (гидроксильную). Это обстоятельство позволяет РНК легче вступать в химические реакции.
Главной функцией РНК в клетках живых организмов можно назвать реализацию генетической информации.
Именно благодаря разным типам рибонуклеиновой кислоты генетический код считывается (транскрибируется) с ДНК, после чего на его основе синтезируются полипептиды (происходит трансляция). Итак, если ДНК в основном отвечает за хранение и передачу из поколения в поколение генетической информации (основной процесс – репликация), то РНК реализует эту информацию (процессы транскрипции и трансляции).
При этом транскрипция происходит на ДНК, так что этот процесс относится к обоим типам нуклеиновых кислот и тогда с этой точки зрения можно сказать, что и ДНК отвечает за реализацию генетической информации.
При более подробном рассмотрении функции РНК намного разнообразнее. Ряд молекул РНК выполняют структурную, каталитическую и другие функции.
Существует так называемая гипотеза РНК-мира, согласно которой вначале в живой природе в качестве носителя генетической информации выступали только молекулы РНК, при этом другие молекулы РНК катализировали различные реакции.
Данная гипотеза подтверждена рядом опытов, показывающих возможную эволюцию РНК. На это указывает и то, что ряд вирусов в качестве нуклеиновой кислоты, хранящей генетическую информацию, имеют молекулу РНК.
Согласно гипотезе РНК-мира ДНК появилась позже в процессе естественного отбора как более устойчивая молекула, что важно для хранения генетической информации.
Выделяют три основных типа РНК (кроме них есть и другие): матричная (она же информационная), рибосомальная и транспортная.
Обозначаются они соответственно иРНК (или мРНК), рРНК, тРНК.
Информационная РНК (иРНК)
Почти все РНК синтезируются на ДНК в процессе транскрипции. Однако часто транскрипция упоминается как синтез именно информационной РНК (иРНК). Связано это с тем, что последовательность нуклеотидов иРНК в последствии определит последовательность аминокислот синтезируемого в процессе трансляции белка.
Перед транскрипцией нити ДНК расплетаются, и на одной из них с помощью комплекса белков-ферментов синтезируется РНК по принципу комплементарности, так же как это происходит при репликации ДНК.
Только напротив аденина ДНК к молекуле РНК присоединяется нуклеотид, содержащий урацил, а не тимин.
На самом деле на ДНК синтезируется не готовая информационная РНК, а ее предшественник — пре-иРНК.
После удаления интронов экзоны сшиваются между собой. Процесс удаления интронов и сшивания экзонов называется сплайсингом. Усложняющей жизнь особенностью является то, что можно вырезать интроны по-разному, в результате получатся разные готовые иРНК, которые будут служить матрицами для разных белков. Таким образом, вроде бы один ген ДНК может играть роль нескольких генов.
Следует отметить, что у прокариотических организмов сплайсинга не происходит.
Обычно их иРНК сразу после синтеза на ДНК готова к трансляции. Бывает, что пока конец молекулы иРНК еще транскрибируется, на ее начале уже сидят рибосомы, синтезирующие белок.
Рибосома, передвигаясь от начала иРНК к ее концу, считывает по три нуклеотида (хотя вмещает шесть, т.

Когда молекула информационной РНК становится не нужна, клетка ее разрушает.
Транспортная РНК (тРНК)
Транспортная РНК — это достаточно маленькая (по меркам полимеров) молекула (количество нуклеотидов бывает разным, в среднем около 80-ти), во вторичной структуре имеет форму клеверного листа, в третичной сворачивается в нечто подобное букве Г.

Функция тРНК — присоединение к себе соответствующей своему антикодону аминокислоты.
Живая природа на Земле использует всего около 20-ти аминокислот для синтеза различных белковых молекул (на самом деле аминокислот куда больше).
Но поскольку, согласно генетическому коду, кодонов больше 60-ти, то каждой аминокислоте может соответствовать несколько кодонов (на самом деле какой-то больше, какой-то меньше). Таким образом, разновидностей тРНК больше 20, при этом разные транспортные РНК переносят одинаковые аминокислоты. (Но и тут не так все просто.)
- Транспортная РНК, тРНК — рибонуклеиновая кислота, функцией которой является транспортировка аминокислот к месту синтеза белка. Имеет типичную длину от 73 до 93 нуклеотидов и размеры около 5 нм. тРНК также принимают непосредственное участие в наращивании полипептидной цепи, присоединяясь — будучи в комплексе с аминокислотой — к кодону мРНК и обеспечивая необходимую для образования новой пептидной связи конформацию комплекса.
Для каждой аминокислоты существует своя тРНК.
На участке C находится антикодон, соответствующий аминокислоте.
тРНК синтезируются обычной РНК-полимеразой в случае прокариот и РНК-полимеразой III в случае эукариот. Транскрипты генов тРНК подвергаются многостадийному процессингу, который в конце концов приводит к формированию типичной для тРНК пространственной структуры. Процессинг тРНК включает 5 ключевых этапов:
* удаление 5'-лидерной нуклеотидной последовательности;
* удаление 3'-концевой последовательности;
* добавление последовательности CCA на 3'-конец;
* вырезание интронов (у эукариот и архей);
Связанные понятия
Рибосо́ма — важнейшая немембранная органелла живой клетки, служащая для биосинтеза белка из аминокислот по заданной матрице на основе генетической информации, предоставляемой матричной РНК (мРНК). Этот процесс называется трансляцией. Рибосомы имеют сферическую или слегка эллипсоидную форму, диаметром от 15—20 нанометров (прокариоты) до 25—30 нанометров (эукариоты), состоят из большой и малой субъединиц.
Трансля́ция (от лат. translatio — перенос, перемещение) — процесс синтеза белка из аминокислот на матрице информационной (матричной) РНК (иРНК, мРНК), осуществляемый рибосомой.
Комплемента́рность (в химии, молекулярной биологии и генетике) — взаимное соответствие молекул биополимеров или их фрагментов, обеспечивающее образование связей между пространственно взаимодополняющими (комплементарными) фрагментами молекул или их структурных фрагментов вследствие супрамолекулярных взаимодействий (образование водородных связей, гидрофобных взаимодействий, электростатических взаимодействий заряженных функциональных групп и т. п.).
Процессинг РНК (посттранскрипционные модификации РНК) — совокупность процессов в клетках эукариот, которые приводят к превращению первичного транскрипта в зрелую РНК.
Гисто́ны — обширный класс ядерных белков, выполняющих две основные функции: они участвуют в упаковке нитей ДНК в ядре и в эпигенетической регуляции таких ядерных процессов, как транскрипция, репликация и репарация. Существует пять различных типов гистонов H1/Н5, H2A, H2B, H3, H4. Гистоны H2A, H2B, H3, H4, называемые кóровыми гистонами (от англ. core — сердцевина), формируют нуклеосому, представляющую собой белковую глобулу, вокруг которой накручена нить ДНК. Гистон H1/H5, называемый линкерным гистоном.
Промотор в генетике — последовательность нуклеотидов ДНК, узнаваемая РНК-полимеразой как стартовая площадка для начала транскрипции. Промотор играет одну из ключевых ролей в процессе инициации транскрипции.
Нуклеоти́ды (нуклеозидфосфаты) — группа органических соединений, представляют собой фосфорные эфиры нуклеозидов. Свободные нуклеотиды, в частности АТФ, цАМФ, АДФ, играют важную роль в энергетических и информационных внутриклеточных процессах, а также являются составляющими частями нуклеиновых кислот и многих коферментов.
Транскри́пция (от лат. transcriptio — переписывание) — процесс синтеза РНК с использованием ДНК в качестве матрицы, происходящий во всех живых клетках. Другими словами, это перенос генетической информации с ДНК на РНК.
Доме́н белка́ — элемент третичной структуры белка, представляющий собой достаточно стабильную и независимую подструктуру белка, фолдинг которой проходит независимо от остальных частей. В состав домена обычно входит несколько элементов вторичной структуры. Сходные по структуре домены встречаются не только в родственных белках (например, в гемоглобинах разных животных), но и в совершенно разных белках.
Консервати́вные после́довательности (англ. conserved sequences) — схожие или идентичные последовательности, встречающиеся в биологических полимерах: нуклеиновых кислотах, первичной и пространственной структурах белков, полисахаридах как в пределах особей разных видов (ортологичные последовательности), так и в пределах одной особи (паралогичные последовательности). Ортологичные последовательности являются подтверждением того, что определённые последовательности могут поддерживаться эволюцией, несмотря.
Оперон — функциональная единица генома у прокариот, в состав которой входят цистроны (гены, единицы транскрипции), кодирующие совместно или последовательно работающие белки и объединенные под одним (или несколькими) промоторами. Такая функциональная организация позволяет эффективнее регулировать транскрипцию этих генов.
РНК-полимераза II — фермент эукариот, который катализирует транскрипцию ДНК, синтезирует предшественников мРНК и большинство мяРНК и микроРНК. Эта полимераза представляет собой комплекс массой 550 кДа, состоящий из 12 субъединиц. РНК-полимераза II является наиболее изученным типом РНК-полимеразы. Ей необходим широкий спектр транскрипционных факторов для того, чтобы связываться с генами выше промоторов и начинать транскрипцию.
Белковая субъединица в структурной биологии — полипептид, который вместе с другими компонентами собирается в мультимерный или олигомерный белковый комплекс. Многие природные ферменты и другие белки состоят из нескольких белковых субъединиц.
Открытая рамка считывания (англ. Open Reading Frame, ORF) — последовательность нуклеотидов в составе ДНК или РНК, потенциально способная кодировать белок. Основным признаком наличия ORF служит отсутствие стоп-кодонов (в случае РНК — обычно UAA, UGA и UAG) на достаточно длинном участке последовательности после стартового кодона (в подавляющем большинстве случаев — AUG). Поскольку в некоторых случаях стартовый и терминирующие кодоны отличаются от канонических, а также ввиду возможности супрессии (подавления.
Полиаденили́рование — это процесс присоединения большого количества остатков аденозинмонофосфата (поли(А)-хвоста) к 3'-концу первичной мРНК (пре-мРНК). Иными словами, поли(А)-хвост — это фрагмент молекулы мРНК, азотистые основания которого представлены только аденином. У эукариот полиаденилирование является частью процессинга мРНК — процесса созревания первичного транскрипта в зрелую мРНК, готовую для трансляции. Процессинг, в свою очередь, является одним из этапов экспрессии генов.
Экзоны (от англ. ex(pressi)on — выражение) — участки ДНК, копии которых составляют зрелую РНК. По мнению некоторых исследователей экзоны соответствуют доменам (структурно автономным областям) в белке и являются первичными генетическими единицами, перекомбинация которых приводит к возникновению в ходе эволюции новых генов и соответственно новых белков. Экзоны чередуются в структуре гена с другими фрагментами — интронами. При альтернативном сплайсинге некоторые экзоны удаляются из зрелой РНК.
Эндонуклеазы — белки из группы нуклеаз, расщепляющие фосфодиэфирные связи в середине полинуклеотидной цепи. Эндонуклеазы рестрикции, или рестриктазы, расщепляют ДНК в определенных местах (так называемых сайтах рестрикции), они подразделяются на три типа (I, II и III) на основании механизма действия. Эти белки часто используют в генной инженерии для создания рекомбинантных ДНК, которые вводят затем в бактериальные, растительные или животные клетки.
Вторичная структура — конформационное расположение главной цепи (англ. backbone) макромолекулы (например, полипептидная цепь белка или цепи нуклеиновых кислот), независимо от конформации боковых цепей или отношения к другим сегментам. В описании вторичной структуры важным является определение водородных связей, которые стабилизируют отдельные фрагменты макромолекул.
Старт-кодон или инициаторный кодон — первый кодон матричной РНК, c которого начинается трансляция белка в рибосоме. У эукариот и архей старт-кодон всегда кодирует метионин, а у прокариот— модифицированный метионин (N-формилметионин). В большинстве случаев роль инициаторного кодона играет триплет AUG. Старт-кодону предшествует 5′-нетранслируемая область (5'-UTR). В 5'-UTR бактерий локализована последовательность Шайна — Дальгарно (AGGAGG), которая служит для связывания рибосомы и отделёна спейсером.
Цис-регуляторные элементы (или цис-элементы) — участки ДНК или РНК, регулирующие экспрессию генов, находящихся на той же молекуле (обычно хромосоме).
Кодо́н (кодирующий тринуклеотид) — единица генетического кода, тройка нуклеотидных остатков (триплет) в ДНК или РНК, обычно кодирующих включение одной аминокислоты. Последовательность кодонов в гене определяет последовательность аминокислот в полипептидной цепи белка, кодируемого этим геном.
Спаренные основания — пара двух азотистых оснований нуклеотидов на комплементарных цепочках нуклеиновых кислот (противоположных ДНК или одинаковых РНК), соединённая с помощью водородных связей.
Сплайсосо́ма — структура, состоящая из молекул РНК и белков и осуществляющая удаление некодирующих последовательностей (интронов) из предшественников мРНК. Этот процесс называется сплайсингом (от англ. splicing — сращивание).
Факторы транскрипции (транскрипционные факторы) — белки́, контролирующие процесс синтеза мРНК на матрице ДНК (транскрипцию) путём связывания со специфичными участками ДНК. Транскрипционные факторы выполняют свою функцию либо самостоятельно, либо в комплексе с другими белками. Они обеспечивают снижение (репрессоры) или повышение (активаторы) константы связывания РНК-полимеразы с регуляторными последовательностями регулируемого гена.
Транскрипт — молекула РНК, образующаяся в результате транскрипции (экспрессии соответствующего гена или участка ДНК).
Повторя́ющиеся после́довательности ДНК (англ. Repetitive DNA) — участки ДНК, включённые в геном, последовательность которых состоит из повторяющихся фрагментов. Выделяют 2 типа таких повторяющихся последовательностей.
Нуклеосома — это структурная часть хромосомы, образованная совместной упаковкой нити ДНК с гистоновыми белками H2А, H2B, H3 и H4. Последовательность нуклеосом, соединенная гистоновым белком H1, формирует нуклеофиламент, или иначе нуклеосомную нить.
Стоп-кодон или кодон терминации — единица генетического кода, тройка нуклеотидных остатков (триплет) в ДНК — кодирующая прекращение (терминацию) синтеза полипептидной цепи (трансляцию).
Кэп, 5'-кэп (произносится как пять-штрих-кэп), или кэп-структура (от англ. cap — шапка) — структура на 5'-конце матричных РНК (мРНК) и некоторых других РНК эукариот. Кэп состоит из одного или нескольких модифицированных нуклеотидов и характерен только для транскриптов, синтезируемых РНК-полимеразой II. Наличие кэпа — один из признаков, отличающих эукариотические мРНК от прокариотических, которые несут трифосфат на 5'-конце. Это и другие отличия обуславливают существенно более высокую стабильность.
Праймер (англ. primer) — это короткий фрагмент нуклеиновой кислоты (олигонуклеотид), комплементарный ДНК- или РНК-мишени, служит затравкой для синтеза комплементарной цепи с помощью ДНК-полимеразы (при репликации ДНК). Затравка необходима ДНК-полимеразам для инициации синтеза новой цепи, с 3'-конца (гидроксильной группы) праймера. ДНК-полимераза последовательно добавляет к 3'-концу праймера нуклеотиды, комплементарные матричной цепи.
Рибонуклеазы (РНКазы, англ. Ribonuclease, RNase) — ферменты-нуклеазы, катализирующие деградацию РНК. Рибонуклеазы классифицируют на эндорибонуклеазы и экзорибонуклеазы. К рибонуклеазам относят некоторые подклассы КФ 2.7 и КФ 3.1.
Ми́кроРНК (англ. microRNA, miRNA) — малые некодирующие молекулы РНК длиной 18—25 нуклеотидов (в среднем 22), обнаруженные у растений, животных и некоторых вирусов, принимающие участие в транскрипционной и посттранскипционной регуляции экспрессии генов путём РНК-интерференции. Помимо внутриклеточной обнаружена внеклеточная (циркулирующая) микроРНК.МикроРНК кодируются ядерной ДНК растений и животных и вирусной ДНК у некоторых ДНК-содержащих вирусов.
Шпи́лька (англ. stem-loop, hairpin) — в молекулярной биологии элемент вторичной структуры РНК, а также одноцепочечной ДНК. Шпилька образуется в том случае, когда две последовательности одной и той же цепи комплементарны друг другу и соединяются друг с другом, перегибаясь одна к другой и образуя на конце неспаренный участок — петлю. Такие комплементарные последовательности нередко представляют собой палиндромные последовательности.
ДНК-метилтрансфера́зы (ДНК-метилазы, англ. DNA methyltransferase, DNA MTase, DNMT) — группа ферментов, катализирующих метилирование нуклеотидных остатков в составе ДНК. Активность метилтрансфераз, заключающаяся в переносе метильных (CH3—) групп на азотистое основание цитозин в составе ДНК, ведет к изменению свойств ДНК, при этом изменяется активность, функции соответствующих генов, а также пространственная структура нуклеиновой кислоты (конформация).
Изоформа белка — любая из нескольких разных форм одного и того же белка. Различные формы белка могут быть образованы связанными генами, или могут возникнуть из того же гена путём альтернативного сплайсинга. Большое количество изоформ вызваны однонуклеотидными полиморфизмами — небольшими генетическими различиями между аллелями одного и того же гена. Это происходит в определенных отдельных местах расположения нуклеотидов на гене.
Экзонуклеазы — белки из группы нуклеаз, отщепляющие концевые мононуклеотиды от полинуклеотидной цепи путём гидролиза фосфодиэфирных связей между нуклеотидами.
Последовательность Шайна — Дальгарно (англ. Shine-Dalgarno sequence, Shine-Dalgarno box) — сайт связывания рибосом на молекуле мРНК прокариот, обычно на расстоянии около 10 нуклеотидов до стартового кодона AUG. Описана австралийскими учёными Джоном Шайном и Линн Дальгарно.Консенсусом является последовательность из шести нуклеотидов AGGAGG; в случае E. coli последовательность Шайна — Дальгарно — AGGAGGU. Комплементарная последовательность CCUCCU, называемая последовательностью анти-Шайна — Дальгарно.
Комплементарная ДНК (кДНК, англ. сDNA) — это ДНК, синтезированная на матрице зрелой мРНК в реакции, катализируемой обратной транскриптазой.
Аминоацил-тРНК-синтетаза (АРСаза) — фермент-синтетаза, катализирующий образование аминоацил-тРНК в реакции этерификации определённой аминокислоты с соответствующей ей молекулой тРНК. Для каждой протеиногенной аминокислоты существует своя аминоацил-тРНК-синтетаза.
Эукарио́ты (устар. эвкарио́ты; лат. Eukaryota от др.-греч. εὖ- ‘хорошо’ или ’полностью’ + κάρυον ‘ядро’), или я́дерные, — домен (надцарство) живых организмов, клетки которых содержат ядро. Все организмы, кроме прокариот (бактерий и архей), являются ядерными. Вирусы и вироиды также не являются ни прокариотами, ни эукариотами; более того, сам вопрос, считать ли их живыми организмами, является дискуссионным.

Вторым этапом реализации генетической информации является синтез белковой молекулы на основе матричной РНК (трансляция). Однако в отличие от транскрипции нуклеотидная последовательность не может быть переведена в аминокислотную напрямую, так как эти соединения имеют разную химическую природу. Поэтому для осуществления трансляции требуется посредник в виде транспортной РНК (тРНК), функция которого состоит в переводе генетического кода на "язык" аминокислот.
Общая характеристика транспортной РНК
Транспортные РНК или тРНК - это небольшие молекулы, которые доставляют аминокислоты к месту синтеза белка (в рибосомы). Количество этого вида рибонуклеиновой кислоты в клетке составляет примерно 10 % от общего пула РНК.
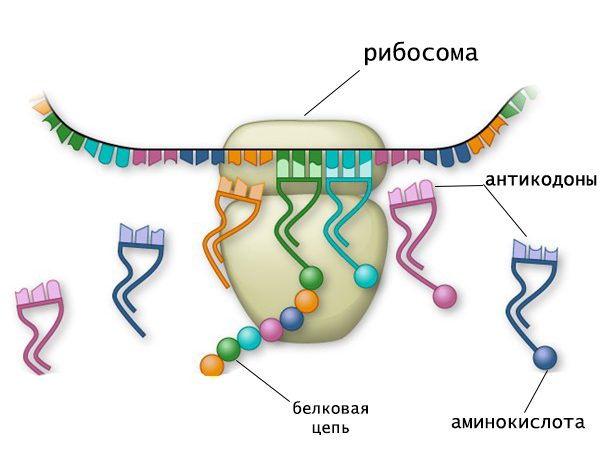
Как и другие разновидности рибонуклеиновых кислот, тРНК состоит из цепочки рибонуклеозидтрифосфатов. Длина нуклеотидной последовательности насчитывает 70-90 звеньев, и около 10 % состава молекулы приходится на минорные компоненты.
Из-за того, что каждой аминокислоте соответствует свой переносчик в виде тРНК, клетка синтезирует большое количество разновидностей этой молекулы. В зависимости от вида живого организма этот показатель варьирует от 80 до 100.
Функции тРНК
Транспортная РНК является поставщиком субстрата для белкового синтеза, который происходит в рибосомах. За счет уникальной способности связываться и с аминокислотами, и с матричной последовательностью тРНК выполняет функцию смыслового адаптера при переводе генетической информации из формы РНК в форму белка. Взаимодействие такого посредника с кодирующей матрицей, как в транскрипции, основано на принципе комплементарности азотистых оснований.
Главная функция тРНК заключается в акцептировании аминокислотных звеньев и их транспортировке в аппарат белкового синтеза. За этим техническим процессом стоит огромный биологический смысл - реализация генетического кода. Осуществление этого процесса основано на следующих особенностях:
- все аминокислоты кодируются триплетами нуклеотидов;
- для каждого триплета (или кодона) существует антикодон, входящий в состав тРНК;
- каждая тРНК может связаться только с определенной аминокислотой.
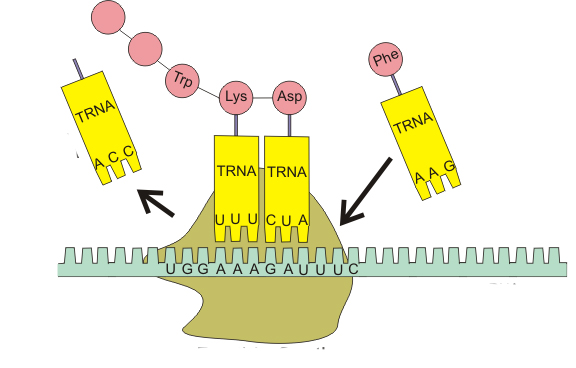
Таким образом, аминокислотная последовательность белка определяется тем, какие тРНК и в каком порядке будут комплементарно взаимодействовать с матричной РНК в процессе трансляции. Это возможно благодаря наличию в транспортной РНК функциональных центров, один из которых отвечает за избирательное присоединение аминокислоты, а другой - за связывание с кодоном. Поэтому функции и структура тРНК тесно взаимосвязаны.
Строение транспортной РНК
Уникальность тРНК заключается в том, что ее молекулярная структура не линейна. Она включает в себя спиральные двуцепочечные участки, которые называют стеблями, и 3 одноцепочечные петли. По форме такая конформация напоминает клеверный лист.
В структуре тРНК различают следующие стебли:
- акцепторный;
- антикодоновый;
- дигидроуридиловый;
- псевдоуридиловый;
- добавочный.
Двойные спирали стеблей содержат от 5 до 7 Уотсон-Криксоновских пар. На конце акцепторного стебля расположена небольшая цепочка неспаренных нуклеотидов, 3-гидроксил которой является местом прикрепления соответствующей молекулы аминокислоты.
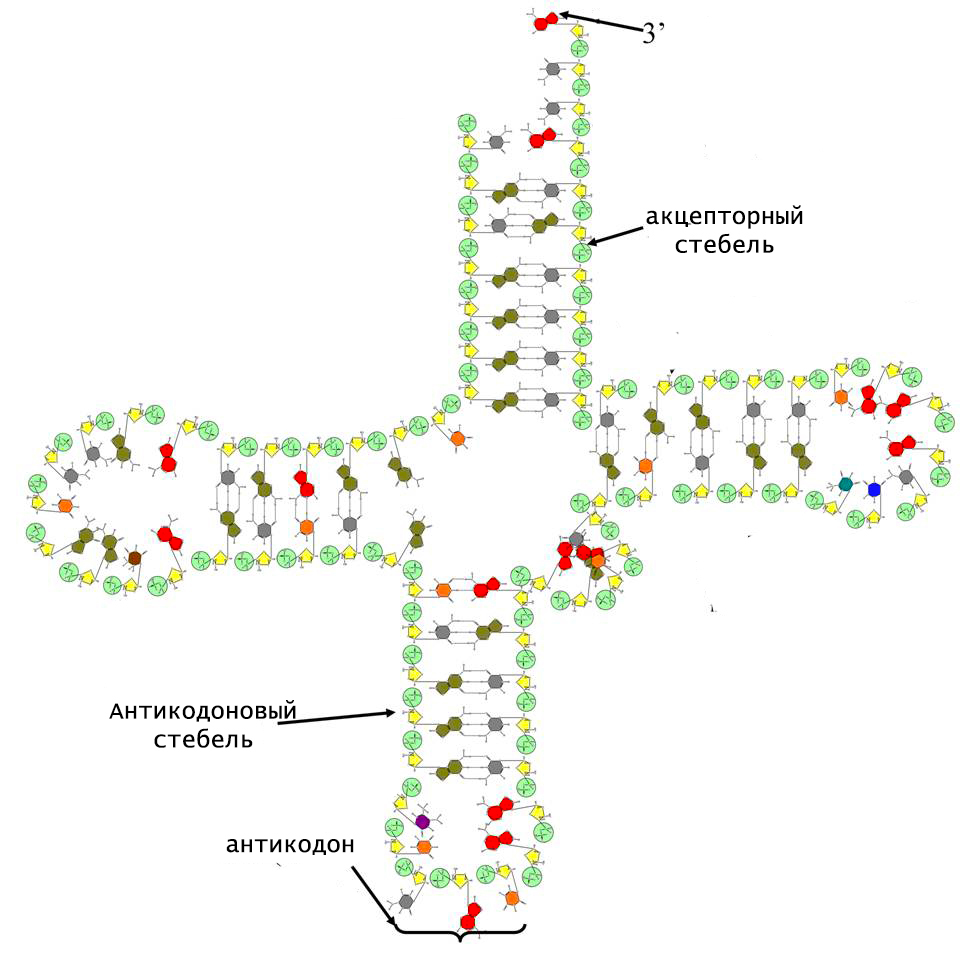
Структурной областью для соединения с мРНК служит одна из петель тРНК. Она содержит антикодон, комплементарный смысловому триплету в матричной РНК. Именно антикодон и акцептирующий конец обеспечивают адапторную функцию тРНК.
Третичная структура молекулы
"Клеверный лист" является вторичной структурой тРНК, однако за счет фолдинга молекула приобретает L-образную конформацию, которая скрепляется дополнительными водородными связями.
L-форма представляет собой третичную структуру тРНК и состоит из двух практически перпендикулярных А-РНК спиралей, имеющих длину 7 нм и толщину 2 нм. Такая форма молекулы имеет всего 2 конца, на одном из которых расположен антикодон, а на другом - акцепторный центр.

Особенности связывания тРНК с аминокислотой
Активацию аминокислот (их присоединение к транспортной РНК) осуществляет аминоацил-тРНК-синтетаза. Этот фермент одновременно выполняет 2 важных функции:
- катализирует образования ковалентной связи между 3`-гидроксильной группой акцепторного стебля и аминокислотой;
- обеспечивает принцип избирательного соответствия.
Для каждой из 20 аминокислот есть своя аминоацил-тРНК-синтетаза. Она может взаимодействовать только с соответствующим видом транспортной молекулы. Это означает, что антикодон последней должен быть комплементарен триплету, кодирующему именно эту аминокислоту. Например, синтетаза лейцина будет соединяться только с предназначеным для лейцина тРНК.
В молекуле аминоацил-тРНК-синтетазы есть три нуклеотид-связывающих кармана, конформация и заряд которых комплементарны нуклеотидам соответствующего антикодона в тРНК. Таким образом, фермент определяет нужную транспортную молекулу. Гораздо реже фрагментом опознавания служит нуклеотидная последовательность акцепторного стебля.
И транскрипция, и трансляция относятся к матричным биосинтезам. Матричным биосинтезом называется синтез биополимеров (нуклеиновых кислот, белков) на матрице - нуклеиновой кислоте ДНК или РНК. Процессы матричного биосинтеза относятся к пластическому обмену: клетка расходует энергию АТФ.
Матричный синтез можно представить как создание копии исходной информации на несколько другом или новом "генетическом языке". Скоро вы все поймете - мы научимся достраивать по одной цепи ДНК другую, переводить РНК в ДНК и наоборот, синтезировать белок с иРНК на рибосоме. В данной статье вас ждут подробные примеры решения задач, генетический словарик пригодится - перерисуйте его себе :)

Возьмем 3 абстрактных нуклеотида ДНК (триплет) - АТЦ. На иРНК этим нуклеотидам будут соответствовать - УАГ (кодон иРНК). тРНК, комплементарная иРНК, будет иметь запись - АУЦ (антикодон тРНК). Три нуклеотида в зависимости от своего расположения будут называться по-разному: триплет, кодон и антикодон. Обратите на это особое внимание.
Репликация ДНК - удвоение, дупликация (лат. replicatio — возобновление, лат. duplicatio - удвоение)
Процесс синтеза дочерней молекулы ДНК по матрице родительской ДНК. Нуклеотиды достраивает фермент ДНК-полимераза по принципу комплементарности. Переводя действия данного фермента на наш язык, он следует следующему правилу: А (аденин) переводит в Т (тимин), Г (гуанин) - в Ц (цитозин).
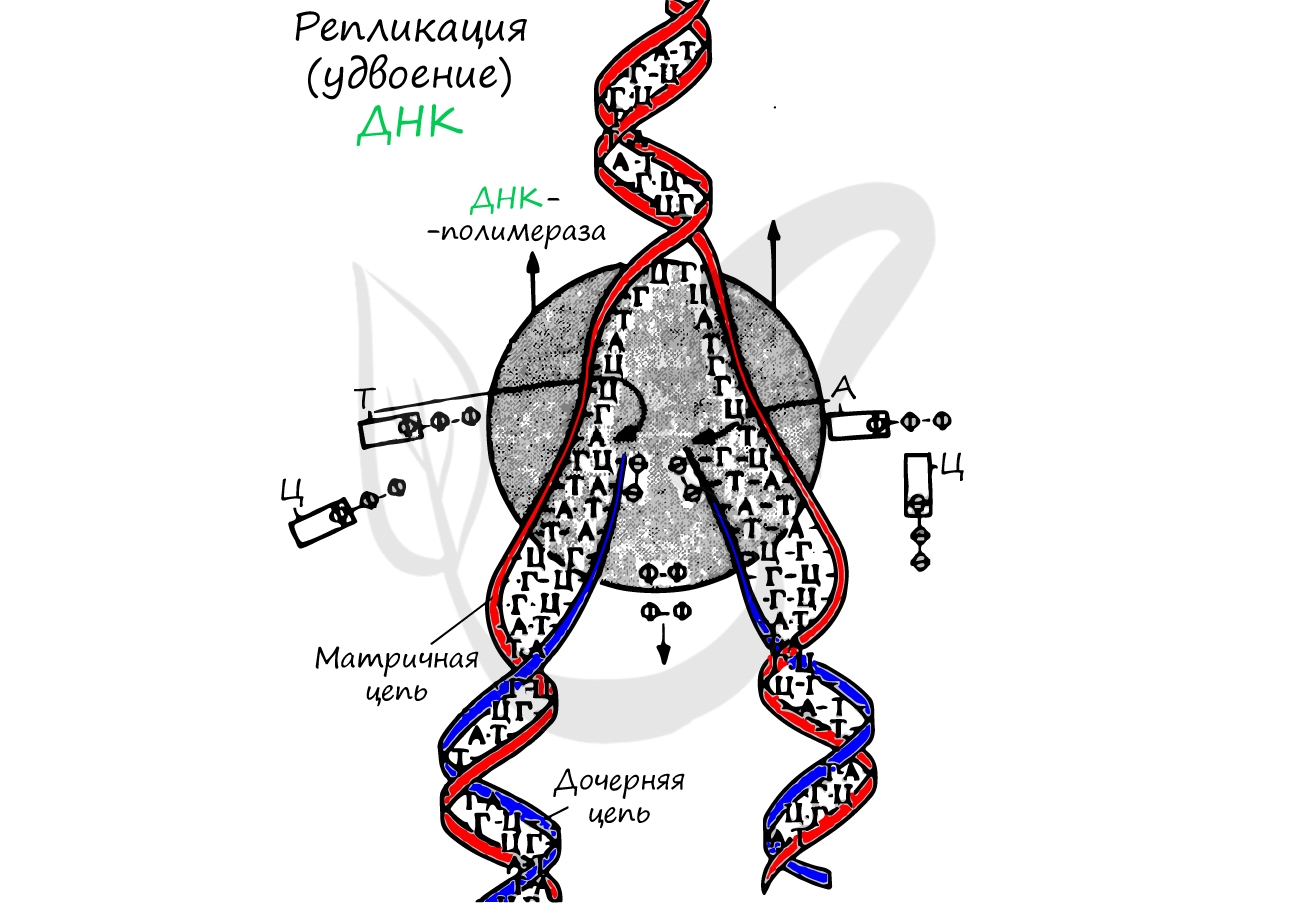
Удвоение ДНК происходит в синтетическом периоде интерфазы. При этом общее число хромосом не меняется, однако каждая из них содержит к началу деления две молекулы ДНК: это необходимо для равномерного распределения генетического материала между дочерними клетками.
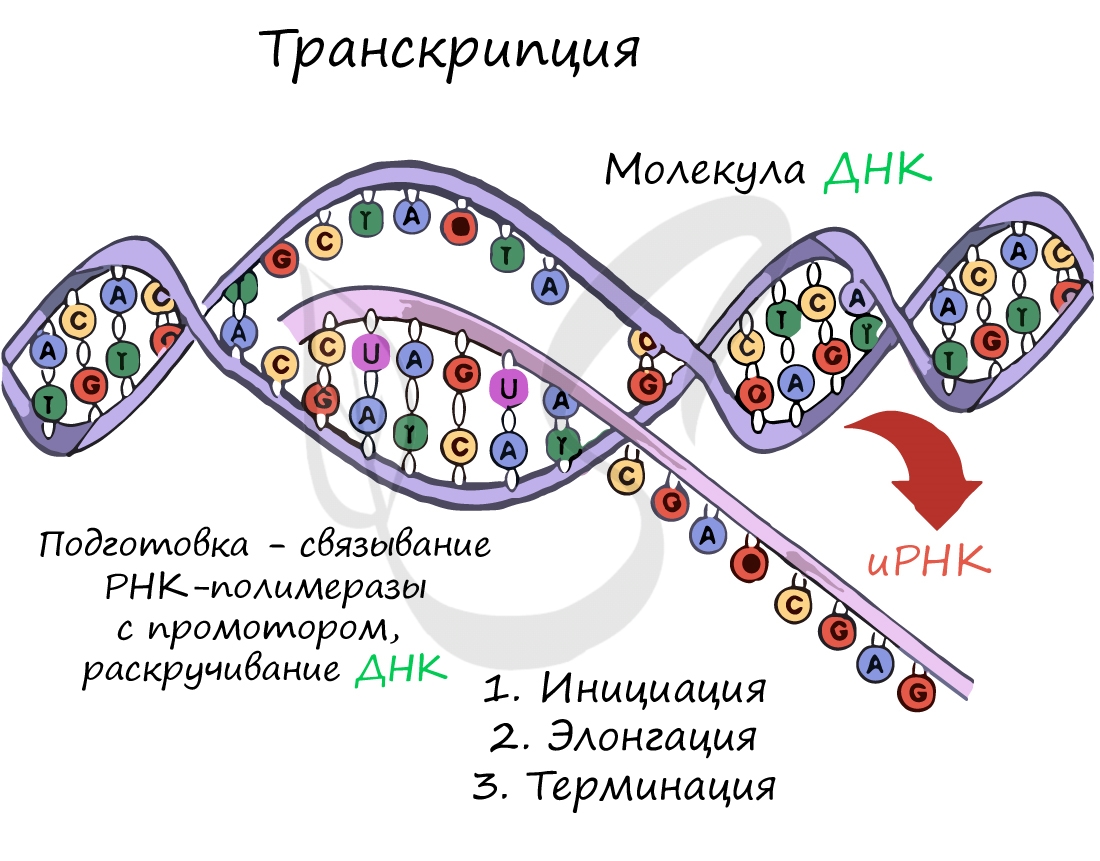
Транскрипция (лат. transcriptio — переписывание)
Транскрипция представляет собой синтез информационной РНК (иРНК) по матрице ДНК. Несомненно, транскрипция происходит в соответствии с принципом комплементарности азотистых оснований: А - У, Т - А, Г - Ц, Ц - Г (загляните в "генетический словарик" выше).
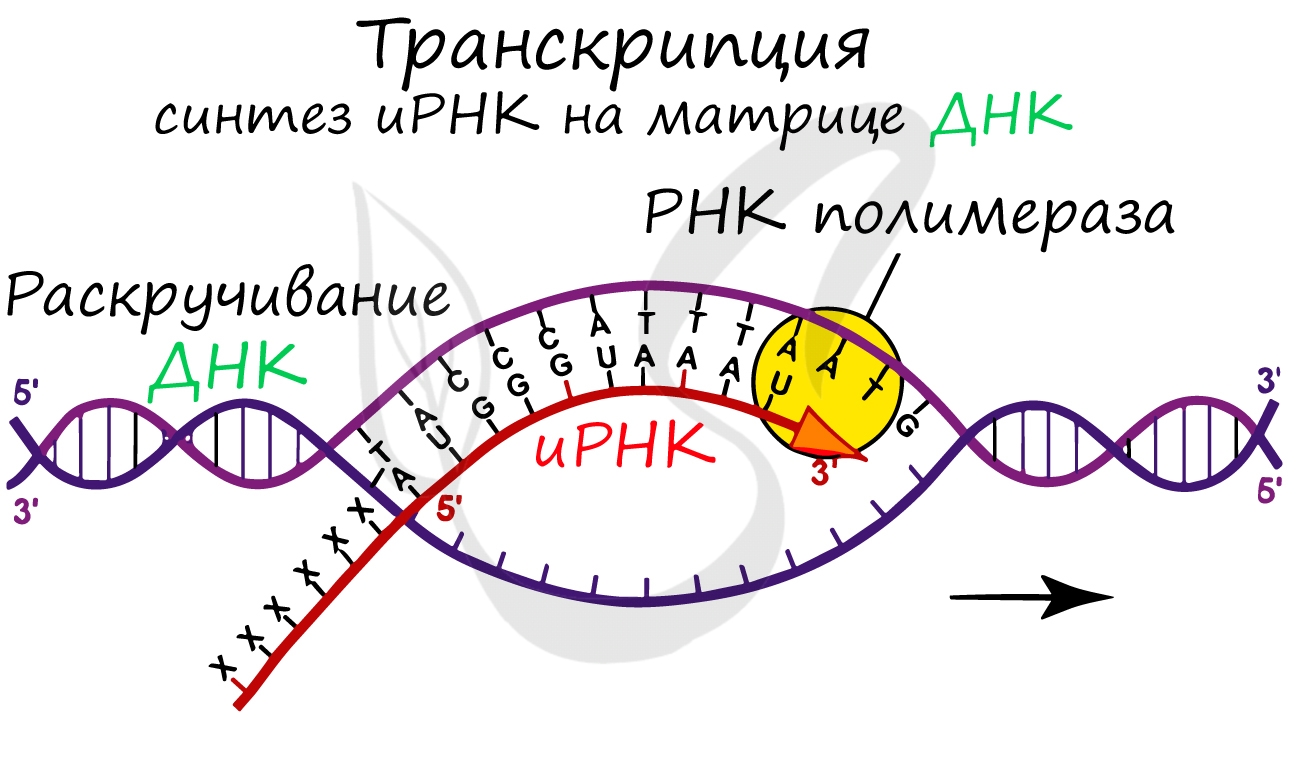
До начала непосредственно транскрипции происходит подготовительный этап: фермент РНК-полимераза узнает особый участок молекулы ДНК - промотор и связывается с ним. После связывания с промотором происходит раскручивание молекулы ДНК, состоящей из двух цепей: транскрибируемой и смысловой. В процессе транскрипции принимает участие только транскрибируемая цепь ДНК.
-
Инициация (лат. injicere — вызывать)
Образуется несколько начальных кодонов иРНК.
Нити ДНК последовательно расплетаются, освобождая место для передвигающейся РНК-полимеразы. Молекула иРНК быстро растет.
Достигая особого участка цепи ДНК - терминатора, РНК-полимераза получает сигнал к прекращению синтеза иРНК. Транскрипция завершается. Синтезированная иРНК направляется из ядра в цитоплазму.
Трансляция (от лат. translatio — перенос, перемещение)
Куда же отправляется новосинтезированная иРНК в процессе транскрипции? На следующую ступень - в процесс трансляции. Он заключается в синтезе белка на рибосоме по матрице иРНК. Последовательность кодонов иРНК переводится в последовательность аминокислот.
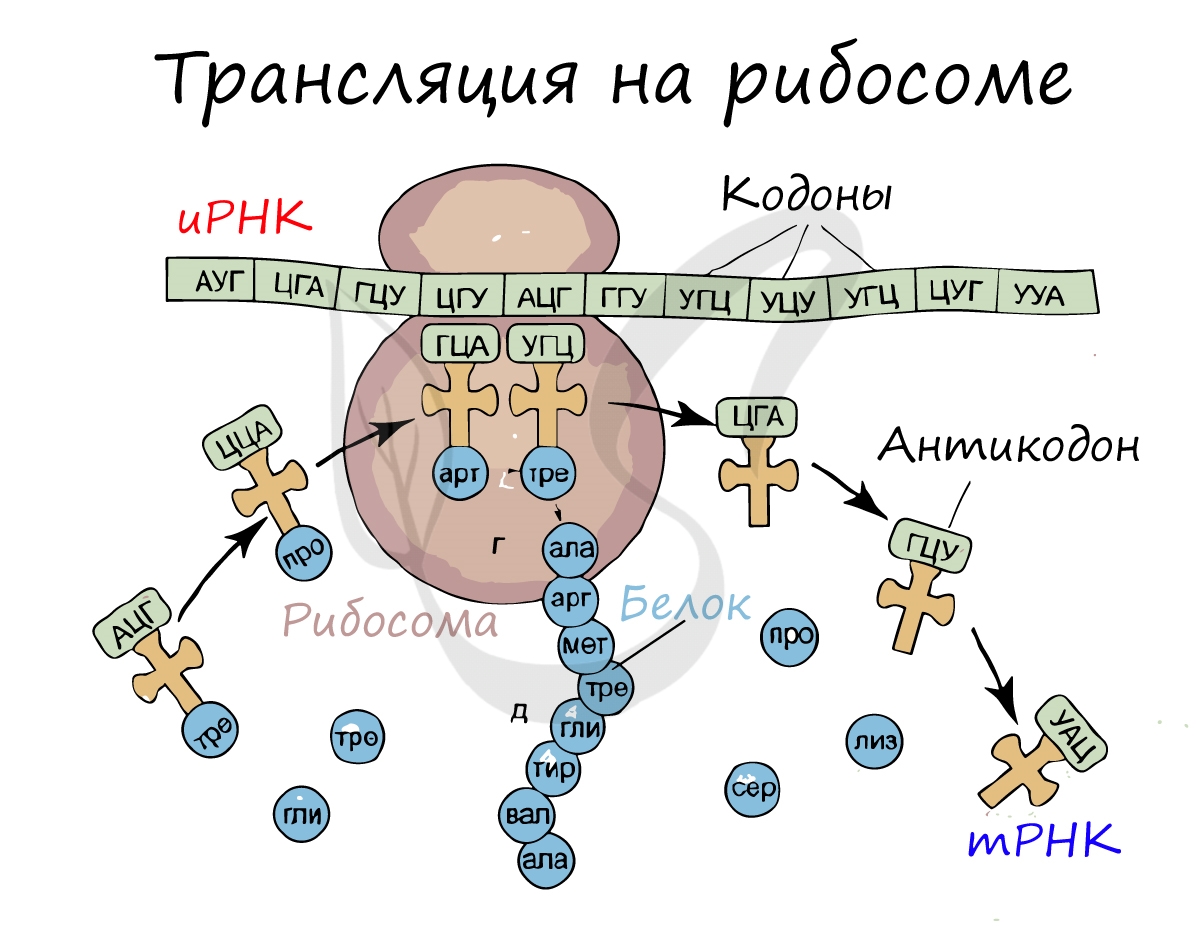
Информационная РНК (иРНК, синоним - мРНК (матричная РНК)) присоединяется к рибосоме, состоящей из двух субъединиц. Замечу, что вне процесса трансляции субъединицы рибосом находятся в разобранном состоянии.
Первый кодон иРНК, старт-кодон, АУГ оказывается в центре рибосомы, после чего тРНК приносит аминокислоту, соответствующую кодону АУГ - метионин.
Рибосома делает шаг, и иРНК продвигается на один кодон: такое в фазу элонгации происходит десятки тысяч раз. Молекулы тРНК приносят новые аминокислоты, соответствующие кодонам иРНК. Аминокислоты соединяются друг с другом: между ними образуются пептидные связи, молекула белка растет.
Доставка нужных аминокислот осуществляется благодаря точному соответствию 3 нуклеотидов (кодона) иРНК 3 нуклеотидам (антикодону) тРНК. Язык перевода между иРНК и тРНК выглядит как: А (аденин) - У (урацил), Г (гуанин) - Ц (цитозин). В основе этого также лежит принцип комплементарности.
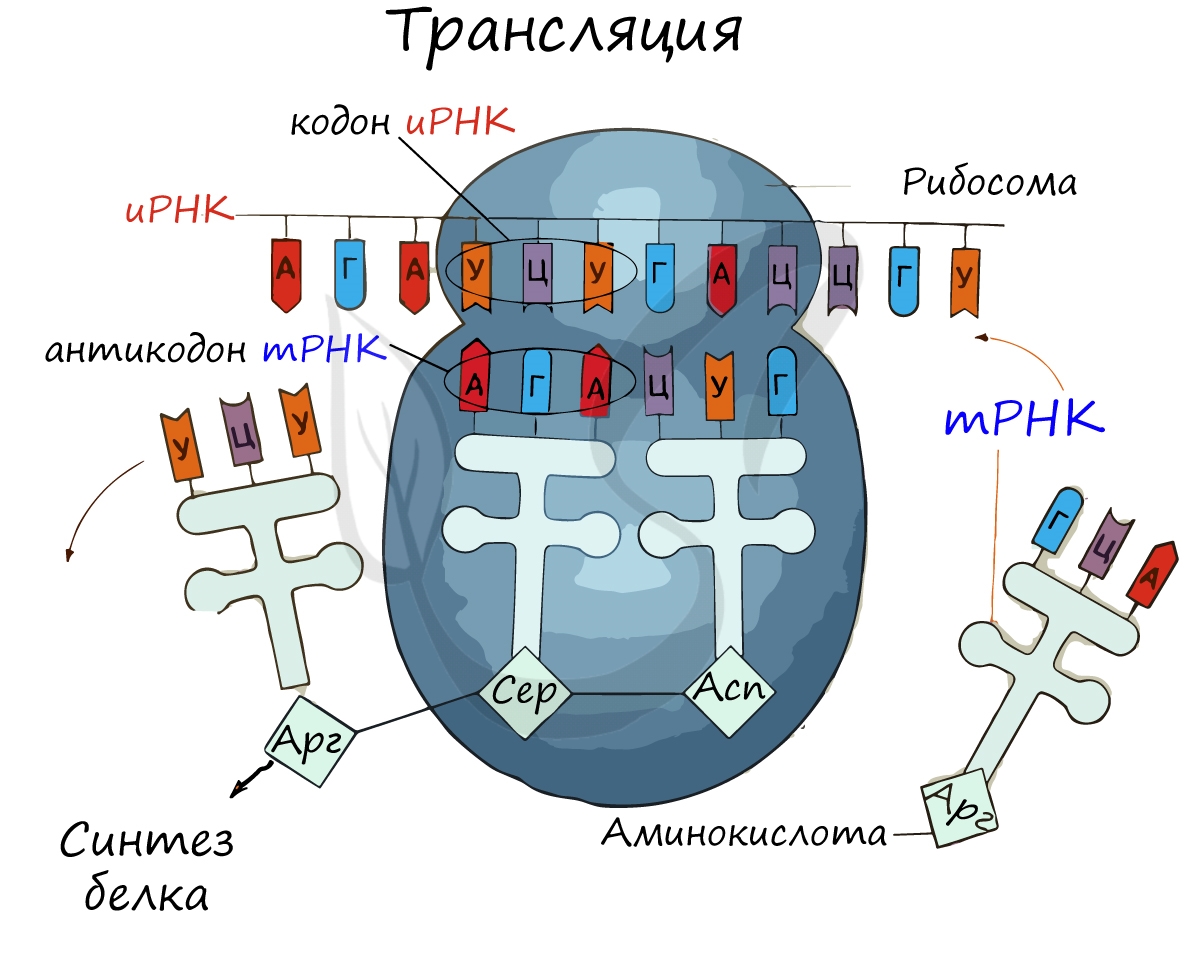
Движение рибосомы вдоль молекулы иРНК называется транслокация. Нередко в клетке множество рибосом садятся на одну молекулу иРНК одновременно - образующаяся при этом структура называется полирибосома (полисома). В результате происходит одновременный синтез множества одинаковых белков.
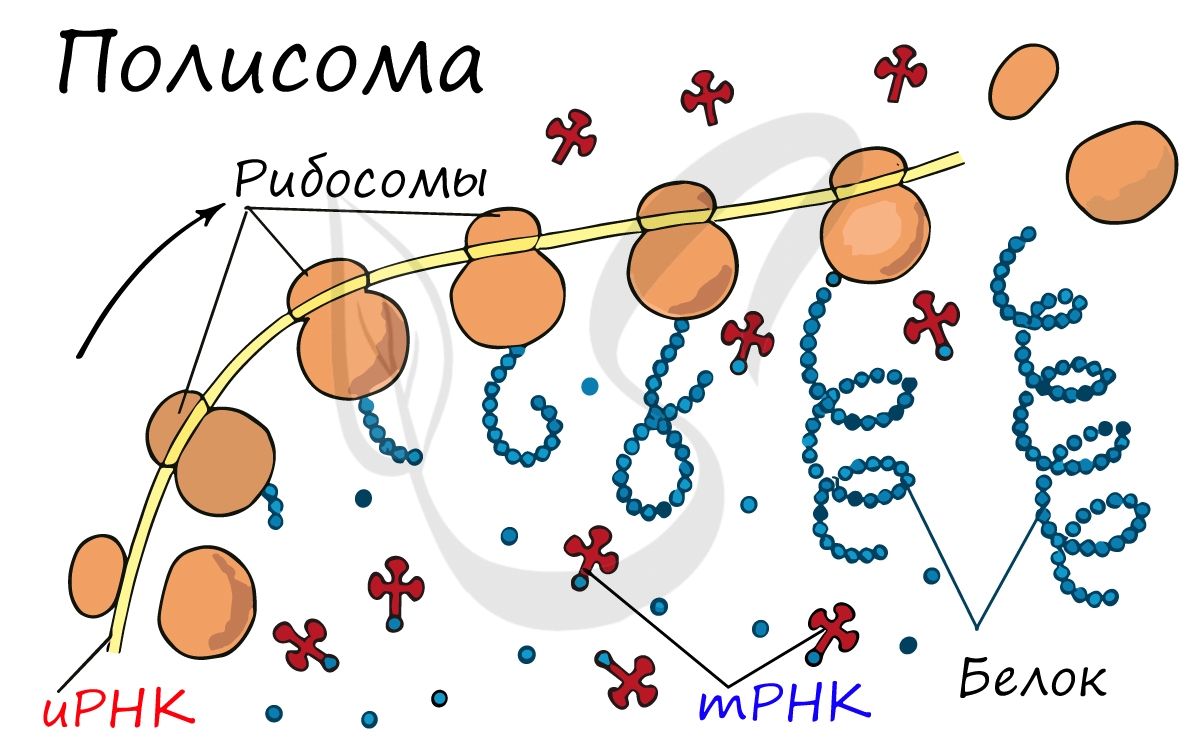
Синтез белка - полипептидной цепи из аминокислот - в определенный момент завершатся. Сигналом к этому служит попадание в центр рибосомы одного из так называемых стоп-кодонов: УАГ, УГА, УАА. Они относятся к нонсенс-кодонам (бессмысленным), которые не кодируют ни одну аминокислоту. Их функция - завершить синтез белка.
Существует специальная таблица для перевода кодонов иРНК в аминокислоты. Пользоваться ей очень просто, если вы запомните, что кодон состоит из 3 нуклеотидов. Первый нуклеотид берется из левого вертикального столбика, второй - из верхнего горизонтального, третий - из правого вертикального столбика. На пересечении всех линий, идущих от них, и находится нужная вам аминокислота :)
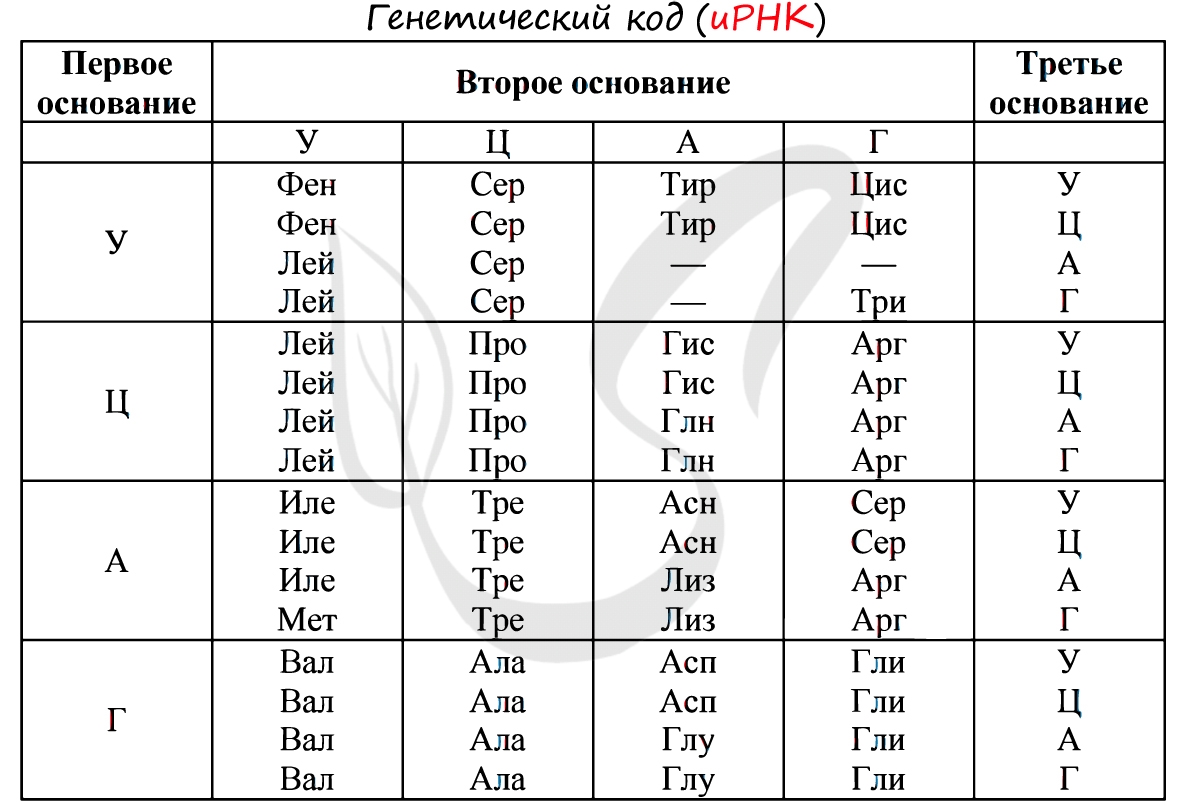
Давайте потренируемся: кодону ЦАЦ соответствует аминокислота Гис, кодону ЦАА - Глн. Попробуйте самостоятельно найти аминокислоты, которые кодируют кодоны ГЦУ, ААА, УАА.
Кодону ГЦУ соответствует аминокислота - Ала, ААА - Лиз. Напротив кодона УАА в таблице вы должны были обнаружить прочерк: это один из трех нонсенс-кодонов, завершающих синтез белка.
Примеры решения задачи №1
Без практики теория мертва, так что скорее решим задачи! В первых двух задачах будем пользоваться таблицей генетического кода (по иРНК), приведенной вверху.
"Фрагмент цепи ДНК имеет следующую последовательность нуклеотидов: ЦГА-ТГГ-ТЦЦ-ГАЦ. Определите последовательность нуклеотидов во второй цепочке ДНК, последовательность нуклеотидов на иРНК, антикодоны соответствующих тРНК и аминокислотную последовательность соответствующего фрагмента молекулы белка, используя таблицу генетического кода"

По принципу комплементарности мы нашли вторую цепочку ДНК: ГЦТ-АЦЦ-АГГ-ЦТГ. Мы использовали следующие правила при нахождении второй нити ДНК: А-Т, Т-А, Г-Ц, Ц-Г.
Вернемся к первой цепочке, и именно от нее пойдем к иРНК: ГЦУ-АЦЦ-АГГ-ЦУГ. Мы использовали следующие правила при переводе ДНК в иРНК: А-У, Т-А, Г-Ц, Ц-Г.
Зная последовательность нуклеотидов иРНК, легко найдем тРНК: ЦГА, УГГ, УЦЦ, ГАЦ. Мы использовали следующие правила перевода иРНК в тРНК: А-У, У-А, Г-Ц, Ц-Г. Обратите внимание, что антикодоны тРНК мы разделяем запятыми, в отличие кодонов иРНК. Это связано с тем, что тРНК представляют собой отдельные молекулы (в виде клеверного листа), а не линейную структуру (как ДНК, иРНК).
Пример решения задачи №2
"Известно, что все виды РНК синтезируются на ДНК-матрице. Фрагмент цепи ДНК, на которой синтезируется участок центральной петли тРНК, имеет следующую последовательность нуклеотидов: ТАГ-ЦАА-АЦГ-ГЦТ-АЦЦ. Установите нуклеотидную последовательность участка тРНК, который синтезируется на данном фрагменте, и аминокислоту, которую будет переносить эта тРНК в процессе биосинтеза белка, если третий триплет соответствует антикодону тРНК"

Обратите свое пристальное внимание на слова "Известно, что все виды РНК синтезируются на ДНК-матрице. Фрагмент цепи ДНК, на которой синтезируется участок центральной петли тРНК ". Эта фраза кардинально меняет ход решения задачи: мы получаем право напрямую и сразу синтезировать с ДНК фрагмент тРНК - другой подход здесь будет считаться ошибкой.
Итак, синтезируем напрямую с ДНК фрагмент молекулы тРНК: АУЦ-ГУУ-УГЦ-ЦГА-УГГ. Это не отдельные молекулы тРНК (как было в предыдущей задаче), поэтому не следует разделять их запятой - мы записываем их линейно через тире.
Третий триплет ДНК - АЦГ соответствует антикодону тРНК - УГЦ. Однако мы пользуемся таблицей генетического кода по иРНК, так что переведем антикодон тРНК - УГЦ в кодон иРНК - АЦГ. Теперь очевидно, что аминокислота кодируемая АЦГ - Тре.
Пример решения задачи №3
Длина фрагмента молекулы ДНК составляет 150 нуклеотидов. Найдите число триплетов ДНК, кодонов иРНК, антикодонов тРНК и аминокислот, соответствующих данному фрагменту. Известно, что аденин составляет 20% в данном фрагменте (двухцепочечной молекуле ДНК), найдите содержание в процентах остальных нуклеотидов.

Один триплет ДНК состоит из 3 нуклеотидов, следовательно, 150 нуклеотидов составляют 50 триплетов ДНК (150 / 3). Каждый триплет ДНК соответствует одному кодону иРНК, который в свою очередь соответствует одному антикодону тРНК - так что их тоже по 50.
По правилу Чаргаффа: количество аденина = количеству тимина, цитозина = гуанина. Аденина 20%, значит и тимина также 20%. 100% - (20%+20%) = 60% - столько приходится на оставшиеся цитозин и гуанин. Поскольку их процент содержания равен, то на каждый приходится по 30%.
Теперь мы украсили теорию практикой. Что может быть лучше при изучении новой темы? :)
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Читайте также: