
Сведения о пуриновых и пиримидиновых основаниях в днк кратко
Обновлено: 30.06.2024
Пуриновые и пиримидиновые основания, входящие в состав нуклеотидов, представляют собой замещенные производные пурина и пиримидина (рис. 34.1). Положения атомов в ароматическом кольце пронумерованы в соответствии с принятой номенклатурой. Обратите внимание на то, что нумерация в пуриновом и пиримидиновом кольцах ведется в противоположных направлениях, при этом атом углерода под номером 5 в обеих молекулах находится в одном и том же положении. Сопряжение n-электронных облаков обусловливает плоскую структуру пуриновых и пиримидиновых оснований. Значение этого явления обсуждается в гл. 37.
Главные основания
Главные пиримидиновые основания и у прокариот, и у эукариот — это цитозин, тимин и урацил
Рис. 34.1. Структура пурина и пиримидина. Атомы пронумерованы согласно международной системе.
Рис. 34.2. Три главных пиримидиновых основания, входящие в состав нуклеотидов.
(рис. 34.2). Из пуриновых оснований чаще всего встречаются аденин и гуанин. Два других — ксантин и гипоксантин—являются интермедиатами в процессах их метаболизма (рис. 34.3). У человека в роли конечного продукта катаболизма пуринов выступает окисленное пуриновое основание—мочевая кислота (гл. 35).
Помимо пяти названных выше главных оснований известны и менее широко представленные минорные основания. Некоторые из них присутствуют только в нуклеиновых кислотах бактерий и вирусов, но многие также найдены в составе про- и эукариотических ДНК и транспортных РНК.
Рис. 34.3. Главные пуриновые основания, входящие в состав нуклеотидов.
Рис. 34.4. Структура двух необычных природных пиримидиновых оснований.
Так, и бактериальная ДНК, и ДНК человека содержат значительные количества 5-метилцитозина; в бактериофагах обнаружен 5-гидроксиметилцитозин (рис. 34.4). Необычные основания выявлены в матричной РНК - -метиладенин, -диметиладенин и N7-мeтилгуанин (рис. 34.5). У бактерий также обнаружен модифицированный урацил с присоединенной по группой. Функции этих замещенных пуринов и пиримидинов до конца не выяснены.
В клетках растений выявлена серия пуриновых оснований с метальными заместителями (рис. 34.6). Многие из них фармакологически активны. В качестве примера можно привести кофейные зерна, содержащие кофеин , чайный лист, содержащий теофиллин , и какао-бобы, в состав которых входит теобромин . Биологические свойства этих веществ описаны в гл. 35 при обсуждении метаболизма циклических нуклеотидов.

Статья будет посвящена базовому, но крайне интересному разделу биохимии — метаболизму азотистых оснований. Сегодня мы рассмотрим то, как из аминокислот и глюкозы получается материал, кодирующий нашу биологическую сущность, — нуклеиновые кислоты (ДНК и РНК). Это понадобится нам для понимания очень важных вещей. Например, как одна мутация в метаболизме нуклеотидов способна подарить ребенку тяжелейший иммунодефицит или сделать из него глубокого инвалида. Почему не живут долго и счастливо многие из тех, кто злоупотребляет вином, красным мясом, шоколадом и прочими радостями. Почему иногда умирают (во всяком случае, часто умирали раньше) при проведении противоопухолевой химиотерапии. В статье, посвященной нарушению метаболизма азотистых оснований, ты узнаешь и о редких заболеваниях, о которых забывают 90 % врачей, принимая их за другие, более известные и очевидные. Но для начала надо постичь базу, ведь без понимания метаболизма азотистых оснований перечисленные мною факты и заболевания будет крайне сложно понять. Приступим.
Нуклеотидом мы будем называть соединение, представляющее собой совокупность трех частей [3]:
- азотистое основание (аденин, гуанин, цитозин, урацил, тимин);
- пятиуглеродный сахар рибоза (дезоксирибоза);
- остатки фосфорной кислоты (от одного до трех).
Как я уже намекнул, азотистых оснований много во вкусной, но не всегда здоровой пище: бекон, красное мясо, ветчина, фасоль, спаржа, сыр, яйца, пиво, грибы и так далее. Бьюсь об заклад, что-то в твоем рационе из этого хоть иногда, да присутствует (если ты на стипендии. Если нет – работай над этим).
Попадая в твой ЖКТ, в 12-перстную кишку, нуклеиновые кислоты, содержащиеся в пище, расщепляются рибонуклеазами, нуклеотидазами и прочими ферментами поджелудочной железы. Продукты расщепления в виде нуклеотидов и нуклеозидов (иногда и вовсе азотистых оснований) попадают в энтероциты (клетки слизистой оболочки тонкой кишки).
В энтероцитах есть ксантиноксидаза, которая переводит пурины (аденин, гуанин) в мочевую кислоту. Мочевая кислота, как правило, стремится всосаться в кровь и выйти с мочой через почки. Может также пропустить себя в просвет кишки и выйти с калом (кстати, это бывает и весьма ощутимо, если имеется декомпенсированная патология почек).

Пурины — гуанин и аденин — формируются буквально из ничего. Все, что нам нужно для создания пуринового кольца — глюкоза (поступающая в пентозо-фосфатный цикл), глицин, две молекулы глутамина, одна молекула аспартата, пара молекул активной метилированной формы фолиевой кислоты + немного терпения и приправы в виде пяти молекул АТФ [1].
Я не хочу грузить тебя серией реакций синтеза пуринов, ее на экзамене полностью никто не спросит, и, скорее всего, для принятия решений в своей работе она тебе тоже не нужна. Но кое-что ты знать должен.
Синтез пуринов проходит два основных этапа: формирование инозинмонофосфата (ИМФ) и получение из ИМФ таких нуклеотидов, как ГМФ и АМФ.
Необходимо знать ключевые моменты этого синтеза.
Первый из них — лимитирующая реакция синтеза пуринов — формирование фосфорибозилпирофосфата (ФРПФ).
(Для тех, кто не понял: лимитирующая реакция — это та реакция, скорость которой определяет скорость всего процесса, а также та, что подвергается наиболее жесткой и сложной регуляции).
Далее следует еще порядка девяти реакций, которые из 5-фосфорибозил-1-амина сделают инозинмонофосфат (ИМФ). На этом закончится первый этап синтеза. Для экзамена и последующей работы тебе нужно знать, какие молекулы участвуют в формировании пуринового кольца, и эти две реакции. Для жизни — и того меньше.
2 этап. Чрезвычайно важен и интересен. Два брата, два параллельно идущих, как ты со своим другом, ̶п̶о̶ ̶н̶а̶к̶л̶о̶н̶н̶о̶й̶ к синтезу АМФ и ГМФ.
Что интересно: конечные продукты этого чрезвычайно интересного (никому) синтеза — ГТФ и АТФ — помогают дружественному параллельному циклу, даруя энергию для синтеза нуклеотида. Несколько ниже приведена иллюстрация вышеописанных биохимических событий.

Катаболизм пуринов не менее важная и интересная серия процессов, потому что так же, как и синтез пуринов, их катаболизм является ключевым моментом для понимания патогенеза ряда заболеваний. Большинство нуклеотидов подвергаются повторному использованию (отдать некому, выкинуть жалко), заново включаясь в синтез нуклеотидов. Проследим путь их катаболизма.

Как мы видим, аденозин (аденин + рибоза) стремится стать гипоксантином. Но на пути к этому ему нужно перевоплотиться назад в инозинмонофосфат. В этом ему поможет аденозиндезаминаза. Хорошо запомни этот фермент, мы к нему не раз еще вернемся. Гипоксантин, кстати, может стать просто ксантином, чем обязан ксантиноксидазе.
Гуанозин считает стадию гипоксантина для себя недостойным (не может смириться с тем, что именно АТФ, а не ГТФ, является основным источником энергии в клетке, старые обиды несчастной молекулы). В силу этого ГТФ не церемонится и становится ксантином, минуя стадию образования гипоксантина. И здесь перед ним встает выбор из двух путей. Путь 1: гипоксантин и ксантин станут мочевой кислотой и оставят после себя лишь воспоминания, удалившись в мочу (или в суставы и почечные канальцы в виде уратов). Путь 2: камбэкнуться в ИМФ (а затем в АМФ) и ГМФ. В этом им поможет братан, которого во дворе прозвали гипоксантингуанинфосфорибозил-трансферазой (ГГФРТ).
Получившиеся вновь ГМФ и ИМФ (🡪 АМФ) превращаются в ГТФ и АТФ путем присоединения фосфорных остатков. Здесь все просто.
Резюме: для нас клиническое значение будут иметь нижеперечисленные ферменты:
- Дезаминаза (аденозиндезаминаза)
- Ксантиноксидаза
- Гипоксантингуанинфосфорибозил-трансфераза
- Фосфорибозилпирофосфат-трансфераза (она же фосфорибозилдифосфат-трансфераза)
Их значение для реализации той или иной патологии будет обсуждаться в соответствующей статье.

Взглянув на схему, мы видим, что конечным продуктом этого этапа является уридинмонофосфат (УМФ). С обостренным чувством собственной важности УМФ дает начало:
- уридинтрифосфату (УТФ) — путем реакции присоединения двух фосфорных остатков;
- цитидинтрифосфату (ЦТФ) — через образование УТФ и путем изнасилования ЦТФ-синтетазой.
Все образованные соединения — АТФ, ГТФ, ЦТФ, УТФ — используются для синтеза рибонуклеиновых кислот, функцию которых ты должен был помнить не давеча как с третьего класса средней школы. Они имеют важное значение в матричных биосинтезах: в транскрипции ДНК, синтезе белка.
Но у нас есть еще и ДНК. Та штука, которая определяет твой фенотип и все, что из него вытекает. ДНК-полимераза установила фейс-контроль для синтеза ДНК. Он заключается в том, что все субстраты должны быть дезоксирибонуклеотидом [1, 3].

Есть исключение из правил. Это касается УТФ. Чтобы попасть в ДНК, ему нужно притвориться не просто дезоксирибонуклеотидом. Ему нужно притвориться ТТФ (тимидинтрифосфат). Что он и делает:

Далее остается присоединить остатки фосфорной кислоты и получить тимидинтрифосфат [1].
Важно отметить, что тимидилатсинтаза и реакция образования ТМФ являются чрезвычайно важными в отношении синтеза ДНК. Также здесь можно заметить, что необходимым кофактором является тетрагидрофолиевая кислота.
Именно данная реакция является мишенью метотрексата — цитостатического противоопухолевого препарата, являющегося антиметаболитом — структурным аналогом фолиевой кислоты, который проявляет к тимидилатсинтазе недвусмысленное сродство. Связываясь с ней, метотрексат останавливает реакцию образования ТМФ, тем самым обрывая синтез ДНК в самом зародыше — на этапе образования ее структурных компонентов. Метотрексат нашел применения в онкологии (преимущественно при гемобластозах) и в ревматологии (ревматоидный артрит [5]).
В нашей жизни ничего не вечно. Пиримидины тоже гибнут, но распадаясь, оставляют после себя ряд предшественников, которые участвуют в метаболизме, но уже другими путями.
Комментируя сложную схему распада пиримидинов, я акцентирую внимание на двух важных вещах:
- пиримидины, в отличие от пуринов, не подвергаются реутилизации;
- продукты их распада — аланин и бутират — в серии превращений становятся пировиноградной кислотой и оксалоацетатом. Их дальнейшая судьба связана с участием в цикле трикарбоновых кислот и процессе получения энергии [3].
Но этим не исчерпывается значение нуклеотидов.
Детоксикация. Кое-кто из них используется в организме для детоксикации ксенобиотиков — лекарств, токсинов и иных химикатов, поступающих в организм, а также для детоксикации липофильных (т. е. растворимых в липидах и нерастворимых в воде) соединений: гормонов, билирубина. Суть детоксикации (обезвреживания) — перевести плохо растворимое в воде вещество в хорошо растворимое, что необходимо для последующего выведения через почки с мочой из организма. Достигается это присоединением полярной химической группы к исходному хулигану. Сначала формируется гидроксильная группа (если таковой нет, это этап образования более полярных соединений). А затем наступает этап конъюгации… Посмотрим, как это выходит на примере тетрагидрокортизола (рис. 11) [2].

А вот схема для менее дотошных, чем автор данной лекции.


Важное значение эта форма УДФ имеет и для метаболизма билирубина: благодаря УДФ-диглюкурониду токсичный липофильный билирубин становится полярным, растворимым в крови и моче, конъюгированным билирубином [1, 2].
Гликоген. Неоценимое значение УДФ имеет и для образования гликогена.

Гликоген — это сравнительно инертная макромолекула, которая является резервуаром источника энергии в печени и мышцах. Через восприятия сигнала от инсулина глюкоза посредством активации (в виде присоединения фосфорного остатка и изомеризации) прыгает на гликоген благодаря посредничеству УТФ [1].
Вторичные сигнальные молекулы. Нуклеотиды, в частности, цАМФ и цГМФ, играют важную роль в процессе реализации клеточных функций и межклеточного взаимодействия. Клетка, будучи кирпичом биологического коммунизма, воспринимает сигналы от вышестоящих регуляторных систем, которые контролируют ее деятельность.
Это может выражаться в:
- изменении экспрессии генов;
- реализации действия нейромедиаторов в твоей черепной и позвоночной коробках.
Такие молекулы, как цАМФ и цГМФ, являются одними из важных вторичных мессенджеров. Клетка, воспринимая сигнал извне (будь то гормон, нейромедиатор или любой другой разбойник), изменяет внутри себя концентрацию этих мессенджеров. Те вмешиваются в серию внутриклеточных последовательных реакций и тем самым усиливают первоначальный сигнал [4].

Здесь пример воздействия адреналина на метаболизм гликогена. Концентрация цАМФ растет, что приводит к активации протеинкиназы А. Та активирует гликогенфосфорилазу, что приводит к высвобождению из гликогена глюкозы и получению дополнительного источника энергии для выполнения различных нужд организма [4].
Примеров, когда нуклеотиды играют важную роль в организме, чрезвычайно много. Вспомнить хотя бы о том, кто в клетках является главным источником энергии (АТФ). Нуклеотиды в ходе своего метаболизма могут становиться источником иных метаболитов, которые могут принимать участие в энергетическом метаболизме (α-кетоглутарат, пировиноградная кислота 🡪 ацетил-Коа) и так до бесконечности. Придет время, и мы обсудим те случаи, когда метаболизм пуринов и пиримидинов в силу тех или иных причин нарушается [1].
Пурины и пиримидины – органические вещества, входящие в структуру нуклеиновых кислот, коферментов и макроэргических соединений. Нарушение метаболизма азотистых оснований возникает вследствие генетически обусловленных дефектов ферментов, участвующих в обмене пуринов и пиримидинов.
Синонимы русские
Наследственные дефекты метаболизма азотистых оснований.
Синонимы английские
Purine and Pyrimidine Panel, Urine.
Метод исследования
Высокоэффективная жидкостная хроматография-масс-спектрометрия (ВЭЖХ-МС).
Единицы измерения
мкмоль / ммоль креатинина (микромоль на миллимоль креатинина).
Какой биоматериал можно использовать для исследования?
Разовую порцию мочи.
Как правильно подготовиться к исследованию?
- Исключить (по согласованию с врачом) прием мочегонных препаратов в течение 48 часов до сбора мочи.
Общая информация об исследовании
Пурины и пиримидины – гетероциклические органические азотистые вещества, входящие в состав нуклеотидов и нуклеозидов. Они являются важными структурными элементами нуклеиновых кислот (ДНК, РНК), источников энергии (например, АТФ), ферментов (НАДФ, НАД, ФАД).
Дефекты ферментов, принимающих участие в разных этапах метаболизма пуринов и пиримидинов, могут приводить к развитию заболевания. Патогенез данных состояний связан с накоплением в клетках и биологических жидкостях избыточного количества азотистых оснований и их метаболитов, которые могут быть токсичными и способными повреждать генетический материал и функцию клеток. Описано около 30 нарушений различных этапов метаболизма пуринов и пиримидинов, но клинически проявляются только 17. Основные лабораторные признаки данных заболеваний – это изменение содержания пуринов и пиримидинов в биологических жидкостях. Первые симптомы могут возникать как в раннем детстве, так и в старшем возрасте, а клинические проявления могут значительно варьироваться по степени тяжести. Наиболее часто при патологии пуринового и пиримидинового обмена повреждаются нервная система (задержка развития, аутизм, эпилептические приступы), кроветворная ткань (анемия) и почки (нефропатия, мочекаменная болезнь).
Классическим примером приобретенного нарушения пуринового обмена является подагра – заболевание, сопровождающиеся повышением уровня мочевой кислоты в крови и отложением уратов в тканях. Однако необходимо учитывать, что гиперурикемия (повышение уровня мочевой кислоты в крови) у людей старше 40 лет связана не только с генетической предрасположенностью, но и с особенностями питания, употреблением алкоголя, нарушением экскреторной функции почек. Гиперурикемия и подагра в более молодом или детском возрасте в большинстве случаев ассоциированы с наследственными дефектами ферментов пуринового обмена.
Тяжелое нарушение метаболизма пуринов – синдром Леша – Нихана (абсолютный дефицит гипоксантин-гуанин-фосфорибозилтрансферазы), который наследуется совместно с Х-хромосомой и проявляется тяжелыми неврологическими нарушениями, задержкой психомоторного развития, церебральным параличом, аутоагрессивным поведением и уратной нефропатией. Симптомы заболевания чаще становятся заметными в возрасте 3-12 месяцев. При данной патологии в биологических жидкостях возрастает концентрация мочевой кислоты и гипоксантина.
К врождённым нарушениям пуринового обмена относятся дегидроксиаденинурия, наследственная ксантинурия, синдром Келли – Зигмиллера и др. Дефицит аденозин-дезаминазы и пурин-нуклеозид-фосфорилазы приводит не только к неврологическим нарушениям, но и к снижению количества лимфоцитов и иммунодефициту, который проявляется рецидивирующими тяжелыми инфекциями.
Патология пиримидинового обмена наблюдается и диагностируется значительно реже. К нарушениям метаболизма пиримидинов относятся оротовая ацидурия, дефицит пиримидин-5-нуклеотидазы, дефицит дигидропиримидин-дегидрогеназы.
Некоторые из наследственных нарушений обмена азотистых оснований при своевременном выявлении поддаются коррекции, для других применяется симптоматическая терапия и разрабатываются новые методы лечения. Диагноз верифицируется на основании комплексных клинико-генеалогических данных и результатов лабораторного исследования.
Азотистые основания — это ароматические гетероциклические соединения, производные пиримидина или пурина. Пять соединений этого класса являются основными структурными компонентами нуклеиновых кислот. Общими для всей живой материи. Пуриновые основания аденин (Ade, но не А) и гуанин (Guа), а также пиримидиновое основание цитозин (Cyt), входят в состав ДНК и РНК. В состав ДНК входит также тимин (Thy), 5-метил-производное урацила. Основание урацил (Ura) входит только в состав РНК.
| 2,4-диоксопиримидин | 5-метил-2,4-диоксопиримидин 5-метилурацил | 4-амино-2-оксопиримидин |
Проявляют свойства очень слабого основания, т.к. атомы азота в sp 2 -гибридизированном состоянии довольно прочно удерживают неподеленную электронную пару.

| 6-аминопурин | 2-амино-6-оксопурин |
роявляют амфотерные свойства. Слабые основные свойства связаны с атомами азота в пиримидиновом (шестичленном) цикле, а слабые килотные обусловлены N-H группой в пятичленном цикле.
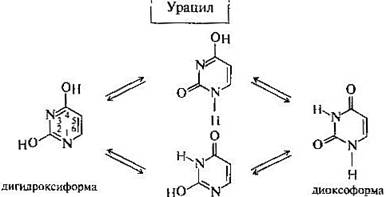
Одним из важных свойств свободных азотистых оснований (содержащих оксигруппы) является возможность их существования в двух таутомерных формах, в частности лактим- и лактамной формах, в зависимости от значения рН среды: при рН 7,0 они представлены в лактамной форме, при снижении величины рН – в лактимной форме.



Азотистые основания — это ароматические гетероциклические соединения, производные пиримидина или пурина. Пять соединений этого класса являются основными структурными компонентами нуклеиновых кислот. Общими для всей живой материи. Пуриновые основания аденин (Ade, но не А) и гуанин (Guа), а также пиримидиновое основание цитозин (Cyt), входят в состав ДНК и РНК. В состав ДНК входит также тимин (Thy), 5-метил-производное урацила. Основание урацил (Ura) входит только в состав РНК.
| 2,4-диоксопиримидин | 5-метил-2,4-диоксопиримидин 5-метилурацил | 4-амино-2-оксопиримидин |
Проявляют свойства очень слабого основания, т.к. атомы азота в sp 2 -гибридизированном состоянии довольно прочно удерживают неподеленную электронную пару.
| 6-аминопурин | 2-амино-6-оксопурин |
роявляют амфотерные свойства. Слабые основные свойства связаны с атомами азота в пиримидиновом (шестичленном) цикле, а слабые килотные обусловлены N-H группой в пятичленном цикле.
Одним из важных свойств свободных азотистых оснований (содержащих оксигруппы) является возможность их существования в двух таутомерных формах, в частности лактим- и лактамной формах, в зависимости от значения рН среды: при рН 7,0 они представлены в лактамной форме, при снижении величины рН – в лактимной форме.
Читайте также: