
Теория жакоба и моно кратко
Обновлено: 06.07.2024
Во всех клетках экспрессия генов (см. с. 234) контролируется регуляторными белками, которые связываются с определенным участком ДНК (DNA) и таким образом стимулируют пли подавляют транскрипцию гена (контроль транскрипции, см. с. 240). Действие регуляторных белков обратимо и, как правило, требует присутствия лиганда. Постоянно открывают все новые и новые регуляторные белки, в настоящее время известна, вероятно, только малая их часть. Несовершенна также их номенклатура. Как для белков, так и для участков ДНК, с которыми они связываются, используются различные наименования в зависимости от принципа действия. Регуляторный белок, который влияет на транскрипцию генов, называют фактором транскрипции. Белок, подавляющий транскрипцию, называют репрессором, а стимулирующий — индуктором. Последовательности ДНК, с которыми связываются регуляторные белки, называются регуляторными элементами. У прокариот регуляторные алименты, которые служат участками связывания РНК-полимеразы, называют промоторами, в то время как для репрессорных участков связывания употребляется название оператор. Регуляторные элементы, связывающие активирующие факторы, называют энхансерами (от англ. enhancer — усилитель), в то время как элементы, связывающие негативные (ингибирующие) факторы, — сайленсерами (от англ. silencer — успокоитель).
Многочисленные известные регуляторные белки можно разделить по механизму действия на четыре группы. Негативная генетическая регуляция, т. е. выключение соответствующих генов, может вызываться репрессорами. Некоторые репрессоры связываются с ДНК только в отсутствие специфического лиганда (1а). Комплекс репрессора с лигандом в этом случае теряет способность к связыванию и оставляет свободным участок промотора для присоединения РНК-полимеразы (1б). Часто свободный от лиганда репрессор не может связываться с ДНК, т. е. транскрипция подавляется только в присутствии лигандов (2а, 2б). Аналогично при позитивной генетической регуляции можно различать два случая. Если связывается только свободный индуктор, транскрипция подавляется соответствующими лигандами . Напротив, многие индукторы становятся активными только после образования комплекса с лигандом. К этой группе принадлежат, например, стероидные гормоны .
Согласно теории Жакоба и Моно в биосинтезе белка у бактерий участвуют по крайней мере три типа генов: структурные гены, ген-регулятор и ген-оператор. Структурные гены определяют первичную структуру синтезируемого белка. Именно эти гены в цепи ДНК являются основой для биосинтеза мРНК, которая затем поступает в рибосому и, как было указано выше, служит матрицей для биосинтеза белка.
В свою очередь деятельность оперона находится под контролирующим влиянием другого участка цепи ДНК, получившего название гена-регулятора. Поскольку структурные гены и ген-регулятор находятся в разных участках цепи ДНК, связь между ними, как предполагают Ф. Жакоб и Ж. Моно, осуществляется при помощи вещества-посредника, оказавшегося белком и названного репрессором. Образование репрессора происходит в рибосомах ядра на матрице специфической мРНК, синтезированной на гене-регуляторе. Репрессор имеет сродство к гену-оператору и обратимо соединяется с ним в комплекс. Образование такого комплекса приводит к блокированию синтеза мРНК и, следовательно, синтеза белка, т.е. функция гена-регулятора состоит, в том, чтобы через белок-репрессор прекращать деятельность структурных генов, синтезирующих мРНК. Репрессор, кроме того, обладает способностью строго специфически связываться с определенными низкомолекулярными веществами, называемыми индукторами, или эффекторами. Когда такой индуктор соединяется с репрессором, последний теряет способность связываться с геном-оператором, который таким образом выходит из-под контроля гена-регулятора, и начинается синтез мРНК.
Это типичный пример отрицательной формы контроля, когда индуктор, соединяясь с белком-репрессором, вызывает изменения его третичной структуры настолько, что репрессор теряет способность связываться с геном-оператором. Этот процесс аналогичен взаимоотношениям аллостерического центра фермента с эффектором, под влиянием которого изменяется третичная структура фермента и он теряет способность связываться со своим субстратом.
В качестве примера приведен лактозный оперон бактерии Е. coli (участок ДНК), который подвержен одновременно негативному и позитивному контролю. Оперон содержит структурные гены трех ферментов, которые необходимы для утилизации лактозы, и регуляторные элементы для управления транскрипцией оперона.
Так как лактоза превращается в клетке в глюкозу, экспрессия генов лактозного оперона не имеет смысла, когда глюкоза присутствует в клетке. Действительно гены транскрибируются только в отсутствие глюкозы и в присутствии лактозы . Регуляция достигается благодаря взаимодействию двух регуляторных белков. В отсутствие лактозы lac-penpeccop блокирует участок промотора . При наличии лактозы она превращается в изомерную аллолактозу, которая связывается с белком-репрессором и тем самым вызывает диссоциацию репрессора и оператора. Тем не менее этого недостаточно для транскрипции структурных генов. Для связывания РНК-полимеразы необходим индуктор, белок-активатор катаболитных оперонов (САР от англ. catabolite activator protein), который связывается с ДНК только в комплексе с цАМФ (cAMP). Сигнал голодания возникает только в отсутствие глюкозы.
Ф. Жакоб и Ж. Моно выдвинули в 1961 году гипотезу оперона. По этой схеме гены функционально неодинаковы. Один из них - структурный ген, содержит информацию о расположении аминокислот в молекуле белка фермента, другие выполняют регуляторные функции, оказывающие влияние на активность структурных генов – гены – регуляторы. Структурные гены располагаются рядом и образуют блок – оперон. Они программируют синтез ферментов. Кроме того в оперон входят участки, относящиеся к процессу включения транскрипции. Вся группа генов одного оперона функционирует одновременно, поэтому ферменты одной цепи реакции либо синтезируются все, либо не синтезируется ни один из них. В самом начале структуры оперона находится ген – оператор, который включает и выключает структурные гены. Оператор контролирует ген – регулятор. Ген-регулятор кодирует синтез белка-репрессора. Репрессор в активной форме блокирует транскрипцию, считывание генетической информации прекращается и весь оперон выключается. До тех пор, пока репрессор связан с геном-оператором, оперон находится в выключенном состоянии. При переходе в неактивную форму ген-оператор освобождается, происходит включение оперона и начинается синтез соответствующей РНК с последующим процессом синтеза ферментов. Оперонная система представляет собой один из механизмов регуляции синтеза белка.
2.Исходные причины экологического кризиса. Восполняемые и не восполняемые ресурсы планеты. Минимум воспроизводимого субстрата и порог воспроизводимого вещества. Сущность экологических программ (ЮНЕСКО, МАГАТЭ, ФАО, МСОП, МБП и МАВ). Основные принципы экологической этики.
У современного экологического кризиса есть несколько причин:
• безудержный и очень быстрый рост населения Земли
• несовершенные сельскохозяйственные и промышленные технологии
• легкомысленность человечества и пренебрежение законами развития биосферы
Восполняемые ресурсы: лес, вода, почва и т.д
Невосполняемые ресурсы: нефть, газ, уголь
РЕБЯТА, Минимум воспроизводимого субстрата и порог воспроизводимого вещества Я НЕ СМОГЛА НИГДЕ НАЙТИ.
ЮНЕСКО (UNESCO — United Nations Educational, Scientific and Cultural Organization) — Организация Объединённых Наций по вопросам образования, науки и культуры. Основные цели, декларируемые организацией, — содействие укреплению мира и безопасности за счёт расширения сотрудничества государств и народов в области образования, науки и культуры; обеспечение справедливости и соблюдения законности, всеобщего уважения прав и основных свобод человека, провозглашённых в Уставе Организации Объединённых Наций, для всех народов, без различия расы, пола, языка или религии. Организация была создана 16 ноября 1945 года и её штаб-квартира располагается в Париже, во Франции.
МАГАТЭ (англ. IAEA, сокр. International Atomic Energy Agency) — международная организация для развития сотрудничества в области мирного использования атомной энергии. Основана в 1957 году. Штаб-квартира расположена в Вене (Международный Венский Центр).
ФАО - продовольственная и сельскохозяйственная организация ООН (ФАО) (англ. Food and Agriculture Organization, FAO) — международная организация под патронатом ООН.
Деятельность ФАО направлена на уменьшение остроты проблемы нищеты и голода в мире путем содействия развитию сельского хозяйства, улучшению питания и решения проблемы продовольственной безопасности — доступности всем и всегда питания, необходимого для активной и здоровой жизни.
МБП - МЕЖДУНАРОДНАЯ БИОЛОГИЧЕСКАЯ ПРОГРАММА (МБП), комплекс исследований биопродуктивности экосистем, проблем охраны природы, адаптация человека к различным условиям жизни и др.

Основные принципы экологической этики:
Это – общие, или установочные принципы и практические принципы.
Установочные принципы: а) уважение ко всем формам жизни, б) биоразнообразие, в) поддержание устойчивости биосферы, г) экологическая справедливость, д) принцип предосторожности, е) природные ресурсы – общее достояние.
Принцип уважения ко всем формам жизни утверждает ценность жизни самой по себе, моральное значение каждого живого существа безотносительно к интересам человека
Принцип биоразнообразия утверждает ценность биоразнообразия и необходимость его сохранения как одного из проявлений богатства природы. Признание ценности биоразнообразия не должно иметь те же практические последствия, что и принцип уважения к любой форме возможным неизбежным потерям.
Принцип поддержания устойчивости биосферы. Сохранение биосферы, возможно, более важно, чем сохранение любой индивидуальной жизни, вида или экосистемы, за исключением, возможно, людей.
Принцип экологической справедливости утверждает равное распределение между людьми права на экологическую безопасность, при этом каждому вменяется ответственность за ее сохранение
Согласно принципу предосторожности при разработке политики, в особенности политики, имеющей гуманитарные и экологические последствия, прямые или косвенные, следует в первую очередь принимать во внимание наиболее опасный из возможных вариантов развития событий. Когда активность человека может вести к морально неприемлемому ущербу, пусть даже его возможность научно не доказана, необходимы целенаправленные усилия по их уменьшению или предотвращению. Под морально неприемлемым ущербом понимается ущерб людям или окружающей среде, когда имеется угроза жизни или здоровью, когда возможны последствия, необратимые для окружающей среды (а деятельность человека все более и более имеет необратимые последствия для окружающей среды, причем в глобальном масштабе) или ущемляющие права будущих поколений, когда предпринимаемые действия нарушают фундаментальные права тех, кто оказывается жертвой их негативных последствий и т.д.
Принцип общего достояния природных ресурсов выражает представление о планете Земля как целостности. Люди несут равную ответственность за природные ресурсы и окружающую среду, поскольку истощение природных ресурсов будет иметь свои последствия для всех людей, как ныне живущих, так и будущих поколений.
Принцип ответственности перед будущими поколениями может также вытекать из принципа экологической справедливости.
Принцип разделенной ответственности утверждает, что поскольку природные ресурсы – всеобщее достояние, ответственность за охрану окружающей среды должны разделить между собой все, она не должна быть делегирована какой либо отдельной организации или группе.
Принцип презумпции опасности вытекает из принципа уважения к жизни и принципа предосторожности и выражается в практическом требовании: те, кто предпринимает действия, имеющие последствия для окружающей среды, должны нести бремя доказательства их безопасности.
Принцип сокращения и конвергенции вытекает из принципов экологической справедливости и всеобщего достояния природных ресурсов и касается выбрасываемых в атмосферу газов, накопление которых в атмосфере усиливает парниковый эффект. Согласно этому принципу, необходимо введение ограничений на объемы выбрасываемых газов.
Ф. Жакоб и Ж. Моно выдвинули в 1961 году гипотезу оперона. По этой схеме гены функционально неодинаковы. Один из них - структурный ген, содержит информацию о расположении аминокислот в молекуле белка фермента, другие выполняют регуляторные функции, оказывающие влияние на активность структурных генов – гены – регуляторы. Структурные гены располагаются рядом и образуют блок – оперон. Они программируют синтез ферментов. Кроме того в оперон входят участки, относящиеся к процессу включения транскрипции. Вся группа генов одного оперона функционирует одновременно, поэтому ферменты одной цепи реакции либо синтезируются все, либо не синтезируется ни один из них. В самом начале структуры оперона находится ген – оператор, который включает и выключает структурные гены. Оператор контролирует ген – регулятор. Ген-регулятор кодирует синтез белка-репрессора. Репрессор в активной форме блокирует транскрипцию, считывание генетической информации прекращается и весь оперон выключается. До тех пор, пока репрессор связан с геном-оператором, оперон находится в выключенном состоянии. При переходе в неактивную форму ген-оператор освобождается, происходит включение оперона и начинается синтез соответствующей РНК с последующим процессом синтеза ферментов. Оперонная система представляет собой один из механизмов регуляции синтеза белка.
2.Исходные причины экологического кризиса. Восполняемые и не восполняемые ресурсы планеты. Минимум воспроизводимого субстрата и порог воспроизводимого вещества. Сущность экологических программ (ЮНЕСКО, МАГАТЭ, ФАО, МСОП, МБП и МАВ). Основные принципы экологической этики.
У современного экологического кризиса есть несколько причин:
• безудержный и очень быстрый рост населения Земли
• несовершенные сельскохозяйственные и промышленные технологии
• легкомысленность человечества и пренебрежение законами развития биосферы
Восполняемые ресурсы: лес, вода, почва и т.д
Невосполняемые ресурсы: нефть, газ, уголь
РЕБЯТА, Минимум воспроизводимого субстрата и порог воспроизводимого вещества Я НЕ СМОГЛА НИГДЕ НАЙТИ.
ЮНЕСКО (UNESCO — United Nations Educational, Scientific and Cultural Organization) — Организация Объединённых Наций по вопросам образования, науки и культуры. Основные цели, декларируемые организацией, — содействие укреплению мира и безопасности за счёт расширения сотрудничества государств и народов в области образования, науки и культуры; обеспечение справедливости и соблюдения законности, всеобщего уважения прав и основных свобод человека, провозглашённых в Уставе Организации Объединённых Наций, для всех народов, без различия расы, пола, языка или религии. Организация была создана 16 ноября 1945 года и её штаб-квартира располагается в Париже, во Франции.
МАГАТЭ (англ. IAEA, сокр. International Atomic Energy Agency) — международная организация для развития сотрудничества в области мирного использования атомной энергии. Основана в 1957 году. Штаб-квартира расположена в Вене (Международный Венский Центр).
ФАО - продовольственная и сельскохозяйственная организация ООН (ФАО) (англ. Food and Agriculture Organization, FAO) — международная организация под патронатом ООН.
Деятельность ФАО направлена на уменьшение остроты проблемы нищеты и голода в мире путем содействия развитию сельского хозяйства, улучшению питания и решения проблемы продовольственной безопасности — доступности всем и всегда питания, необходимого для активной и здоровой жизни.
МБП - МЕЖДУНАРОДНАЯ БИОЛОГИЧЕСКАЯ ПРОГРАММА (МБП), комплекс исследований биопродуктивности экосистем, проблем охраны природы, адаптация человека к различным условиям жизни и др.
Основные принципы экологической этики:
Это – общие, или установочные принципы и практические принципы.
Установочные принципы: а) уважение ко всем формам жизни, б) биоразнообразие, в) поддержание устойчивости биосферы, г) экологическая справедливость, д) принцип предосторожности, е) природные ресурсы – общее достояние.
Принцип уважения ко всем формам жизни утверждает ценность жизни самой по себе, моральное значение каждого живого существа безотносительно к интересам человека
Принцип биоразнообразия утверждает ценность биоразнообразия и необходимость его сохранения как одного из проявлений богатства природы. Признание ценности биоразнообразия не должно иметь те же практические последствия, что и принцип уважения к любой форме возможным неизбежным потерям.
Принцип поддержания устойчивости биосферы. Сохранение биосферы, возможно, более важно, чем сохранение любой индивидуальной жизни, вида или экосистемы, за исключением, возможно, людей.
Принцип экологической справедливости утверждает равное распределение между людьми права на экологическую безопасность, при этом каждому вменяется ответственность за ее сохранение
Согласно принципу предосторожности при разработке политики, в особенности политики, имеющей гуманитарные и экологические последствия, прямые или косвенные, следует в первую очередь принимать во внимание наиболее опасный из возможных вариантов развития событий. Когда активность человека может вести к морально неприемлемому ущербу, пусть даже его возможность научно не доказана, необходимы целенаправленные усилия по их уменьшению или предотвращению. Под морально неприемлемым ущербом понимается ущерб людям или окружающей среде, когда имеется угроза жизни или здоровью, когда возможны последствия, необратимые для окружающей среды (а деятельность человека все более и более имеет необратимые последствия для окружающей среды, причем в глобальном масштабе) или ущемляющие права будущих поколений, когда предпринимаемые действия нарушают фундаментальные права тех, кто оказывается жертвой их негативных последствий и т.д.
Принцип общего достояния природных ресурсов выражает представление о планете Земля как целостности. Люди несут равную ответственность за природные ресурсы и окружающую среду, поскольку истощение природных ресурсов будет иметь свои последствия для всех людей, как ныне живущих, так и будущих поколений.
Принцип ответственности перед будущими поколениями может также вытекать из принципа экологической справедливости.
Принцип разделенной ответственности утверждает, что поскольку природные ресурсы – всеобщее достояние, ответственность за охрану окружающей среды должны разделить между собой все, она не должна быть делегирована какой либо отдельной организации или группе.
Принцип презумпции опасности вытекает из принципа уважения к жизни и принципа предосторожности и выражается в практическом требовании: те, кто предпринимает действия, имеющие последствия для окружающей среды, должны нести бремя доказательства их безопасности.
Принцип сокращения и конвергенции вытекает из принципов экологической справедливости и всеобщего достояния природных ресурсов и касается выбрасываемых в атмосферу газов, накопление которых в атмосфере усиливает парниковый эффект. Согласно этому принципу, необходимо введение ограничений на объемы выбрасываемых газов.
Основным условием существования любых живых организмов является наличие тонкой, гибкой, согласованно действующей системы регуляции, в которой все элементы тесно связаны друг с другом. В белковом синтезе не только количественный и качественный состав белков, но и время синтеза имеют большое значение. От этого зависит приспособление микроорганизмов к условиям окружающей питательной среды как биологической необходимости или приспособление сложного многоклеточного организма к физиологическим потребностям при изменении внутренних и внешних условий.
Клетки живых организмов обладают способностью синтезировать огромное количество разнообразных белков. Однако они никогда не синтезируют все белки. Количество и разнообразие белков, в частности ферментов, определяются степенью их участия в метаболизме. Более того, интенсивность обмена регулируется скоростью синтеза белка и параллельно контролируется аллостерическим путем (см. главу 4). Таким образом, синтез белка регулируется внешними и внутренними факторами и условиями, которые диктуют клетке синтез такого количества белка и такого набора белков, которые необходимы для выполнения физиологических функций. Все это свидетельствует о весьма сложном, тонком и целесообразном механизме регуляции синтеза белка в клетке.
Согласно теории Ф. Жакоба и Ж. Моно, в биосинтезе белка у бактерий участвуют по крайней мере 3 типа генов: структурные гены, ген-регулятор и ген-оператор. Структурные гены определяют первичную структуру синтезируемого белка. Именно эти гены в цепи ДНК являются основой для биосинтеза мРНК, которая затем поступает в рибосому и, как было указано, служит матрицей для биосинтеза белка. Регуляция синтеза белка путем индукции представлена на рис. 14.12.
В свою очередь деятельность оперона находится под контролирующим влиянием другого участка цепи ДНК, получившего название гена-регулятора. Структурные гены и ген-регулятор расположены в разных участках цепи ДНК, поэтому связь между ними, как предполагают Ф. Жакоб и Ж. Моно, осуществляется при помощи вещества-посредника, оказавшегося белком и названного репрессором. Образование репрес-сора происходит в рибосомах ядра на матрице специфической мРНК, синтезированной на гене-регуляторе (рис. 14.13). Репрессор имеет сродство к гену-оператору и обратимо соединяется с ним в комплекс. Образование такого комплекса приводит к блокированию синтеза мРНК и, следовательно, синтеза белка, т.е. функция гена-регулятора состоит в том, чтобы через белок-репрессор прекращать (запрещать) деятельность структурных генов, синтезирующих мРНК. Репрессор, кроме того, обладает способностью строго специфически связываться с определенными низкомолекулярными веществами, называемыми индукторами, или эффекторами. Если такой индуктор соединяется с репрессором, то последний теряет способность связываться с геном-оператором, который, таким образом, выходит из-под контроля гена-регулятора, и начинается синтез мРНК. Это типичный пример отрицательной формы контроля, когда индуктор, соединяясь с белком-репрессором, вызывает изменения его третичной структуры настолько, что репрессор теряет способность связываться с геном-оператором. Процесс этот аналогичен взаимоотношениям алло-стерического центра фермента с эффектором, под влиянием которого изменяется третичная структура фермента и он теряет способность связываться со своим субстратом.
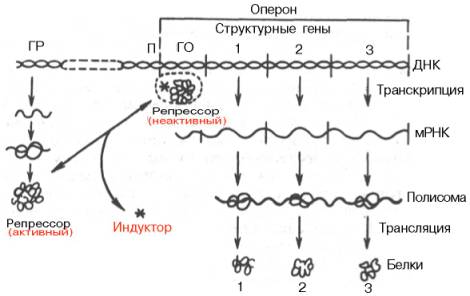
Рис. 14.12. Регуляция синтеза белка путем индукции (схема). ГР - ген-регулятор; П - промотор; ГО - ген-оператор.
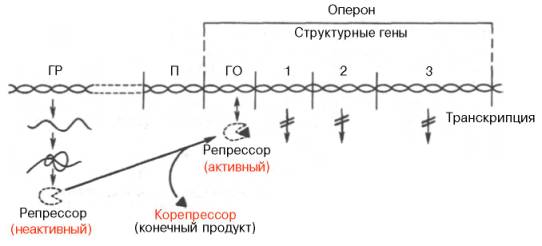
Рис. 14.13. Регуляция синтеза белка путем репрессии (схема). Обозначения те же, что на рис. 14.12.
Таким образом, биосинтез мРНК, контролирующий синтез белка в рибосомах, зависит от функционального состояния репрессора. Этот реп-рессор представляет собой тетрамерный белок с общей мол. массой около 150000. Если он находится в активном состоянии, т.е. не связан с индуктором, то блокирует ген-оператор и синтеза мРНК не происходит. При поступлении метаболита – индуктора – в клетку его молекулы связывают репрессор, превращая его в неактивную форму (или, возможно, снижают его сродство к гену-оператору). Структурные гены выходят из-под запрещающего контроля и начинают синтезировать нужную мРНК.
Конечный продукт выступает, таким образом, в качестве корепрес-сора. Имеются данные, что в качестве корепрессоров в синтезе ферментов обмена аминокислот, по-видимому, выступает не только свободная аминокислота как конечный продукт биосинтетической реакции, но и комплекс ее с тРНК – аминоацил-тРНК.
В регуляции экспрессии структурных генов специфическое участие принимает особый белок – катаболитный генактивирующий белок (от англ. catabolite gene activation protein, сокращенно CAP). Этот белок, взаимодействующий с цАМФ, образует комплекс, способствующий прикреплению РНК-полимеразы к промоторному участку генома. В присутствии комплекса САР-цАМФ фермент может начать транскрипцию оперона, включая структурные гены, т.е. в клетках имеется еще один, дополнительный САР-цАМФ-регулятор, действующий, скорее всего, в качестве положительного регулятора, поскольку его присутствие необходимо для начала экспрессии гена.
Таким образом, концепция Ф. Жакоба и Ж. Моно о механизме проявления (экспрессии) активности генов признана одним из блестящих достижений молекулярной биологии. Она явилась логическим развитием многочисленных исследований, проведенных генетиками и биохимиками в предшествующие десятилетия.
Регуляция экспрессии активности генов у эукариот осуществляется значительно более сложным путем, поскольку процессы транскрипции и трансляции разделены не только пространственно ядерной биомембраной, но и во времени. Эта регуляция базируется как минимум на 6 уровнях сложных биологических процессов, определяющих скорость синтеза и распада генетического продукта (рис. 14.14).
Для большинства эукариотических клеток, как и клеток прокариот, стадия инициации транскрипции является основной, главной регуляторной точкой экспрессии активности генов. Тем не менее имеются существенные различия: во-первых, место процессов транскрипции (в ядре) и трансляции (в цитоплазме); во-вторых, активирование транскрипции у эукари-от связано с множеством сложных изменений структуры хроматина в транскрибируемой области; в-третьих, в эукариотических клетках превалируют положительные регуляторные механизмы над отрицательными.
Положительная или отрицательная регуляция определяется типом белков, вовлеченных в механизм регуляции. Получены доказательства существования минимум 3 типов белков, участвующих в регуляции процесса инициации транскрипции, опосредованного через РНК-полимеразу: специфические факторы, репрессоры и активаторы. Первые вызывают изменение специфичности РНК-полимеразы к данному промотору или группе промоторов; репрессоры связываются с промотором, блокируя тем самым доступ РНК-полимеразы к промотору; активаторы, напротив, связываются вблизи промоторного участка, повышая связывание промотора и РНК-полимеразы.
В многоклеточных организмах среднее число регуляторных сайтов для одного гена минимум равно пяти; положительные регуляторные белки связываются со своими специфическими последовательностями в структуре ДНК (вероятнее всего, посредством водородных связей между амидной группой Глн или Асн и пуриновыми и пиримидиновыми основаниями нуклеотидов). Следует указать еще на один момент, почему эукариоти-ческая клетка использует положительные механизмы регуляции экспрессии генов. Подсчитано, что в геноме человека содержится около 100000 генов, соответственно каждая клетка при отрицательном механизме регуляции могла бы синтезировать 100000 разных репрессоров, причем в достаточных количествах. При положительном механизме регуляции большинство генов в принципе неактивно, соответственно молекула РНК-полимеразы не связывается с промотором и клетка синтезирует ограниченный и избирательный круг активаторных белков, необходимых для инициации транскрипции.
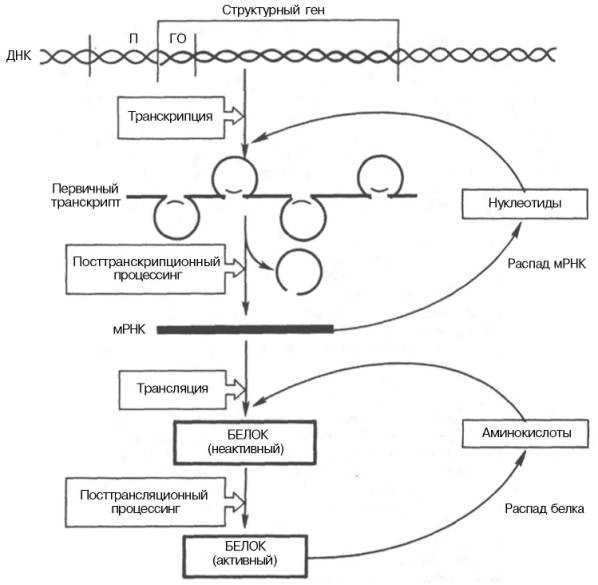
Рис. 14.14. Схематическое изображение регуляции экспрессии активности гена у эукариот.
У эукариот выделены и охарактеризованы также пять регуляторных белков, получивших название транскрипционных факторов (TF: IIА, IIВ, IID, IIЕ и IIF). Они необходимы для узнавания участка (сайта) ДНК, названного TATA (concensus последовательности, ТАТАААА). Детальный молекулярный механизм действия факторов транскрипции пока не раскрыт.
Более подробно в структурном и функциональном отношении у эука-риот изучена группа белков, получивших название белков – активаторов транскрипции. Эти белки имеют специфические структурные домены для связывания с другими, но определенными регуляторными нуклеотидными последовательностями в молекуле ДНК. В частности, они содержат домен, специфически связывающийся с ДНК, и один или несколько доменов, необходимых для активирования или взаимодействия с другими регуляторными белками. Среди этих белков – активаторов транскрипции имеются белки, содержащие богатые глутамином домены (до 25%) и богатые пролином домены. Следует отметить, однако, что некоторые из них или почти все регуляторные белки активируют транскрипцию не прямо, а опосредованно – через промежуточные белки, названные коактиваторами. Происхождение и механизм действия последних также не выяснены.
Современные знания о механизмах регуляции экспрессии генов на посттранскрипционном и посттрансляционном уровнях (см. рис. 14.4) были подробно рассмотрены ранее (см. главы 13 и 14).
Рассмотрим кратко вопрос о регуляции процессов дифференцировки клеток высших организмов. ДНК, присутствующая во всех соматических клетках, вероятнее всего, имеет одинаковую первичную структуру у данного организма и соответственно располагает информацией для синтеза любых или всех белков тела. Тем не менее клетки печени, например, синтезируют сывороточные белки, а клетки молочной железы – белки молока. Нет сомнения в том, что в дифференцированных клетках имеется весьма тонкий механизм контроля деятельности ДНК в разных тканях, обеспечивающий синтез многообразия белков.
Механизмы, лежащие в основе этой регуляции, пока неизвестны. Для их объяснения существует ряд гипотез. Предполагают, что контроль осуществляется на уровне транскрипции по аналогии с индукцией ферментов у бактерий и что в этом случае в клетках животных должны функционировать аналогичные репрессоры. С молекулой ДНК у эукариот связаны гистоны, поэтому считается, что именно эти белки выполняют роль репрессоров. Прямых доказательств их роли в качестве репрессоров не получено, хотя, как было показано, в клетках эукариот открыт класс регуляторных белков процесса транскрипции. Высказано предположение, что в ядре синтезируется высокомолекулярная молекула мРНК, содержащая информацию для синтеза широкого разнообразия белков, но в цитоплазму попадает только небольшая часть зрелой мРНК, а основная часть ее распадается. Неясны, однако, биологический смысл и назначение этого механизма избирательного распада и соответственно траты огромной массы молекулы мРНК.
Существует еще одно предположение, что на ДНК клетки синтезируются все мыслимые, возможные мРНК, которые поступают в цитоплазму, и процесс трансляции регулируется путем специфического и избирательного взаимодействия рибосом с определенными молекулами мРНК.
Исследования на клетках Е. coli позволили установить, что у бактерий существуют ферменты 3 типов:
конститутивные, присутствующие в клетках в постоянных количествах независимо от метаболического состояния организма (например, ферменты гликолиза);
индуцируемые, их концентрация в обычных условиях мала, но может возрастать в 1000 раз и более, если, например, в среду культивирования клеток добавить субстрат такого фермента;
1. Теория оперона
На основании генетических исследований индукции β-галактозидазы, участвующей в клетках Е. coli, в гидролитическом расщеплении лактозы (рис. 4-46), Франсуа Жакоб и Жак Моно в 1961 г. сформулировали гипотезу оперона, которая объясняла механизм контроля синтеза белков у прокариотов.

Рис. 4-46. Гидролиз лактозы β-галактозидазой.
В экспериментах гипотеза оперона получила полное подтверждение, а предложенный в ней тип регуляции стали называть контролем синтеза белка на уровне транскрипции, так как в этом случае изменение скорости синтеза белков осуществляется за счёт изменения скорости транскрипции генов, т.е. на стадии образования мРНК.
Большинство механизмов регуляции синтеза белков направлено на изменение скорости связывания РНК-полимеразы с промотором, влияя таким образом на этап инициации транскрипции. Гены, осуществляющие синтез регуляторных белков, могут быть удалены от оперона, транскрипцию которого они контролируют.
2. Индукция синтеза белков. Lac-оперон
Теория оперона была предложена на основании данных, полученных при изучении свойств лактозного оперона (lac-оперона) Е. coli, т.е. оперона, в котором закодированы белки, участвующие в усвоении лактозы.
Клетки Е. coli обычно растут на среде, используя в качестве источника углерода глюкозу. Если в среде культивирования глюкозу заменить на дисахарид лактозу, то по прошествии нескольких минут клетки адаптируются к изменившимся условиям. Они начинают продуцировать 3 белка, обеспечивающих утилизацию лактозы. Один из этих белков — фермент β-галактозидаза, катализирующий гидролитическое расщепление лактозы до глюкозы и галактозы.
В присутствии глюкозы клетки Е. coli содержат менее 10 молекул этих ферментов на клетку. Перенос клеток на среду, содержащую лактозу, вызывает индукцию — увеличение количества молекул каждого из ферментов до 5000 (рис. 4-47).

Рис. 4-47. Механизм индукции лактозного оперона. А - в отсутствие индуктора (лактозы) белок-репрессор связан с оператором. РНК-полимераза не может присоединиться к промотору, транскрипция структурных генов оперона не идёт; Б - в присутствии лактозы белок-репрессор присоединяет её, изменяет свою конформацию и теряет сродство к оператору. РНК-полимераза связывается с промотором и транскрибирует структурные гены: β-галактозидазы (А), катализирующей гидролиз лактозы до глюкозы и галактозы; галактозидпермеазы (В), осуществляющей транспорт лактозы и других галактозидов в клетки; тиогалакто-зидтрансацетилазы (С) — фермента, способного переносить ацетильную группу ацетил-КоА на тиогалактозу. Функция его в процессе утилизации лактозы пока неясна
Теория оперона объясняет это явление следующим образом. В отсутствие индуктора (лактозы) белок-репрессор связан с оператором. А поскольку участки оператора и промотора перекрываются, то присоединение репрессора к оператору препятствует связыванию РНК-полимеразы с промотором, и транскрипция структурных генов оперона не идёт. Когда в среде появляется индуктор, т.е. лактоза, то он присоединяется к белку-репрессору, изменяет его конформацию и снижает сродство к оператору. РНК-полимераза связывается с промотором и транскрибирует структурные гены.
3. Репрессия синтеза белков. Триптофановый и гистидиновый опероны
Снижение концентрации фермента в бактериальной клетке может осуществляться путём репрессии синтеза ферментов. Сущность этого механизма регуляции заключается в следующем: когда клетки Е. coli растут на среде, содержащей в качестве единственного источника азота соль аммония, то им приходится синтезировать все азотсодержащие вещества. Такие клетки, в частности, должны содержать все ферменты, необходимые для синтеза 20 различных аминокислот. Однако если добавить в среду культивирования одну из аминокислот, например триптофан или гистидин, то клетка перестанет вырабатывать весь набор ферментов, необходимых для синтеза этих аминокислот из аммиака и источника углерода. Репрессия синтеза ферментов, катализирующих последовательность реакций метаболического пути конечным продуктом, как это имеет место в случае ферментов синтеза гистидина или триптофана, называется репрессией конечным продуктом.

Рис. 4-48. Механизм репрессии синтеза ферментов, участвующих в образовании гистидина. А - в отсутствие корепрессора (гистидина) белок-репрессор не имеет сродства к оператору, РНК-полимераза присоединяется к промотору, и происходит транскрипция 10 структурных генов, кодирующих строение ферментов, участвующих в синтезе гистидина; Б - в присутствии гистидина в среде комплекс бел ка-реп рессора с корепрессором, т.е. Гис, связывается с оператором, препятствует присоединению РНК-полимеразы к промотору и останавливает транскрипцию
Следует иметь в виду, что репрессия и индукция синтеза белков у прокариотов реализуют принципы адаптации к меняющимся условиям существования и клеточной экономии: ферменты появляются в клетках, когда в них существует потребность, и перестают вырабатываться, если потребность исчезает.
Читайте также: