
Типы фотосинтеза с3 с4 кратко
Обновлено: 01.07.2024
-главный путь ассимиляции СО2, , представляет собой циклический процесс, в который вводится СО2 и из которого выходит углевод.
Процесс можно разделить на три фазы:
первая фаза - непосредственная фиксация углекислого газа – карбоксилирование: углекислый газ, связываясь с рибулозобифосфатом (фосфатом сахара с пятью атомами С - РДФ), образует две молекулы фосфоглицерата. Эту реакцию катализирует рибулозобифосфат-карбоксилаза;
вторая фаза - восстановление. Фосфоглицерат при участии NADPН (восстановитель) и АТР (донор энергии) восстанавливается до 3-фосфоглицеральдегида (ФГА).
третья фаза – образование продуктов фотосинтеза.
четвертая стадия- восстановление первоначальных реагентов
Для того чтобы растение могло акцептировать новую молекулу углекислого газа, необходимо иметь РДФ, основной акцептор углекислого газа. РДФ образуется из ФГА в результате цепи реакций, в процессе которых образуются пяти- и семиуглеродные сахара. Надо отметить, что основная масса ФГА идет именно на восстановление нужного количества РДФ: из 12 образовавшихся молекул ФГА только две идут на образование продуктов фотосинтеза.
С окончанием этой фазы цикл замыкается. Ферменты цикла находятся в строме хлоропласта, а рибулозобифосфат-карбоксилаза - на наружной стороне тилакоидных мембран.
Суммарное уравнение синтеза глюкозы в цикле Кальвина:
Фотосинтез по С3-пути всегда сопровождается фотодыханием – процессом потребления на свету кислорода и выделения СО2. фотодыхание – довольно затратный процесс, т.к. он не сопровождается синтезом АТФ, а, наоборот забирает энергию для восстановления кислорода. Более того, 50% углерода, используемого при фотосинтезе у С3-растений, восстанавливается до СО2.
Фотодыхание – это стимулированное светом быстрое высвобождение СО2 листьями, этот процесс протекает только на свету и сопровождается поглощением О2. Такое изменение хода реакции фотосинтеза объясняется тем, что участвующий в ней фермент обладает двойной каталитической активностью – по отношению к углекислому газу и кислороду.
Этот фермент называется рибулозо-1,5-бифосфаткарбоксилаза-оксигеназа (РуБФ-карбоксилаза, или Rubisco). Он составляет около 50% всех растворимых белков в листьях и потому может считаться самым распространенным белком в природе.
В норме этот фермент связывает СО2 в цикле Кальвина, однако он может действовать также как оксигенеза (т.е. связывается с О2), катализируя окисление рибулозофосфата до 3-фосфоглицериновой кислоты (идет в цикл Кальвина) и 2-фосфогликолевой кислоты. Далее фосфат гликолевой кислоты дефосфорилируется и гликолевая кислота транспортируется в пероксисомы, где окисляется до глиоксилевой кислоты и аминируется с получением глицина. В митохондриях из двух молекул глицина образуется серин и углекислый газ. Серин может использоваться для синтеза белка, либо превращается в 3-фосфоглицериновую кислоту и возвращается в цикл Кальвина.
При высоких концентрациях СО2 и низких О2 преобладает карбоксилирование, соответственно при низких концентрациях СО2 - окисление. Повышение температуры также ускоряет окисление.
Фотодыхание снижает эффективность фотосинтеза, приводит к потерям ассимилированного углерода, однако имеет некоторое синтетическое значение.
На ранних этапах развития жизни, когда в атмосфере было мало кислорода, рубиско заняло ключевую позицию в фотосинтезе, и ее оксигеназная функция не доставляла проблем. По мере увеличения содержания кислорода потери на фотодыхание нарастали, и у ряда растений возникли механизмы активной доставки к месту работы рубиско углекислого газа (С4 и САМ-растения), увеличивающее долю её карбоксилазной активности до 100%.
Таким образом, на первых этапах фиксации углерода имеет место конкуренция между двумя процессами – фиксацией углерода и фотодыханием.
С4 тип фотосинтеза
Для С4 растений характерна особая морфология листа и они содержат хлоропласты двух различных типов.
Хлоропласты находятся в клетках листа, расположенных вокруг сосудистого пучка двумя концентрическими слоями:
внутренний слой –клетки обкладки сосудистого пучка, хлоропласты довольно крупные, не имеют, как правило гран и содержат много зерен крахмала.
внешний – клетки мезофилла, хлоропласты мезофилла довольно мелкие расположены в клетке беспорядочно, содержат стопки гран и небольшое количество зерен крахмала.
У С4-растений фиксация СО2 может осуществляться двумя путями:
обычный цикл Кальвина, функционирующий в клетках обкладки. Первичный продукт фиксации СО2 – фосфоглицериновая кислота – С3
САМ-фотосинтез Растения с данным типом фотосинтеза являются в основном
суккулентами, которые обитают в засушливых, безводных областях.
Для этих растений характерны следующие особенности. 1. Их устьица обычно открыты ночью (т.е. в темноте) и закрыты в течение дня. 2. Фиксация углекислого газа происходит в темное время суток с образованием кислот, содержащих четыре атома углерода – оксалоацетата и малата.
3. Малат запасается в больших вакуолях, которые характерны для клеток МОКТ-растений. 4. В светлое время суток малат переносится в цитоплазму и там декарбоксилируется, образуя пируват и СО2, которая поступает в цикл Кальвина, где превращается в сахарозу или запасной углевод глюкан. 5. В темновой период суток часть запасенного глюкана распадается с образованием молекул-акцепторов для темновой фиксации углекислого газа
Таким образом у МОКТ-растений существует суточный ритм: ночью содержание запасного глюкана падает и содержание яблочной кислоты повышается, а днем происходят противоположные изменения. При достаточном количестве воды эти растения могут вести себя как С3 растения

Усвоение углекислого газа из солнечного света для процесса фотосинтеза, а затем превращение его в глюкозу (энергию), синтезирующую различные продукты, является ключевым отличием между ними. Таким образом, во время фиксации CO2, когда фотосинтетические растения производят 3-фосфоглицериновую кислоту (PGA) или 3-углеродную кислоту в качестве первого продукта, это называется C3-путь .
Но когда фотосинтезирующее растение, прежде чем идти по пути С3, производит щавелевоуксусную кислоту (ОАА) или 4 -углеродное соединение в качестве своего первого стабильного продукта, его называют С4 или путь Хэтча и Шлака . Но когда растения поглощают энергию солнечного света в дневное время и используют эту энергию для усвоения или фиксации углекислого газа в ночное время, это называется метаболизмом толстой кислоты или САМ .
За этими процедурами следуют растения, определенные виды бактерий и водорослей для производства энергии, независимо от среды их обитания. Синтез энергии с использованием углекислого газа и воды в качестве основного источника для получения питательных веществ из воздуха и воды называется фотосинтезом. Это основной процесс для живого существа, которое производит пищу самостоятельно
В этом материале мы рассмотрим существенное различие между тремя типами путей, по которым следуют растения, и несколькими микроорганизмами, и небольшое описание о них.
Сравнительная таблица
| Основа для сравнения | C3 путь | C4 путь | CAM |
|---|---|---|---|
| Определение | Такие растения, чей первый продукт после ассимиляции углерода от солнечного света - это 3-углеродная молекула или 3-фосфоглицериновая кислота для производство энергии называется растениями С3, а путь называется путём С3. Это чаще всего используется растениями. | Растения в тропической зоне преобразуют энергию солнечного света в молекулу углерода С4 или в щавелево-уксусную кислоту, что происходит до цикла С3 и затем он далее преобразуется в энергию, называется растениями С4, а путь называется путём С4. Это более эффективно, чем путь C3. | Растения, которые накапливают энергию от солнца, а затем преобразуют ее в энергию в течение ночи, следуют за CAM или crassulacean кислотой. метаболизм. |
| Клетки участвуют | Клетки мезофилла. | Мезофилловая клетка, пучки оболочек клеток. | И С3, и С4 в одних и тех же клетках мезофилла. |
| пример | Подсолнечник, шпинат, фасоль, рис, хлопок. | Сахарный тростник, сорго и кукуруза. | Кактусы, орхидеи. |
| Можно увидеть в | Все фотосинтезирующие растения. | В тропических растениях | Полузасушливое состояние. |
| Типы растений, использующих этот цикл | Мезофитный, гидрофитный, ксерофитный. | Мезофильных. | Ксерофитные. |
| Фотодыхание | Присутствует в высоком темпе. | Не легко обнаружить. | Обнаруживается днем. |
| Для производства глюкозы | Требуется 12 NADPH и 18 ATP. | Требуется 12 NADPH и 30 ATP. | Требуется 12 NADPH и 39 ATP. |
| Первый стабильный продукт | 3-фосфоглицерат (3-PGA). | Оксалоацетат (ОАА). | Оксалоацетат (ОАА) ночью, 3 ПГА в дневное время. |
| Кальвин циклоператор | В одиночестве. | Наряду с циклом Хэтча и Слэка. | С3 и хэтч и слабый цикл. |
| Оптимальная температура для фотосинтеза | 15-25 ° C | 30-40 ° С | > 40 градусов ° C |
| Карбоксилирующий фермент | RuBP карбоксилаза. | В мезофилле: PEP карбоксилаза. В связке оболочка: RuBP карбоксилаза. | В темноте: ПКП-карбоксилаза. В свете: RUBP карбоксилаза. |
| Соотношение CO2: ATP: NADPH2 | 1: 3: 2 | 1: 5: 2 | 1: 6, 5: 2 |
| Начальный акцептор СО2 | Рибулозо-1, 5-biphophate (RuBP). | Фосфоенолпируват (ПКП). | Фосфоенолпируват (ПКП). |
| Анатомия Кранца | Отсутствует. | Настоящее время. | Отсутствует. |
| Точка компенсации CO2 (промилле) | 30-70. | 6-10. | 0-5 в темноте. |
Определение пути С3 или цикла Кальвина.
Растения С3 известны как растения холодного сезона или умеренные . Они лучше всего растут при оптимальной температуре от 65 до 75 ° F с температурой почвы, подходящей на уровне 40-45 ° F. Эти типы растений демонстрируют меньшую эффективность при высокой температуре .
Основным продуктом растений C3 является 3-углеродная кислота или 3-фосфоглицериновая кислота (PGA) . Это считается первым продуктом при фиксации углекислого газа. Путь C3 проходит в три этапа: карбоксилирование, восстановление и регенерация.
Растения С3 превращаются в СО2 непосредственно в хлоропласте. С помощью рибулозобифосфаткарбоксилазы (RuBPcase) получают две молекулы 3-углеродной кислоты или 3-фосфоглицериновой кислоты . Этот 3-фосфоглицерик оправдывает название пути как C3.
На другом этапе NADPH и ATP фосфорилируют с получением 3-PGA и глюкозы. И затем цикл снова начинается с регенерации RuBP.
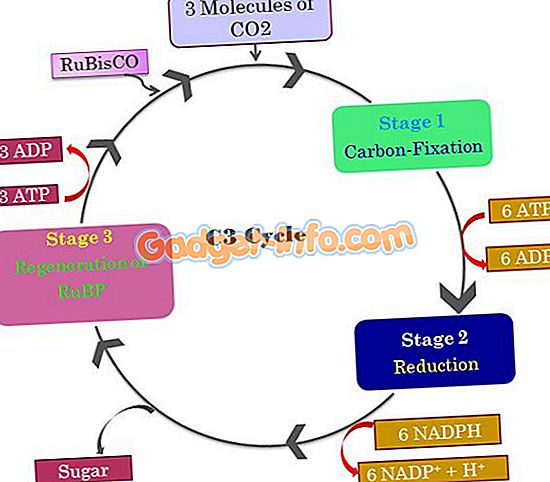
Путь C3 является одностадийным процессом, происходящим в хлоропласте. Эта органелла действует как хранилище солнечной энергии. Из общего количества растений, присутствующих на земле, 85 процентов используют этот путь для производства энергии.
Растения С3 могут быть многолетними или однолетними. Они очень белковые, чем растения C4. Примерами однолетних растений С3 являются пшеница, овес и рожь, а многолетние растения включают в себя феску, райграс и фруктовый сад. Растения С3 содержат больше белка, чем растения С4.
Определение пути С4 или пути Люка и Слака.
Растения, особенно в тропическом регионе, следуют по этому пути. Перед циклом Calvin или C3 некоторые растения следуют по пути C4 или Hatch and Slack. Это двухстадийный процесс, в котором получают щавелевоуксусную кислоту (ОАА), которая представляет собой 4-углеродное соединение . Это происходит в клетках оболочки мезофилла и связки, присутствующих в хлоропласте.
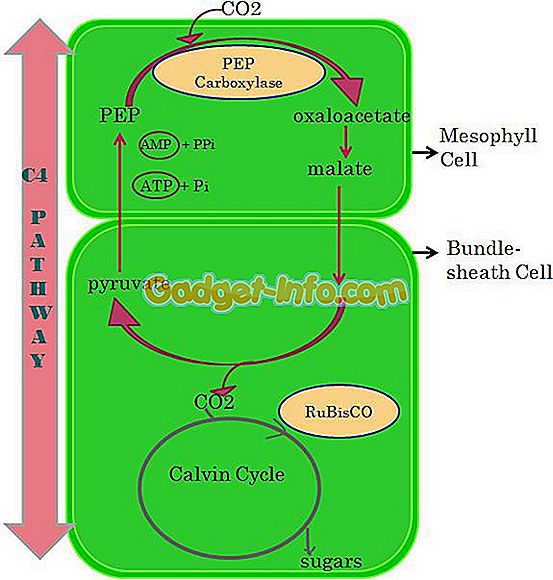
Когда 4-углеродное соединение произведено, оно отправляется в ячейку оболочки пучка, здесь молекула 4-углерода дополнительно расщепляется на диоксид углерода и соединение 3-кабона. В конце концов, путь C3 начинает генерировать энергию, где 3-углеродное соединение выступает в качестве предшественника.
Растения С4 также известны как теплые или тропические растения . Они могут быть многолетними или однолетними. Идеальная температура для этих растений - 90-95 ° F. Растения С4 гораздо эффективнее используют азот и собирают углекислый газ из почвы и атмосферы. Содержание белка низкое по сравнению с растениями С3.
Эти растения получили свое название от продукта под названием оксалоацетат, который является 4-угольной кислотой. Примерами многолетних растений C4 являются индийская трава, бермудские острова, просевная трава, крупный синий, а однолетние растения C4 - это суданга, кукуруза, жемчужное просо.
Определение растений CAM
Примечательное замечание, которое отличает этот процесс от вышеупомянутых двух, заключается в том, что при этом типе фотосинтеза организм поглощает энергию солнечного света в дневное время и использует эту энергию в ночное время для ассимиляции углекислого газа.
Это своего рода адаптация во время периодической засухи. Этот процесс допускает обмен газами в ночное время, когда температура воздуха ниже, и происходит потеря водяного пара.
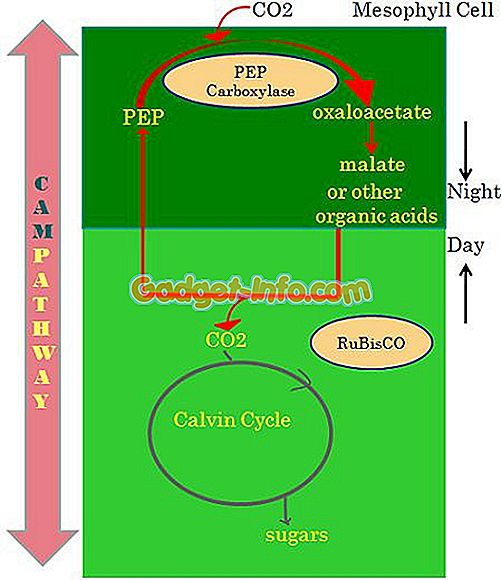
Около 10% сосудистых растений адаптированы к фотосинтезу САМ, но в основном обнаруживаются у растений, выращенных в засушливых районах. Растения, такие как кактус и молочай, являются примерами. Даже орхидеи и бромелии приспособили этот путь из-за нерегулярного водоснабжения.
В дневное время малат декарбоксилируется для обеспечения CO2 для фиксации цикла Бенсона-Кальвина в закрытых устьицах. Главной особенностью растений CAM является ассимиляция ночью CO2 в яблочную кислоту, хранящуюся в вакуоле. PEP карбоксилаза играет основную роль в производстве малата.
Основные отличия растений C3, C4 и CAM.
Выше мы обсудим процедуру получения энергии этих различных типов, ниже мы обсудим ключевые различия между тремя:
- Путь C3 или растения C3 можно определить как те виды растений, у которых первым продуктом после ассимиляции углерода от солнечного света является 3-углеродная молекула или 3-фосфоглицериновая кислота для производства энергии. Это наиболее часто используется растениями; В то время как растения в тропической зоне преобразуют энергию солнечного света в молекулу углерода С4 или щавелевоуксусную кислоту, этот цикл происходит до цикла С3, а затем с помощью ферментов он осуществляет дальнейший процесс получения питательных веществ, называется растениями С4, а путь называется как путь С4. Этот путь более эффективен, чем путь C3. С другой стороны, растения, которые накапливают энергию солнца в дневное время, а затем преобразуют ее в энергию ночью, следуют за метаболизмом CAM или crassulacean кислотой .
- Клетки, участвующие в пути C3, представляют собой клетки мезофилла, а клетки, участвующие в пути C4, представляют собой клетки мезофилла, клетки оболочки пучка, но CAM следует за C3 и C4 в одних и тех же клетках мезофилла.
- Примером С3 являются Подсолнечник, Шпинат, Бобы, Рис, Хлопок, тогда как примером растений С4 является Сахарный тростник, Сорго и Кукуруза, а Кактусы, орхидеи, являются примерами растений САМ.
- C3 можно увидеть во всех фотосинтезирующих растениях, в то время как за C4 следуют тропические растения, а CAM - полузасушливые растения.
- Типы растений, использующих цикл C3, являются мезофитными, гидрофитными, ксерофитными, но C4 следует за мезофитными растениями, а Xerophytic следует за CAM.
- Фотодыхание присутствует с большей скоростью, но его трудно обнаружить в C4 и CAM.
- 12 NADPH и 18 ATP в цикле C3; 12 НАДФН и 30 АТФ в С4 и 12 НАДФН и 39 АТФ необходимы для производства глюкозы.
- 3-фосфоглицерат (3-PGA) является первым стабильным продуктом пути C3; Оксалоацетат (ОАА) для пути С4 и Оксалоацетат (ОАА) ночью, 3 PGA в дневное время в САМ.
- Оптимальная температура для фотосинтеза в С3 составляет 15-25 ° С; 30-40 ° C в растениях C4 и> 40 ° C в CAM
- Карбоксилирующий фермент представляет собой RuBP-карбоксилазу в растениях C3, но в растениях C4 это PEP-карбоксилаза (в мезофилле) и RuBP-карбоксилазу (в оболочке пучка), тогда как в CAM это PEP-карбоксилаза (в темноте) и RuBP-карбоксилаза (в свете).
- Соотношение CO2: ATP: NADPH2 1: 3: 2 в C3, 1: 5: 2 в C4 и 1: 6, 5: 2 в CAM.
- Исходным акцептором CO2 является рибулозо-1, 5-бифосфат (RuBP) в пути C3 и фосфоенолпируват (PEP) в C4 и CAM.
- Анатомия Кранца присутствует только в пути С4 и отсутствует у растений С3 и САМ.
- Точка компенсации CO2 (ppm) составляет 30-70 для установки C3; 6-10 в C4 растениях и 0-5 в темноте в CAM.
Вывод
Мы все знаем о том, что растения готовят пищу в процессе фотосинтеза. Они превращают атмосферный углекислый газ в растительную пищу или энергию (глюкозу). Но поскольку растения растут в разных местах обитания, они имеют разные атмосферные и климатические условия; они отличаются в процессе получения энергии.
Как и в случае, когда пути C4 и CAM являются двумя адаптациями, возникшими в результате естественного отбора, для выживания растений с высокой температурой и засушливой области. Таким образом, мы можем сказать, что это три различных биохимических метода растений для получения энергии, и C3 является наиболее распространенным среди них.
Фотосинтез — синтез органических веществ из углекислого газа и воды с обязательным использованием энергии света:

Хлорофиллы поглощают красный и сине-фиолетовый свет, отражают зеленый и поэтому придают растениям характерную зеленую окраску. Молекулы хлорофилла в мембранах тилакоидов организованы в фотосистемы. У растений и синезеленых водорослей имеются фотосистема-1 и фотосистема-2, у фотосинтезирующих бактерий — фотосистема-1. Только фотосистема-2 может разлагать воду с выделением кислорода и отбирать электроны у водорода воды.
Фотосинтез — сложный многоступенчатый процесс; реакции фотосинтеза подразделяют на две группы: реакции световой фазы и реакции темновой фазы.
Световая фаза
Эта фаза происходит только в присутствии света в мембранах тилакоидов при участии хлорофилла, белков-переносчиков электронов и фермента — АТФ-синтетазы. Под действием кванта света электроны хлорофилла возбуждаются, покидают молекулу и попадают на внешнюю сторону мембраны тилакоида, которая в итоге заряжается отрицательно. Окисленные молекулы хлорофилла восстанавливаются, отбирая электроны у воды, находящейся во внутритилакоидном пространстве. Это приводит к распаду или фотолизу воды:
Ионы гидроксила отдают свои электроны, превращаясь в реакционноспособные радикалы •ОН:
Радикалы •ОН объединяются, образуя воду и свободный кислород:
2Н + + 2е — + НАДФ → НАДФ·Н2.
Таким образом, в световую фазу происходит фотолиз воды, который сопровождается тремя важнейшими процессами: 1) синтезом АТФ; 2) образованием НАДФ·Н2; 3) образованием кислорода. Кислород диффундирует в атмосферу, АТФ и НАДФ·Н2 транспортируются в строму хлоропласта и участвуют в процессах темновой фазы.

1 — строма хлоропласта; 2 — тилакоид граны.
Темновая фаза
Эта фаза протекает в строме хлоропласта. Для ее реакций не нужна энергия света, поэтому они происходят не только на свету, но и в темноте. Реакции темновой фазы представляют собой цепочку последовательных преобразований углекислого газа (поступает из воздуха), приводящую к образованию глюкозы и других органических веществ.
Кроме глюкозы, в процессе фотосинтеза образуются другие мономеры сложных органических соединений — аминокислоты, глицерин и жирные кислоты, нуклеотиды. В настоящее время различают два типа фотосинтеза: С3- и С4-фотосинтез.
С3-фотосинтез

Фотодыхание

Фотодыхание:
1 — хлоропласт; 2 — пероксисома; 3 — митохондрия.
Это светозависимое поглощение кислорода и выделение углекислого газа. Еще в начале прошлого века было установлено, что кислород подавляет фотосинтез. Как оказалось, для РиБФ-карбоксилазы субстратом может быть не только углекислый газ, но и кислород:
О2 + РиБФ → фосфогликолат (2С) + ФГК (3С).
Фермент при этом называется РиБФ-оксигеназой. Кислород является конкурентным ингибитором фиксации углекислого газа. Фосфатная группа отщепляется, и фосфогликолат становится гликолатом, который растение должно утилизировать. Он поступает в пероксисомы, где окисляется до глицина. Глицин поступает в митохондрии, где окисляется до серина, при этом происходит потеря уже фиксированного углерода в виде СО2. В итоге две молекулы гликолата (2С + 2С) превращаются в одну ФГК (3С) и СО2. Фотодыхание приводит к понижению урожайности С3-растений на 30–40% (С3-растения — растения, для которых характерен С3-фотосинтез).
С4-фотосинтез
С4-фотосинтез — фотосинтез, при котором первым продуктом являются четырехуглеродные (С4) соединения. В 1965 году было установлено, что у некоторых растений (сахарный тростник, кукуруза, сорго, просо) первыми продуктами фотосинтеза являются четырехуглеродные кислоты. Такие растения назвали С4-растениями. В 1966 году австралийские ученые Хэтч и Слэк показали, что у С4-растений практически отсутствует фотодыхание и они гораздо эффективнее поглощают углекислый газ. Путь превращений углерода в С4-растениях стали называть путем Хэтча-Слэка.
Для С4-растений характерно особое анатомическое строение листа. Все проводящие пучки окружены двойным слоем клеток: наружный — клетки мезофилла, внутренний — клетки обкладки. Углекислый газ фиксируется в цитоплазме клеток мезофилла, акцептор — фосфоенолпируват (ФЕП, 3С), в результате карбоксилирования ФЕП образуется оксалоацетат (4С). Процесс катализируется ФЕП-карбоксилазой. В отличие от РиБФ-карбоксилазы ФЕП-карбоксилаза обладает большим сродством к СО2 и, самое главное, не взаимодействует с О2. В хлоропластах мезофилла много гран, где активно идут реакции световой фазы. В хлоропластах клеток обкладки идут реакции темновой фазы.
Оксалоацетат (4С) превращается в малат, который через плазмодесмы транспортируется в клетки обкладки. Здесь он декарбоксилируется и дегидрируется с образованием пирувата, СО2 и НАДФ·Н2.
Пируват возвращается в клетки мезофилла и регенерирует за счет энергии АТФ в ФЕП. СО2 вновь фиксируется РиБФ-карбоксилазой с образованием ФГК. Регенерация ФЕП требует энергии АТФ, поэтому нужно почти вдвое больше энергии, чем при С3-фотосинтезе.
С4-фотосинтез:
1 — клетка мезофилла; 2 — клетка обкладки проводящего пучка.
Значение фотосинтеза
Купить проверочные работы
и тесты по биологии




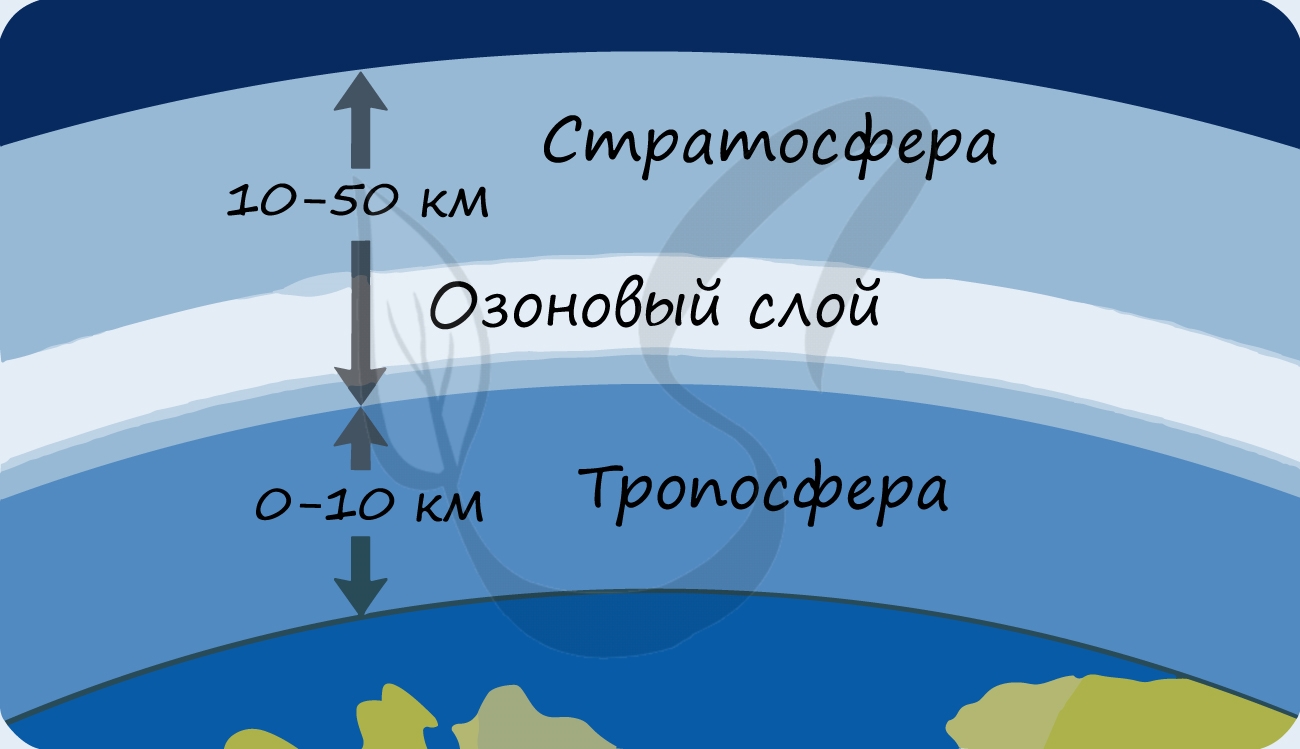
Благодаря фотосинтезу, ежегодно из атмосферы поглощаются миллиарды тонн углекислого газа, выделяются миллиарды тонн кислорода; фотосинтез является основным источником образования органических веществ. Из кислорода образуется озоновый слой, защищающий живые организмы от коротковолновой ультрафиолетовой радиации.
При фотосинтезе зеленый лист использует лишь около 1% падающей на него солнечной энергии, продуктивность составляет около 1 г органического вещества на 1 м 2 поверхности в час.
Хемосинтез
Синтез органических соединений из углекислого газа и воды, осуществляемый не за счет энергии света, а за счет энергии окисления неорганических веществ, называется хемосинтезом. К хемосинтезирующим организмам относятся некоторые виды бактерий.
Нитрифицирующие бактерии окисляют аммиак до азотистой, а затем до азотной кислоты (NH3 → HNO2 → HNO3).
Железобактерии превращают закисное железо в окисное (Fe 2+ → Fe 3+ ).
Серобактерии окисляют сероводород до серы или серной кислоты (H2S + ½O2 → S + H2O, H2S + 2O2 → H2SO4).
В результате реакций окисления неорганических веществ выделяется энергия, которая запасается бактериями в форме макроэргических связей АТФ. АТФ используется для синтеза органических веществ, который проходит аналогично реакциям темновой фазы фотосинтеза.
Хемосинтезирующие бактерии способствуют накоплению в почве минеральных веществ, улучшают плодородие почвы, способствуют очистке сточных вод и др.
По типу питания живые организмы делятся на автотрофы, гетеротрофы и миксотрофы. Автотрофы (греч. αὐτός — сам + τροφ - пища) - организмы, которые самостоятельно способны синтезировать органические вещества из неорганических. Гетеротрофы (греч. ἕτερος - иной + τροφή - пища) - организмы, использующие для питания готовые органические вещества.
Наконец, миксотрофы (греч. μῖξις - смешение + τροφή - пища) - организмы, которые могут использовать как гетеротрофный, так и автотрофный способ питания. К примеру, эвглена зеленая на свету начинает фотосинтезировать, а в темноте питается гетеротрофно.

Фотосинтез
Фотосинтез (греч. φῶς - свет и σύνθεσις - синтез) - сложный химический процесс преобразования энергии квантов света в энергию химических связей. В результате фотосинтеза происходит синтез органических веществ из неорганических.

Этот процесс уникален и происходит только в растительных клетках, а также у некоторых бактерий. Фотосинтез осуществляется при участии хлорофилла (греч. χλωρός - зелёный и φύλλον - лист) - зеленого пигмента, окрашивающего органы растений в зеленый цвет. Существуют и другие вспомогательные пигменты, которые вместе с хлорофиллом выполняют светособирающую или светозащитную функции.
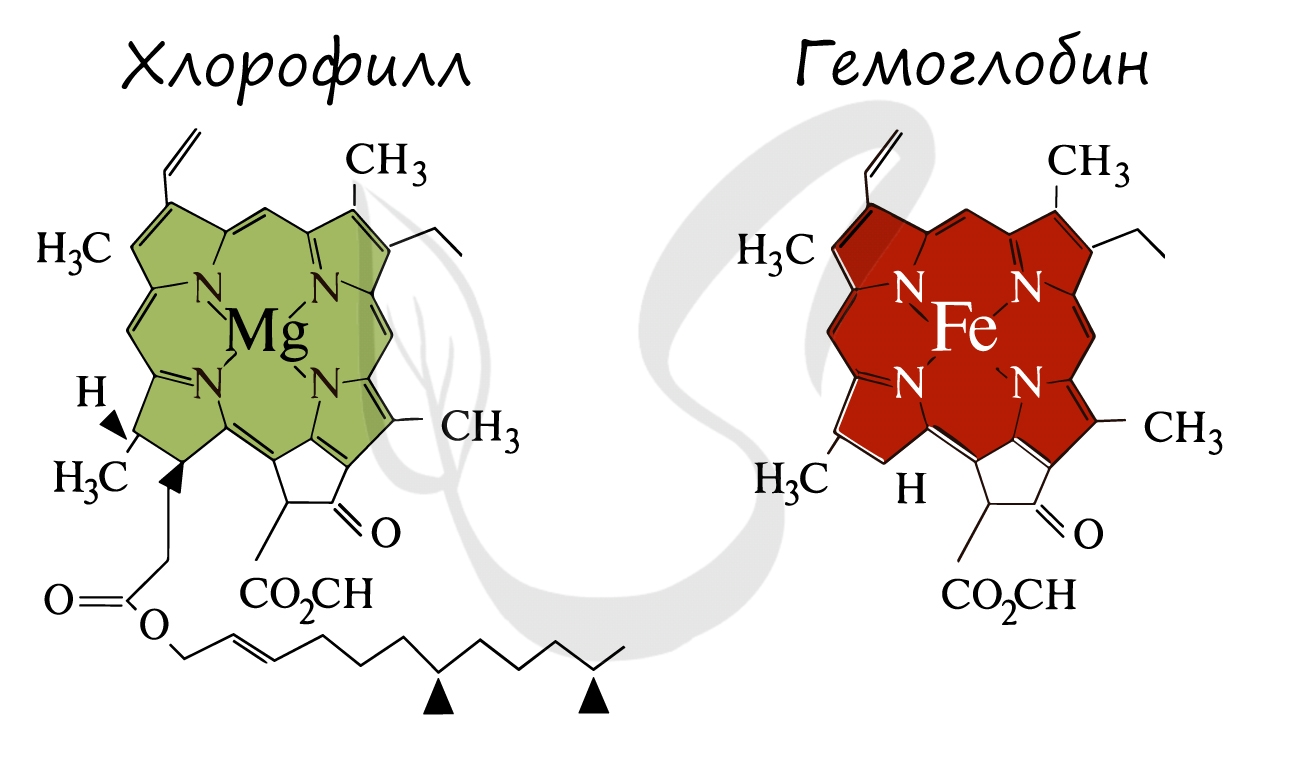
Ниже вы увидите сравнение строения хлорофилла и гемоглобина. Обратите внимание, что в центре молекулы хлорофилла находится ион Mg.
В высшей степени гениально значение процесса фотосинтеза подчеркнул русский ученый К.А. Тимирязев: "Все органические вещества, как бы они ни были разнообразны, где бы они ни встречались, в растении ли, в животном или человеке, прошли через лист, произошли от веществ, выработанных листом. Вне листа или, вернее, вне хлорофиллового зерна в природе не существует лаборатории, где бы выделялось органическое вещество. Во всех других органах и организмах оно превращается, преобразуется, только здесь оно образуется вновь из вещества неорганического"
Более подробно мы обсудим значение фотосинтеза в завершение этой статьи. Фотосинтез состоит из двух фаз: светозависимой (световой) и светонезависимой (темновой). Я рекомендую использовать названия светозависимая и светонезависимая, так как они способствуют более глубокому (и правильному!) пониманию фотосинтеза.
Светозависимая фаза (световая)
Эта фаза происходит только на свету на мембранах тилакоидов в хлоропластах. В ней принимают участие различные ферменты, белки-переносчики, молекулы АТФ-синтетазы и зеленый пигмент хлорофилл.
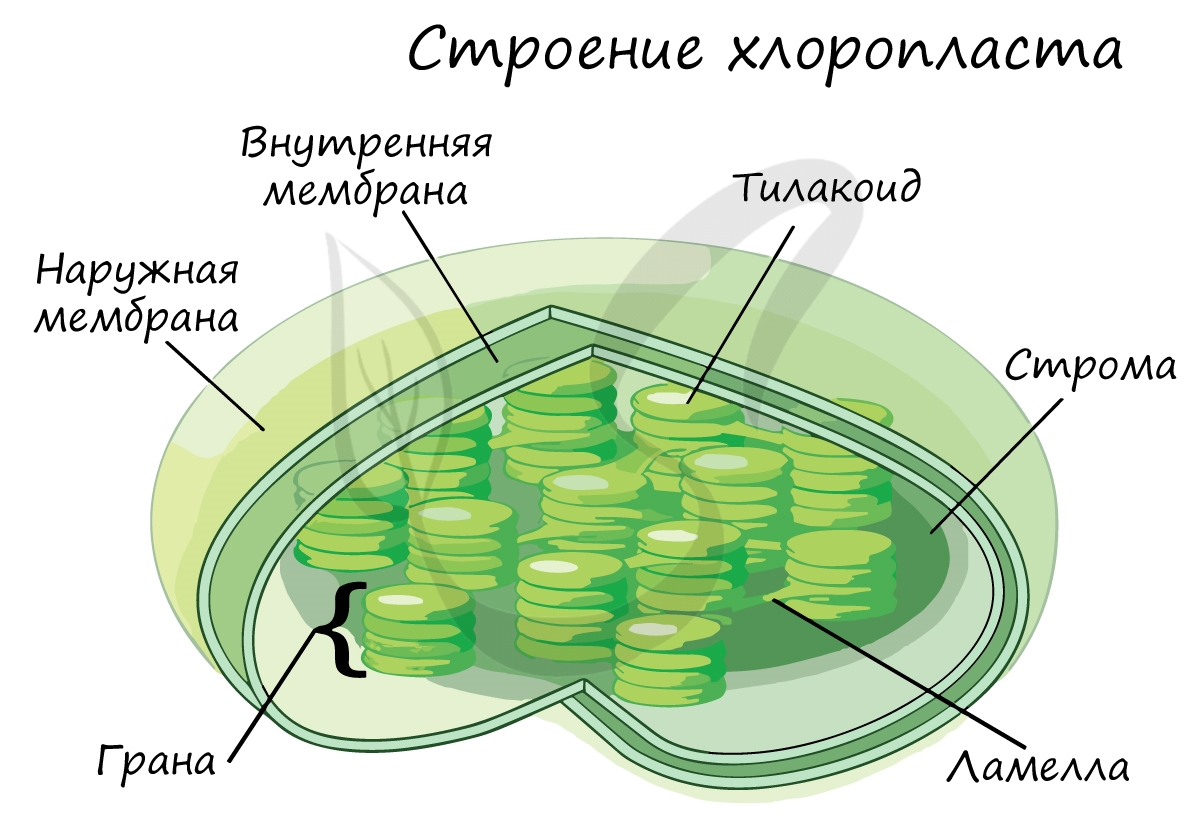
Хлорофилл выполняет две функции: поглощения и передачи энергии. При воздействии кванта света хлорофилл теряет электрон, переходя в возбужденное состояние. С помощью переносчиков электроны скапливаются с наружной поверхности мембраны тилакоидов, тем временем внутри тилакоида происходит фотолиз воды (разложение под действием света):
Гидроксид-ионы отдают лишний электрон, превращаясь в реакционно способные радикалы OH, которые собираются вместе и образуют молекулу воды и свободный кислород (это побочный продукт, который в дальнейшем удаляется в ходе газообмена).
Образовавшиеся при фотолизе воды протоны (H + ) скапливаются с внутренней стороны мембраны тилакоидов, а электроны - с внешней. В результате по обе стороны мембраны накапливаются противоположные заряды.
При достижении критической разницы, часть протонов проталкивается на внешнюю сторону мембраны через канал АТФ-синтетазы. В результате этого выделяется энергия, которая может быть использована для фосфорилирования молекул АДФ:
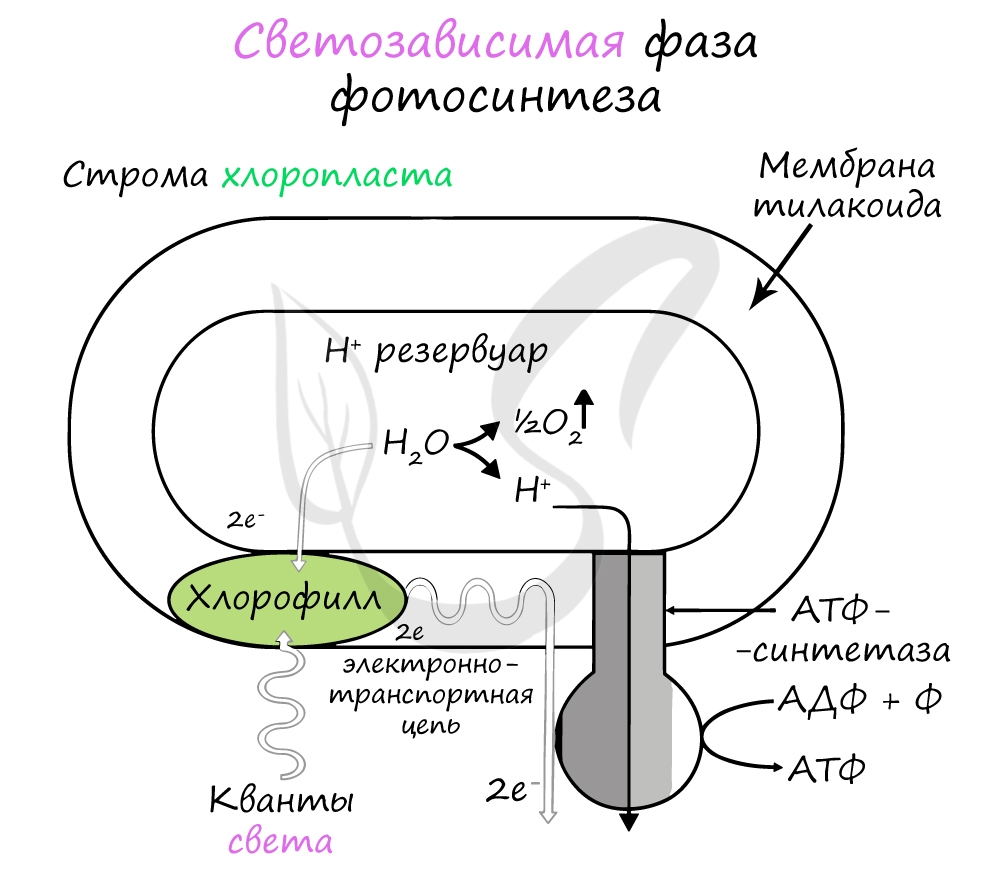
Протоны, попав на поверхность мембраны тилакоидов, соединяются с электронами и образуют атомарный водород, который используется для восстановления молекулы-переносчика НАДФ (никотинамиддинуклеотидфосфат). Благодаря этому окисленная форма - НАФД + превращается в восстановленную - НАДФ∗H2.
- Свободный кислород O2 - в результате фотолиза воды
- АТФ - универсальный источник энергии
- НАДФ∗H2 - форма запасания атомов водорода
Кислород удаляется из клетки как побочный продукт фотосинтеза, он совершенно не нужен растению. АТФ и НАДФ∗H2 в дальнейшем оказываются более полезны: они транспортируются в строму хлоропласта и принимают участие в светонезависимой фазе фотосинтеза.
Светонезависимая (темновая) фаза
Светонезависимая фаза происходит в строме (матриксе) хлоропласта постоянно: и днем, и ночью - вне зависимости от освещения.
При участии АТФ и НАДФ∗H2 происходит восстановление CO2 до глюкозы C6H12O6. В светонезависимой фазе происходит цикл Кальвина, в ходе которого и образуется глюкоза. Для образования одной молекулы глюкозы требуется 6 молекул CO2, 12 НАДФ∗H2 и 18 АТФ.
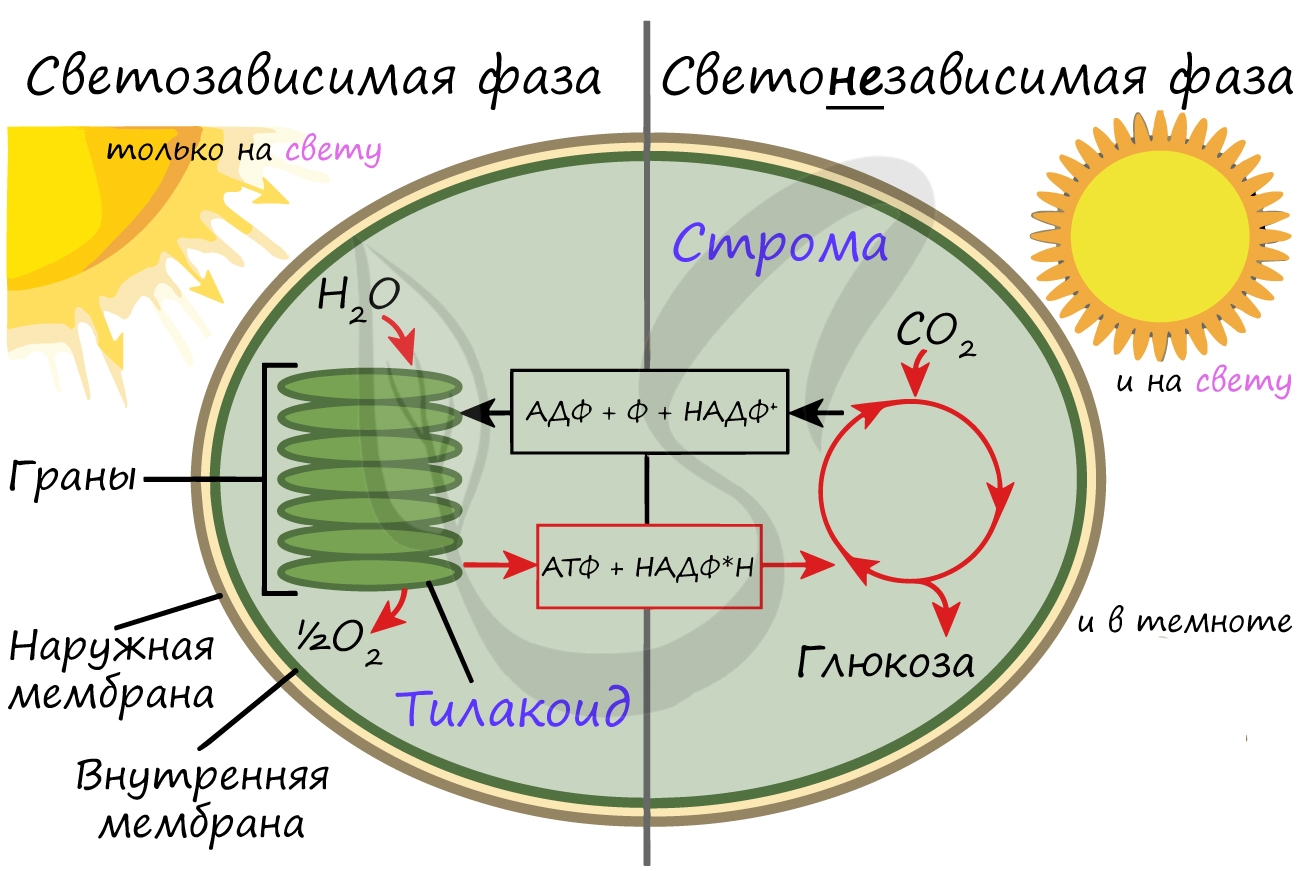
Таким образом, в результате темновой (светонезависимой) фазы фотосинтеза образуется глюкоза, которая в дальнейшем может быть преобразована в крахмал, служащий для запасания питательных веществ у растений.
Значение фотосинтеза
Значение фотосинтеза невозможно переоценить. Уверенно утверждаю: именно благодаря этому процессу жизнь на Земле приобрела такие чудесные и изумительные формы, какие мы видим вокруг себя: удивительные растения, прекрасные цветы и самые разнообразные животные.
В разделе эволюции мы уже обсуждали, что изначально в составе атмосферы Земли не было кислорода: миллиарды лет назад его начали вырабатывать первые фотосинтезирующие бактерии - сине-зеленые водоросли (цианобактерии). Постепенно кислород накапливался, и со временем на Земле стало возможно аэробное (кислородное) дыхание. Возник озоновый слой, защищающий все живое на нашей планете от губительного ультрафиолета.
- Синтезируют органические вещества, являющиеся пищей для всего живого на планете
- Преобразуют энергию света в энергию химических связей, создают органическую массу
- Растения поддерживают определенный процент содержания O2 в атмосфере, очищают ее от избытка CO2
- Способствуют образованию защитного озонового экрана, поглощающего губительное для жизни ультрафиолетовое излучение

Хемосинтез (греч. chemeia – химия + synthesis - синтез)
Хемосинтез - автотрофный тип питания, который характерен для некоторых микроорганизмов, способных создавать органические вещества из неорганических. Это осуществляется за счет энергии, получаемой при окислении других неорганических соединений (железо- , азото-, серосодержащих веществ).
Хемосинтез был открыт русским микробиологом С.Н. Виноградским в 1888 году. Большинство хемосинтезирующих бактерий относится к аэробам, для жизни им необходим кислород.
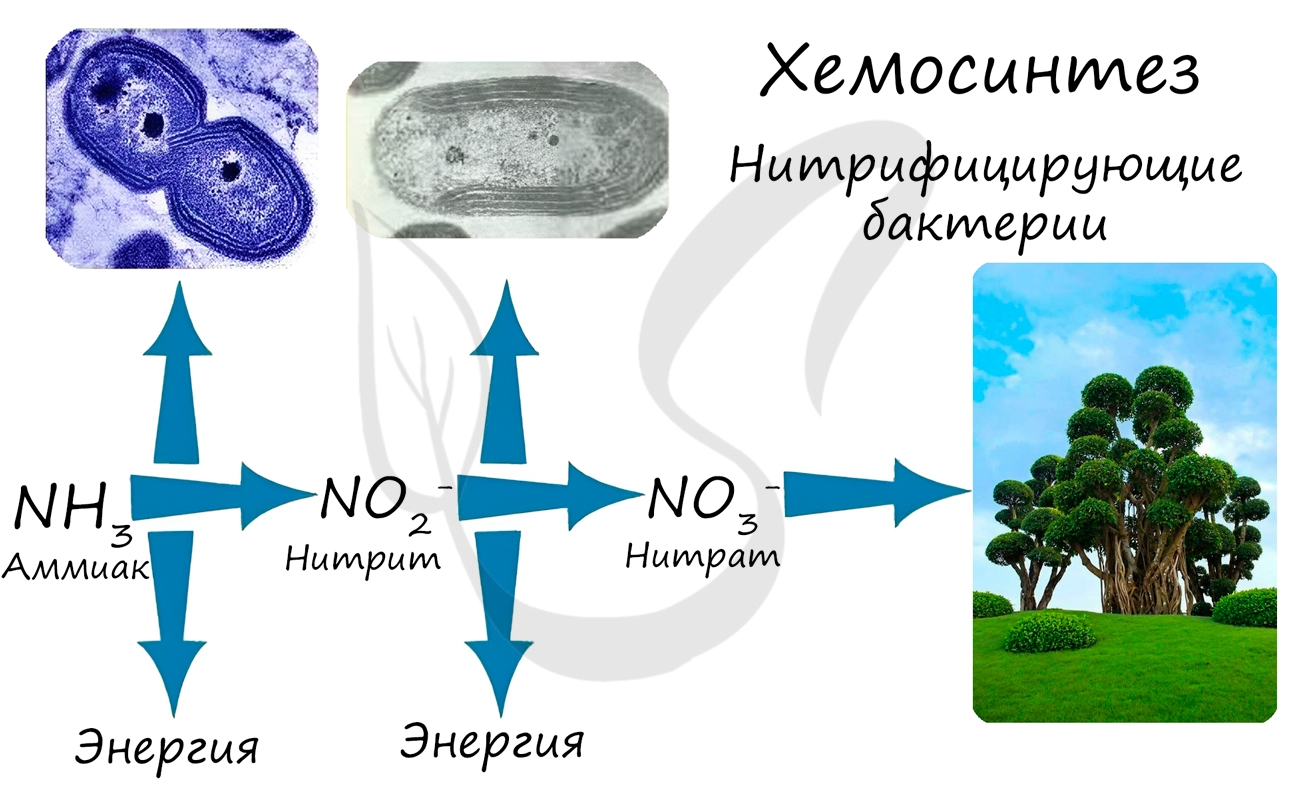
При окислении неорганических веществ выделяется энергия, которую организмы запасают в виде энергии химических связей. Так нитрифицирующие бактерии последовательно окисляют аммиак до нитрита, а затем - нитрата. Нитраты могут быть усвоены растениями и служат удобрением.
- Серобактерии - окисляют H2S --> S 0 --> (S +4 O3) 2- --> (S +6 O4) 2-
- Железобактерии - окисляют Fe +2 -->Fe +3
- Водородные бактерии - окисляют H2 --> H +1 2O
- Карбоксидобактерии - окисляют CO до CO2
Значение хемосинтеза
Хемосинтезирующие бактерии являются неотъемлемым звеном круговорота в природе таких элементов как: азот, сера, железо.
Нитрифицирующие бактерии обеспечивают переработку (нейтрализацию) ядовитого вещества - аммиака. Они также обогащают почву нитратами, которые очень важны для нормального роста и развития растений.
Усвоение нитратов происходит за счет клубеньковых бактерий на корнях бобовых растений, однако важно помнить, что клубеньковые (азотфиксирующие) бактерии, в отличие от нитрифицирующих бактерий, питаются гетеротрофно.
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Читайте также: