
Строение клеточной стенки грамотрицательных бактерий кратко
Обновлено: 04.07.2024
Клеточная стенка грамотрицательных бактерий значительно тоньше, и у большинства из них ее толщина составляет 14 – 18 нм. Четко выделяются два слоя – пластичный и ригидный, они связаны лабильно и отделяются друг от друга при обработке додецилсульфатом натрия. Основная особенность клеточной стенки грамотрицательных бактерий: ригидный слой тонкий, представлен одним или, редко, двумя слоями пептидогликана, на долю которого приходится до 5 – 10 % сухого веса стенки. Для пептидогликана характерно низкое содержание поперечных сшивок между пептидными цепочками, однако в нем почти всегда имеется диаминопимелиновая кислота.
В составе клеточной стенки содержится много липопротеинов, фосфолипидов, липополисахарид, больше белка и, как правило, отсутствуют тейхоевые кислоты. Пластичный слой клеточной стенки у грамотрицательных бактерий представляет собой сложную мозаику, образованную из липопротеинов, липополисахаридов и наружной мембраны.
Липопротеины связывают наружную мембрану с пептидогликаном (белок связан с диаминопимелиновой кислотой бокового тетрапептида, а липид – нековалентно с наружной мембраной).
Липополисахарид (ЛПС) состоит из комплекса липида А и связанного с ним полисахарида, состоящего из ядра, которое одинаково у всех грамотрицательных бактерий, и терминальной цепочки из повторяющихся сахаров (рис. 6). Последние у разных видов бактерий различаются по химической природе. Они обычно представлены линейными трисахаридами или разветвляющимися тетра– или пентасахаридами. Терминальные повторяющиеся единицы полисахарида ЛПС располагаются на поверхности клетки в виде микроворсинок и определяют ее антигенную специфичность. ЛПС синтезируется на цитоплазматической мембране, а затем транспортируется в наружную часть клетки, он прикреплен к наружной мембране с помощью гидрофобных связей. ЛПС выполняет две важнейшие функции у грамотрицательных бактерий: во-первых, он определяет их антигенную специфичность, а во-вторых, является одним из главных факторов их патогенности. ЛПС – это эндотоксин. Его токсичность определяется липидом А. Кроме того, ЛПС в организме запускает синтез около 20 различных биологически активных соединений, которые опосредуют патогенез эндотоксикоза, и обладает пирогенным действием.
Наружная мембрана, подобно любой биологической мембране, состоит из двух слоев липидов, но в ней значительная часть фосфолипидов наружного слоя замещена молекулами липополисахаридов и набором белков, локализованных мозаично (рис. 7). В состав этих белков, заключенных в фосфолипидную матрицу, входят 3 или 4 основных (major), которые составляют около 70 % суммарных белков наружной мембраны; липопротеины и второстепенные белки, числом более 10. Два из основных белков проходят через оба слоя мембраны и прочно связаны с пептидогликаном. Эти белки-порины располагаются в виде триплетов и образуют диффузионные поры, через которые в клетку проникают мелкие гидрофильные молекулы. Второстепенные белки выполняют разнообразные специфические функции: одни из них участвуют в облегченной диффузии, другие – в активном транспорте молекул через наружную мембрану и выступают в качестве специфических рецепторов для фагов и колицинов. Некоторые из этих белков участвуют в конъюгации (являются рецепторами для донорных ворсинок), в контроле репликации ДНК и регуляции клеточного деления. Наружная мембрана осуществляет также функцию барьера, через который в клетку не способны проникать крупные молекулы (один из механизмов неспецифической устойчивости грамотрицательных бактерий к антибиотикам). Если бактерии поместить в гипертонический раствор, наступает резкое обезвоживание клеток, цитоплазма съеживается, и протопласт отходит от клеточной стенки. Это явление называется плазмолизом. В результате плазмолиза клетки гибнут. Этим свойством широко пользуются для консервирования пищевых продуктов с помощью концентрированных растворов поваренной соли или сахара. Однако плазмолиз проявляется не в одинаковой степени у разных видов бактерий. К нему особенно устойчивы Bacillus subtilis, Staphylococcus aureus, которые являются частыми виновниками пищевых отравлений. В случае помещения бактерий в дистиллированную воду или гипотонические растворы солей происходит противоположное явление – плазмоптиз: вода устремляется в клетки, происходит их набухание и разрушение.
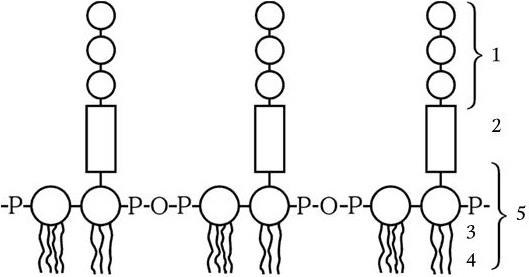
Рис. 6
1 – повторяющиеся единицы; 2 – ядро; 3 – полимер дисахарид-фосфата; 4 – жирные кислоты; 5 – липид
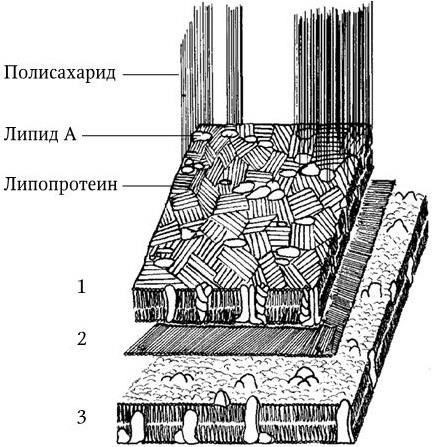
Рис. 7
. Схематическое изображение структур наружной мембраны (1), пептидогликана (2) и плазматической мембраны (3) E. coli
При обработке грамположительных бактерий ферментами, разрушающими пептидогликан, возникают протопласты, т. е. структуры, полностью лишенные клеточной стенки. Обработка грамотрицательных бактерий лизоцимом разрушает только слой пептидогликана клеточной стенки, но наружная мембрана (или, по крайней мере, часть ее) сохраняется. Такие структуры получили название сферопластов. Протопласты и сферопласты имеют сферическую форму и в соответствующих осмотических условиях сохраняют жизнеспособность. Особенно чувствительны к изменению осмотического давления протопласты. При определенных условиях они способны к размножению подобно L-формам бактерий. Нарушение синтеза клеточной стенки лежит в основе L-трансформации бактерий.

Грамотрицательные бактерии – это бактерии не красящиеся по Граму. По структуре и химическому составу клеточной стенки грамположительные бактерии резко отличаются от грамотрицательных бактерий [3] [1] .

Клеточная стенка грамотрицательных бактерий

1. Наружная мембрана; 2. Периплазматическое пространство; 3. Муреин; 4. Периплазматическое пространство; 5. Цитоплазматическая мембрана [2] .
Особенности структуры и химического состава клеточной стенки
Клеточная стенка грамотрицательных бактерий многослойна, толщиной 14–17 нм. Внутренний слой клеточной стенки представлен муреином. На его долю приходится от 1 до 10% ее сухой массы [2] .
Структурные микрофибриллы пептидогликана грамотрицательных бактерий сшиты менее компактно. Поры в их пептидогликановом слое значительно шире, чем в молекулярном каркасе грамположительных бактерий [2] .
Внешний слой клеточной стенки грамотрицательных бактерий (наружная или внешняя мембрана) образован фосфолипидами, липопротеинами и белками [2] .
По строению наружная мембрана характеризуется типичной организацией, характерной для элементарных мембран. Основная фракция наружной мембраны – липиды. Они составляют в среднем 22% сухой массы клеточной стенки [2] .
В наружной мембране находятся трансмембранные белки, которые пронизывают мембрану насквозь. Они представляют собой заполненные водой каналы и гидрофильные поры в липофильной мембране и названы поринами. Существует несколько типов поринов, осуществляющих транспорт через мембрану гидрофильных низкомолекулярных веществ [2] .
Одна из отличительных особенностей грамотрицательных бактерий – отсутствие в клеточной стенке тейхоевых кислот [2] .
Компоненты клеточной стенки грамотрицательных бактерий разделены электронно-прозрачным слоем и четко отделены от цитоплазматической мембраны. Пространство между цитоплазматической и наружной мембраной получило наименование – периплазматическое пространство [2] .
В периплазматическом пространстве расположены белки, такие как протеиназы, нуклеазы, периферические белки цитоплазматической мембраны, рестриктазы, пермеазы (связующие белки, участвующие в переносе некоторых субстратов в цитоплазму) [2] .
Грамотрицательных фитопатогенные бактерий
В девятом издании определителя бактерий Берджи грамотрицательные эубактери, имеющие клеточные стенки, относятся к типу Gracilicutes. К их числу принадлежат многие фитопатогенные бактерии:

Фотография сделанная трансмиссионым электронным микроскопом. Клеточные покровы цианобактерии Phormidium uncinatum. Клеточные покровы (CW) состоит из, цитоплазматической мембраны (CM), пептидогликанового слоя (P), периплазматического пространства (PS) и внешней мембраны (OM). В добавок, у цианобактерий есть два дополнительных внешних слоя, типичных для подвижных клеток, внешний зубчатый слой (EL) и слой осцилирующих волоско-подобных фибрилл (F). (CJ) Место секреции слизи; (JP) поры во внешней мембране. [1]


Грамотрицательные бактерии это бактерии, которые не окрашиваются кристаллическим фиолетовым при окрашивании по Граму . [2] В отличие от грамотрицательных, которые полностью обесцвечиваются, грамположительные бактерии сохранят фиолетовую окраску даже после промывания обесцвечивающим растворителем (спирт). После промывания растворителем при окрашивании по Граму добавляется контрастный краситель (обычно сафранин) , который окрашивает всех грамотрицательных бактерий в красный или розовый цвет. Это происходит из-за наличия внешней мембраны, препятствующей проникновению красителя внутрь клетки. Сам по себе тест полезен при классификации бактерий и разделении их на две группы относительно строения их клеточной стенки. Из-за своей более мощной и непроницаемой клеточной стенки грамотрицательные бактерии более устойчивы к антителам чем грамположительные.
Обычно патогенность грамотрицательных бактерий связывают с определёнными компонентами их клеточных стенок, а именно, с липополисахаридным слоем (ЛПС или эндотоксический слой). [2] В человеческом организме ЛПС вызывает иммунный ответ, который характеризуется синтезем цитокинов и активацией иммунной системы. Обычной реакцией на синтез цитокинов является воспаление, что также может привести к увеличению количества токсичных веществ в организме хозяина.
Содержание
Характеристика
Общие признаки, свойственные большинству грамотрицательных бактерий:
- Наличие двух мембран, между которыми находится клеточная стенка и периплазматическое пространство
- Более тонкий, по сравнению с грамположительными бактериями, пептидогликановый слой
- Наружная мембрана содержит липополисахариды (состоит из липида А, полисахаридного ядра и антигена О снаружи и из фосфолипидов изнутри)
- В наружной мембране присутствуют порины,функционирующие подобно порам для определённых молекул
- S-слой прикреплен к наружной мембране, а не к пептидогликановому слою
- Если есть жгутик, он имеет четыре поддерживающих кольца, а не два
- Отсутствуют тейхоевая и липотейхоевая кислоты
- Обычно не образуют спор (примечательным исключением является Coxiella burnetii, образующая спороподобные структуры)
- Липопртеины прикреплены непосредственно к полисахаридной основе.
- Большинство содержат липопротеин Брауна, который связывает наружную мембрану и цепочки пептидоглюканов ковалентной связью
Химический состав и структура клеточных покровов
Далее рассмотрены слои клеточных покровов грамотрицательных прокариотов, начиная с самого внутреннего:
Клеточная стенка
У грамотрицательных эубактерий строение клеточной стенки намного сложнее, чем у грамположительных. В ее состав входит гораздо большее число макромолекул разного химического типа. Пептидогликан образует только внутренний слой клеточной стенки, неплотно прилегая к ЦПМ. Для разных видов грамотрицательных эубактерий содержание этого гетерополимера колеблется в широких пределах. У большинства видов он образует одно- или двухслойную структуру, характеризующуюся весьма редкими поперечными связями между гетерополимерными цепями.
Некоторые скользящие бактерии (миксобактерии, флексибактерии) способны в процессе перемещения по твердому субстрату периодически менять форму клеток, например путем изгибания, что говорит об эластичности их клеточной стенки, и в первую очередь ее пептидогликанового слоя. Электронно-микроскопическое изучение, однако, обнаружило у них клеточную стенку, типичную для грамотрицательных эубактерий. Наиболее вероятное объяснение гибкости клеточной стенки этих бактерий — чрезвычайно низкая сшитость ее пептидогликанового компонента. [3]
Периплазматическое пространство
Появление у грамотрицательных эубактерий дополнительной мембраны в составе клеточной стенки фактически привело к созданию обособленной полости (периплазматического пространства), отграниченной от цитоплазмы и внешней среды специфическими мембранами и несущей важную функциональную нагрузку. [3] Периплазматическое пространство, куда погружен пептидогликановый слой. заполнено раствором, в состав которого входят специфические белки, олигосахариды и неорганические молекулы. Периплазматические белки представлены двумя типами: транспортными белками и гидролитическими ферментами.

Было обнаружено также, что многие бактерии способны в больших количествах вырабатывать ферменты (гликозидазы, протеазы, липазы и проч.), гидролизующие все типы полимерных молекул. Последними могут быть как молекулы, синтезируемые самой клеткой, так и чужеродные, попавшие в клетку извне. Отрицательные последствия гидролиза собственных молекул (самопереваривание) очевидны. В то же время прокариоты нуждаются в гидролитических ферментах, так как это расширяет круг используемых ими веществ, включая в него полимеры разного типа. Становится понятна необходимость изолирования этих ферментов от цитоплазматического содержимого. Грамположительные эубактерий выделяют гидролитические ферменты во внешнюю среду, у грамотрицательных — они локализованы в периплазматическом пространстве.
Внешняя мембрана
Снаружи от пептидогликана располагается дополнительный слой клеточной стенки — наружная мембрана. Она состоит из фосфолипидов, типичных для элементарных мембран, белков, липопротеина и липополисахарида. Специфическим компонентом наружной мембраны является липополисахарид сложного молекулярного строения, занимающий около 30—40% ее поверхности и локализованный во внешнем слое.

Белки наружной мембраны можно разделить на основные и минорные. Основные белки представлены небольшим числом различных видов, но составляют почти 80% всех белков наружной мембраны. Одна из функций этих белков — формирование в мембране гидрофильных пор диаметром примерно 1-15 нм и длинной 50-70 нм, наклонённых к поверхности клеточной стенки по углом 30–40°. Через них осуществляется неспецифическая диффузия молекул с массой до 600—900 Да. Это означает, что через такие поры могут проходить сахара, аминокислоты, небольшие олигосахариды и пептиды. Белки, пронизывающие наружную мембрану насквозь и образующие гидрофильные поры, называют поринами. Минорные белки наружной мембраны представлены гораздо большим числом видов. Их основная функция — транспортная и рецепторная. Примером минорных белков могут служить белки, ответственные за специфический транспорт в клетку железосодержащих соединений.
Разнообразные функции выполняют макромолекулы, локализованные частично или полностью на внешней стороне клеточной стенки, контактирующей с окружающей средой; это специфические рецепторы для фагов и колицинов; антигены; макромолекулы, обеспечивающие межклеточные взаимодействия при конъюгации, а также между патогенными бактериями и тканями высших организмов.
S-слой и осцилиновые фибриллы
S-слой или зубчатый слой — слой из волоскоподобных фибрил, находится за пределами наружной мембраны в клеточной стенке цианобактерий способных к скольжению. Похожие на волски фибриллы верхнего слоя состоят из стержнеподобного гликопротеина, называемого осцилин.
Скольжение происходит посредством секреции слизи через поры на внешнюю сторону клеточных покровов. [4] Слизь проходит вдоль поверхности из осцилиновых фибрил наружного слоя клетки и по расположенному рядом субстрату, продвигая фибриллы вперёд. Совокупность организованных волоскоподобных фибрилл таким образом, действует как пассивный винт, в то время как слизь проходит по их поверхности в процессе скольжения. [5]
Классификация
Наряду с формой клетки, окрашивание по Граму это быстрый диагностический метод, который ранее использовался для группирования видов бактерий в подотделы.
Основываясь на окраске по Граму, ныне ликвидированное царство Monera было разделено на четыре дивизиона: Firmacutes (+), Gracillicutes (-), Mollicutes (0) и Mendocutes (var.). [6] Но начиная уже с 1987 года, монофилия грамотрицательных бактерий была поставлена под сомнение, а позже и полностью опровергнута бимолекулярными исследованиями. [7] Однако, некоторые авторы, такие как Кавалир-Смит, всё ещё считают их монофелитической кладой и относят к подцарству "Negibacteria". [8]
Строение внешней клеточной мембраны и классификация бактерий
Важно указать на то, что хотя бактерии традиционно делятся на две основные группы, Грамположительные и Грамотрицательные, такая классификация является двусмысленной и условной, поскольку может относится к трём совершенно разным аспектам (результат окрашивания, организация клеточных покровов, таксономическая группа), которые необязательно совпадают для всех видов бактерий. [9] [10] [11] [12] Реакция на окрашивание грамположительных и грамотрицательных бактерий не является надёжной характеристикой также и потому, что эти два вида бактерий не формирует филогенетически единую группу. [9] Как бы то ни было, хотя окрашивание по Граму и является эмпирическим критерием, в его основе лежат отчётливые различия в ультраструктуре и химическом составе двух главных видов прокариотических клеточных стенок, встречающихся в природе. Оба этих вида клеточных стенок отличаются друг от друга по наличию или отсутствию внешней липидной мембраны, которая является более надёжной и фундаментальной характеристикой бактериальных клеток. [9] [13] Все грамположительные бактерии окружены единственным слоем фосфолипидной мембраны и обычно имеют толстый слой (20-80 нм) из пептидогликанов (муреин и проч.), сохраняющий на себе краситель Грама. Ряд других бактерий, окружённых единственной мембраной, но окрашивающихся грамотрицательно из-за отсутствия пептидогликанового слоя (см. микоплазмы) или же своей неспособности удерживать краситель Грама вследствие специфического состава клеточной стенки, также находится в близком родстве с грамположительными бактериями. Для бактериальных клеток, окружённых одной клеточной мембраной был предложен термин Монодермные Бактерии или Монодермные Прокариоты. [9] [9] [13] В противоположность грамположительным бактериям, все архитипичные грамотрицательные бактерии, помимо цитоплазматической мембраны окружены ещё и внешней клеточной мембраной и содержат между ними крайне тонкий слой пептидогликанов (2-3 нм). Наличие внутренней и наружной мембран порождает новый клеточный компартмент — периплазматическое пространство. Таких бактерии/прокариоты были обозначены как Дидермные Бактерии. [9] [9] [13] Ещё одно важное различие межу монодермными и дидермными прокариотами — консервативный набор делеций в ряде важных белков (см. DnaK, GroEL). [9] [10] [13] [14] Из двух структурно различных групп прокариотических организмов, монодермные бактерии считаются предковой группой по отношению к дидермным. Основываясь на ряде наблюдений, включая тот факт, что грамположительные бактерии в большинстве своём продуценты антибиотиков, а грамотрицательные бактерии обычно к таковым устойчивы, было высказано предположение, что у наружной клеточной мембраны грамотрицательный бактерий существует защитный механизм против селективного действия антибиотиков. [9] [10] [13] [14] Некоторые бактерии, как например Deinococcus, окрашиваются грамположительно из-за толстого пептидогликанового слоя, но обладают наружной клеточной мембраной и считаются промежуточным звеном между монодермами (грамположительные) и дидермами (грамотрицательные). [9] [14] Дидермные бактерии в свою очередь могут быть подразделены на простых дидерм, у которых нет слоя липолисахаридов, архитипичных дидерм, чья наружная клточная мембрана содержит липолисахариды и собственно дидерм, наружная мембрана которых состоит из миколовой кислоты. [11] [12] [14] [15] В добавок, многие бактериальные таксоны (см. Negativicutes, Fusobacteria, Synergistetes и Elusimicrobia), которые являются частью типа Firmicutes или его близкородственной ветвью, также обладают дидермной структурой клетки. [12] [14] [15] Однако, консервативная делеция (CSI) в белке HSP60(GroEL) служит отличительным знаком всех традиционных типов грамотрицательных бактерий (т.е. Proteobacteria, Aquificae, Chlamydiae, Bacteroidetes, Chlorobi, Cyanobacteria, Fibrobacteres, Verrucomicrobia, Planctomycetes, Spirochetes, Acidobacteria, итд.) от атипичных дидерм и других типов монодермных бактерий (e.g. Actinobacteria, Firmicutes, Thermotogae, Chloroflexi, etc.). [14] Наличия CSI во всех секвенированых видах из обычных ЛПС-содержащих типов грамотрицательных бактерий, доказывает, что этот тип бактерий формирует монофилитическую кладу, и говорит в пользу того, что в этой группе ни у одного вида не происходила потеря внешней мембраны. Эти данные говорят против гипотезы о происхождении монодермных прокариот от дидермных путём потери внешней мембраны. [14]
Примеры видов
Протеобактерии большая группа грамотрицательных бактерий, включающая Escherichia coli (E. coli), Salmonella, Shigella, и прочих Enterobacteriaceae, Pseudomonas, Moraxella, Helicobacter, Stenotrophomonas, Bdellovibrio, уксуснокислых бактерий, Legionella и множество других. Другие примечательные группы грамотрицательных бактерий включают в себя цианобактерий, спирохет, зелёных серо и не-серо бактерий.
Связанные с медициной грамотрицательные Кокки включают в себя три организма, вызывающих заболевания, передающиеся половым путём (Neisseria gonorrhoeae), вызывающий менингит (Neisseria meningitidis), и респираторные симптомы (Moraxella catarrhalis).
Среди бацилл есть большое количество значимых для медицины видов. Некоторые из них в первую очередь взывают проблемы с дыханием, как например (Hemophilus influenzae, Klebsiella pneumoniae, Legionella pneumophila, Pseudomonas aeruginosa), в первую очередь проблемы с мочеиспусканием(Escherichia coli , Proteus mirabilis, Enterobacter cloacae, Serratia marcescens), и желудочно-кишечного тракта(Helicobacter pylori, Salmonella enteritidis, Salmonella typhi).
Грам-отрицательных бактерий, связанных с внутрибольничные инфекции, включая Acinetobacter baumannii, вызывающий бактериемии, вторичный менингит, и вентилятор-ассоциированной пневмонии при лечении в отделении интенсивной терапии.
Значение для Медицины
Одна из нескольких уникальных характеристик грамотрицательных бактерий это структура внешней клеточной мембраны. Внешний лист мембраны включает в себя комплекс липополисахаридов, липидные части которых выполняют роль эндотоксинов. Если эндотоксины попадают в транспортную систему, происходит интоксикация, с последующим увеличением температуры, повышением частоты дыхания, и понижением кровяного давления. Всё это может привести к токсическому шоку и последующему летальному исходу.

Разнообразие форм клеток прокариот не является (по крайней мере не всегда) случайным феноменом эволюции этих организмов. Исследования показали, что форма бактерий может быть обусловлена физическими законами среды обитания: в вязкой среде эффективнее перемещаются микрообитатели спиральные формы, а следовать направлению лучше могут изогнутые вибрионы и т.д. Согласно расчетам наиболее удобна для микроскопических одноклеточных прокариот форма палочек, которые благодаря своей форме могут противостоять броуновскому движению в жидкостях, имеют эффективное соотношение поверхности к объему клетки и могут закрепляться на субстрате…Авторы статьи проанализировали исследования эволюции и связи с экологией формы клеток бактерий.
Форма и размер бактериальных клеток, как и свойства их клеточной стенки (что отразилось на широко известном делении бактерий на грамположительных и грамотрицательных) – одни из самых первых признаков, использованных для классификации этих организмов. Разнообразие форм клеток и в то же время постоянство формы клеток на видовом уровне (за некоторым обсуждаемом ниже исключением) позволили довольно подробно и точно определять таксономическую принадлежность бактерий. Однако причины возникновения разнообразия формы и ее стабильность внутри разного уровня таксонов прокариот долго оставались загадкой. Новые методы исследований – электронная микроскопия, методы молекулярной биологии и биохимии, а также исследования физических закономерностей и математическое моделирование помогли установить ряд факторов, определяющих внешнее строение бактерий. В обсуждаемой статье авторы представили анализ исследований связи формы клеток бактерий с их экологией и эволюцией.

Несмотря на то, что основными являются три типа клеток бактерий (заглавная иллюстрация) – сферическая, палочковидная и спиральная – специалисты выделяют довольно большое разнообразие других форм (рис. 1). Известно, что бактерии по строению клеточной стенки можно разделить на два типа (рис. 1, 2). Строение оболочки (клеточной стенки бактерий) в значительной степени связано с ее формой. Среди определяющих форму бактерий факторов на данным момент выделяют несколько основных:
- наличие/отсутствие внешней мембраны (у грамотрицательных бактерий);
- относительная толщина пептидогликанового слоя;
- особенности строения продольных пептидных сшивок между гликановыми нитями, ориентированными перпендикулярно длинной оси клетки: у грамотрицательных образуются напрямую, а у грамположительных через дополнительный мостик.

Ряд авторов отмечают, что морфологическое разнообразие грамотрицательных бактерий выше, чем таковое грамположительных (см. рис. 1). Среди грамположительных бактерий преобладают палочки, часто встречаются кокки и нитевидные формы, а вот изогнутые и спиральные формы очень редки. Палочки также преобладают и среди грамотрицательных бактерий, но второе и третье места по распространенности делят изогнутые и спиральные формы. А вот кокки и одноклеточные нитчатые формы среди грамотрицательных бактерий редки, хотя некоторые палочки и спиральные бактерии в определенных условиях могут приобретать округлую форму, например, в стационарной фазе культивирования и при неблагоприятных условиях.

Многочисленные нитевидные формы это производные палочек, длина стенок которых во много раз превышает диаметр клетки. Нитевидная форма одна из стратегий избегания хищничества со стороны простейших. Длинные, разветвленные формы получают возможность функционально дифференцировать клетку, что способствует более эффективному питанию в случае дефицита определенных элементов питания.
Извитые (спиральные) формы. Бактерии могут становится извитыми разными способами в разных эволюционных линиях прокариот. Например, Helicobacter pylori, вызывающий язву желудка, особыми ферментами (группы Csd) контролируемо разрезает сшивки между нитями в пептидогликановом слое, благодаря чему правильно организованный цилиндр клеточной стенки скручивается в спираль (рис. 4). Интересно, что грамположительные бактерии не имеют ферментов этой группы, к тому же их сшивки между нитями содержат дополнительные (пентаглициновые) мостики, а не сшиты напрямую, как у грамотрицательных бактерий. Эти обстоятельства в некоторой степени объясняют редкость спиральных форм среди грамположительных бактерий.

Рис. 4. Схематическое изображение пептидогликанового слоя Helicobacter pylori (по: Sycuro et al., 2010). Ножницами указаны сайты возможного гидролиза (разрыва) пептидных связей эндопептидазами Csd. (Иллюстрация из обсуждаемой статьи)
Изогнутые формы– вибрионы – можно рассматривать как короткие спиральные формы. Но у вибрионов есть покрайней мере еще один способ изогнуться: при помощи "тормозящего" белка кресцетина CreS (см. выше). Ряд исследований показал, что изогнутая форма вибриона способствует активному движению в жидкости и активному поиску лучшего места (хемотаксису).

Помимо общей формы клетки бактерии также могут иметь дополнительные внешние морфологические элементы – жгутики, мембраны, выросты, ножки – отражающие способности прокариот специфически приспосабливаться к определенным условиям жизни, моделируя для себя субнишевые (в экологическом смысле) пространства (рис. 5). Понятно, что целый ряд факторов, таких как свойства среды, способ питания, хищничество со стороны простейших, взаимодействие с субстратом и др. определяют эволюцию формы клеток бактерий. Интересно, что один и тот же тип клеток, как и дополнительных внешних морфологических приспособлений, может обеспечиваться разными структурными элементами оболочки и молекулярными механизмами в ходе эволюции разных таксонов.
Читайте также: