
Сам метаболизм у растений кратко и понятно
Обновлено: 05.07.2024
Оглянитесь вокруг! Пожалуй, в каждом доме есть хотя бы одно зеленое растение, а за окном несколько деревьев или кустарников. Благодаря сложному химическом процессу происходящего в них фотосинтеза стало возможно зарождение жизни на Земле и существование человека. Разберем историю его открытия, суть процесса и реакции, которые протекают в разных фазах.
История открытия фотосинтеза
В настоящее время школьники впервые знакомятся со сложными процессами фотосинтеза уже в 6 классе.
Первым и очевидным ответом было предположение, что из земли. Однако, в далеком 1600 году фламандский ученый Ян Батист ван Гельмонт решил проверить влияние почвы на рост растений и провел уникальный в своей простоте опыт. Естествоиспытатель взял веточку ивы и бочку с почвой. Предварительно их взвесил. А затем посадил отросток ивы в бочку с почвой.
Долгие пять лет ван Гельмонт поливал молодое деревце лишь дождевой водой. А через пять лет выкопал деревце, и вновь взвесил отдельно деревце и отдельно почву. Каково же было его удивление, когда весы показали, что деревце увеличило свой вес практически в тридцать раз, и совсем не походило на тот скромный прутик, что был посажен в кадку. А вес почвы уменьшился всего на 56 граммов.
Ученый сделал вывод. что почва практически не дает строительного материала растениям, а все необходимые вещества растение получает из воды.
Одним из тех, кто попытался возразить этой теории был М.В. Ломоносов. И строил он свои возражения на том, что на пустых, скудных северных землях с редкими дождями растут высокие, мощные деревья. Михаил Васильевич предположил, что часть питательных веществ растения впитывают через листья, но доказать свою теорию экспериментально он не смог.
И как часто бывает в науке, помог его величество случай.
Однажды нерадивая мышь, решившая поживиться церковными запасами, случайно перевернула банку и оказалась в ловушке. И через некоторое время погибла. К нашей удаче, эту мышь в банке обнаружил Джозеф Пристли, который был не просто священником, а по совместительству ученым-химиком, и очень интересовался химией газов и способами очистки испорченного воздуха. И тут церковным мышам не повезло. Они стали участницами различных опытов английского ученого.
Джозеф Пристли ставил под одну банку горящую свечу, а в другую сажал мышь. Свеча тухла, грызун погибал.
В наше время его самого зоозащитники посадили бы в банку, но в далеком 1771 году ученому никто не помешал продолжить свои опыты. Пристли посадил мышь в банку, где до этого потухла свеча. Животное погибло еще быстрее.
И тогда Пристли сделал вывод, что раз все живое на Земле до сих пор не погибло, Бог (мы же помним, что Пристли был священником), придумал некий процесс, чтобы воздух вновь был пригоден для жизни. И скорее всего, основная роль в нем принадлежит растениям.
Чтобы доказать это, ученый взял воздух из банки где погибла мышь, и разделил его на две части. В одну банку он поставил мяту в горшочке. А другая банка ждала своего часа. Через 8 дней растение не только не погибло, а даже выпустило несколько новых побегов. И он опять посадил грызунов в банки. В той, где росла мята — мышь была бодра и закусывала листиками. А в той, где мяты не было — практически моментально лежала дохлая мышиная тушка.

Опыты Пристли вдохновили ученых, и во всем мире начали отлавливать мелких грызунов и пытаться повторить его эксперименты.
Но мы же помним, что Пристли был священником и весь день, до вечерней службы мог заниматься исследованиями.
А Карл Шееле, аптекарь из Швейцарии, экспериментировал в домашней лаборатории в свободное от работы время, т.е. по ночам, и мыши дохли у него независимо от присутствия мяты в банке. В результате его экспериментов получалось, что растения не улучшают воздух, а делают его непригодным для жизни. И Шееле обвинил Пристли в обмане научной общественности. Пристли не уступил, и в результате противостояния ученых было установлено, что для восстановления воздуха растениям необходим солнечный свет.
Именно эти опыты положили начало изучению фотосинтеза.
Исследование фотосинтеза стремительно продолжалось. Уже в 1782 году, спустя всего лишь 11 лет после исследований Пристли, швейцарский ботаник Жан Сенебье доказал, что органоиды растений разлагают углекислый газ в присутствии солнечного света. И практически еще сто лет провальных и удачных экспериментов понадобилась ученым разных специальностей, чтобы в 1864 году немецкий ученый Юлиус Сакс смог доказать, что растения потребляют углекислый газ и выделяют кислород в соотношении 1:1.

Значение фотосинтеза для жизни на Земле
И теперь становится понятна важность процесса фотосинтеза для жизни на земле. Именно благодаря этому сложному химическом процессу стало возможно зарождение жизни на земле и существование человека.
Кто-то может возразить, что на Земле есть места, где не растут ни деревья ни кустарники, например, пустыни или Арктические льды. Ученые доказали, что доля кислорода, выделяемого зеленой массой лесов, кустарников и трав — т. е. растений, что обитают на поверхности суши, составляет всего около 20% газообмена, а 80% кислорода приходится на мельчайшие морские и океанские водоросли, которые потоками воздуха переносятся по всей планете, позволяя дышать животным в экстремальных, практически лишенных растительности регионах нашей удивительной планеты.
Благодаря фотосинтезу вокруг нашей планеты сформировался защитный озоновый экран, защищающий все живое на земле от космической и солнечной радиации, и живые организмы смогли выйти на сушу из глубин океана.
К сожалению, в настоящее время кислород потребляют не только живые существа, но и промышленность. Уничтожаются тропические леса, загрязняются океаны, что приводит к снижению газообмена и увеличению дефицита кислорода.
Определение и формула фотосинтеза
Определение и формула фотосинтеза
Схема фотосинтеза, на первый взгляд, проста:
Вода + квант света + углекислый газ → кислород + углевод
или (на языке формул):
Если копнуть поглубже и посмотреть на лист в электронный микроскоп, выяснится удивительная вещь: вода и углекислый газ ни в одной из структурных частей листа непосредственно друг с другом не взаимодействуют.
Фазы фотосинтеза
К фотосинтезу способны не только растения, но и многие одноклеточные животные благодаря специальным органоидам, которые называются хлоропласты.
Хлоропласты — это пластиды зеленого цвета фотосинтезирующих эукариот. В состав хлоропластов входят:
- две мембраны;
- стопки гранов;
- диски тилакоидов;
- строма — внутреннее вещество хлоропласта;
- люмен — внутреннее вещество тилакоида.
Сложный процесс фотосинтеза состоит из двух фаз: световой и темновой. Как понятно из названия, световая (светозависимая) фаза происходит с участием квантов света. Название темновая фаза вовсе не означает, что процесс происходит в темноте. Более точное определение — светонезависимая. Т.е. для реакций, происходящих в этой этой фазе, свет не нужен, а протекает она одновременно со световой, только в других отделах хлоропласта.
Многие делают ошибку, говоря, что в процессе фотосинтеза происходит производство растениями такого необходимого человечеству кислорода. На самом деле фотосинтез — это синтез углеводов (например, глюкозы), а кислород — лишь побочный продукт реакции.
Световая фаза фотосинтеза
Световая фаза фотосинтеза происходит на мембранах тилакоидов. Фотон света, попадая на хлорофилл, возбуждает его и происходит выделение электронов и скопление отрицательно заряженных электронов на мембране. После того, как хлорофилл потерял все свои электроны, квант света продолжает воздействовать на воду, вызывая фотолиз Н2О.
Положительно заряженные протоны водорода накапливаются на внутренней мембране тилакоида.
Получается такой бутерброд: с одной стороны отрицательно заряженные электроны хлорофилла, с другой – положительно заряженные протоны водорода, а между ними – внутренняя мембрана тилакоида.
Гидроксильные ионы идут на производство кислорода:
Когда количество протонов водорода и электронов достигает максимума, запускается специальный переносчик — АТФ-синтаза. АТФ-синтаза выталкивает протоны водорода в строму, где их подхватывает специальный переносчик никотинамиддинуклеотидфосфат или сокращенно НАДФ. НАДФ — специфический переносчик протонов водорода в реакциях углеводов.
Прохождение протонов водорода через АТФ-синтазу сопровождается синтезом молекул АТФ из АДФ и фосфата или фотофосфорилированием, в отличие от окислительного фосфорилирования.
На этом световая фаза фотосинтеза заканчивается, а НАДФН+ и АТФ переходят в темновую фазу.
Повторим ключевые процессы световой фазы фотосинтеза:
- Фотон попадает на хлорофилл с выделением электронов.
- Фотолиз воды.
- Выделение кислорода.
- Накопление НАДФН+.
- Накопление АТФ.
Существует еще одна группа растений, имеющая особенности в осуществлении первичной фиксации СО2. Это кактусы и суккуленты из семейства Толстянковые (Crassulaceae). Поэтому этот тип фотосинтеза получил название Crassulaceae acid metabolism или САМ-метаболизм (САМ-путь).
При функционировании САМ-пути устьица растений закрыты днем, когда интенсивность транспирации максимальна, и открыты ночью, когда потери воды минимальны. Это предохраняет растения от излишней потери воды, повышает их устойчивость к засухе. Химизм фиксации СО2 при САМ-пути сходен с С4-путем, однако если у С4-растений фотосинтез разделен в пространстве, то при САМ-пути — во времени. У САМ-растений фиксация СО2 происходит в ночное время, когда устьица открыты (рис. 8).
Осуществление фотосинтеза по такому пути позволяет растениям максимально экономить воду и поддерживать процесс фотосинтеза в условиях острого водного дефицита. Однако САМ-путь не может обеспечить высокой продуктивности растений, поэтому данные растения медленно растут и не могут конкурировать с С3- и С4-растениями при менее экстремальных условиях. Возможна смена путей фиксации СО2. При достаточном количестве воды растения с САМ-метаболизмом могут переходить на С3-путь (Mesembryanthemum crystallinum).
В научной литературе имеются указания о наличии и других путей связывания СО2. Однако они еще недостаточно изучены. Известно, что на всех этапах фотосинтетического цикла промежуточные продукты могут претерпевать различные превращения в зависимости от условий среды. Именно это является основой для образования разнообразных продуктов фотосинтеза.
Суккуленты (роды Crassula, Bryophyllum и др.) также приспособились осуществлять фотосинтез в условиях резко засушливого климата. Для них характерен суточный цикл метаболизма С4-кислот с образованием яблочной кислоты ночью. В соответствии с английским выражением Crassulacean Acid Metabolism (САМ) этот тип фотосинтеза часто сокращенно называют САМ-метаболизм. Устьица этих растений днем обычно закрыты, что предотвращает потерю воды, и открываются ночью. С02 поступает в листья, где при участии содержащейся в цитоплазме ФЕП-карбоксилазы взаимодействует с фосфоенолпируватом, образуя оксалоацетат. Источник ФЕП-карбоксилазы взаимодействует с фосфоенолпируватом, образуя оксалоацетат. Источником ФЕП служит крахмал. То же самое происходит и с С02, который освобождается в клетках в процессе дыхания. Образовавшийся оксалоацетат восстанавливается под действием NADH-зависимой малатдегидрогеназы до яблочной кислоты, которая накапливается в вакуолях клеток листа.
Это приводит к закислению клеточного сока в ночное время. Как и у С4-растений, оксалоацетат может быть источником аспартата, однако этот путь здесь менее выражен.
Днем в условиях высокой температуры, когда устьица закрыты, малат транспортируется из вакуолей в цитоплазму и там декарбоксилируется при участии малатдегидрогеназы декарбоксилирующей (малик-энзима) с образованием С02 и пирувата. С02 поступает в хлоропласты и включается в них в цикл Кальвина, участвуя в синтезе сахаров.
Таким образом, у растений с фотосинтезом по типу толстянковых много общего с С4-путем фотосинтеза. Однако при САМ-метаболизме фиксация С02 с образованием малата (ночью) и декарбоксилирование малата с высвобождением С02 и пирувата (днем) разделены во времени. У С4-растений эти же реакции разграничены в пространстве: первая протекает в хлоропластах мезофилла, вторая — в клетках обкладки. При достаточном количестве воды ряд растений с метаболизмом по типу толстянковых могут вести себя как С3-растения. В свою очередь некоторые растения с С3-путем фотосинтеза при недостатке воды проявляют черты САМ-метаболизма.
12. Физиологические особенности С₄- растений
Исследования показали, что в растениях, в которых процесс фотосинтеза протекает по С4-пути, имеются два типа клеток и хлоропластов:
1) мелкие гранальные пластиды в клетках мезофилла листа;
2) крупные пластиды, часто лишенные гран, в клетках обкладки, окружающих сосудистые пучки.
Клетки обкладки имеют утолщенные клеточные стенки, содержат большое количество хлоропластов и митохондрий, расположены вокруг сосудистых пучков в 1 или 2 слоя.
У таких растений специфичное анатомическое строение листа. У них два типа фотосинтезирующих клеток: клетки обкладки, радиально покрывающие проводящие пучки в листе, и клетки мезофилла столбчатой и губчатой ткани. Это может служить диагностическим признаком на определение С4-растений. В структуре этих клеток также есть различия: в клетках обкладках в хлоропластах отсутствуют граны, они агранальные, а в клетках мезофилла все хлоропласты гранальные
К группе с С4-путем фотосинтеза относятся сахарный тростник, кукуруза, сорго и др. Эти растения устойчивы к засолению. Эффективность использования воды , т.е. отношение массы ассимилированного СО2 к массе воды, израсходованной при транспирации, у С4 растений зачастую вдвое выше, чем у С3-растений. Таким образом, С4-растения имеют преимущества перед С3-растениями в засушливых местах обитания, благодаря высокой интенсивности фотосинтеза даже при закрытых устьицах. Фиксация СО2 с участием ФЕП и образование малата(аспартата) служит как бы насосом для поставки СО2 в хлоропласты обкладки, функционирующие по С3 пути.
13. Фотодыхание (химизм, структурная организация процесса) и функциональная роль
Путь восстановления СО2 при фотосинтезе М. Кальвин изучал при относительно высоких концентрациях углекислого газа. При более низких концентрациях СО2, приближающихся к атмосферным, в хлоропласте образуется значительное количество фосфогликолевой кислоты. Это связано с тем, что в условиях низкого парциального давления СО2 и высокого парциального давления О2 РДФ-карбоксилаза может присоединять к рибулозо-1,5-дифосфату кислород; это значит, что она может выполнять не только карбоксилирующую, но и оксигенерирующую функции. Присоединение кислорода к молекуле рибулозодифосфата приводит к тому, что вместо двух молекул ФГК образуется одна молекула ФГК и одна молекула фосфогликолата. Последняя во время транспорта через мембрану хлоропласта дефосфорилируется при участии фосфогликолатфосфатазы и превращается в гликолат, который поступает в специальную органеллу-периксисому.
В последней содержится каталаза – фермент, разлагающий перекиси. В периксисоме гликолат вступает в реакцию с кислородом воздуха, в результате образуется глиоксилат и перекись водорода. Реакцию катализирует гликолатоксидаза.
Перекись сразу разрушается каталазой на воду и кислород, а глиоксилат в результате реакции трансаминирования, которая проходит при участии глиоксилатглицинаминотрансферазы, превращается в глицин, который перемещается в митохондрию. Тут из двух молекул глицина при участии серинглиоксилатаминотрансфераза синтезируется одна молекула серина, образуется СО2, аммиак и восстанавливается НАД + . СО2 или выделяется в атмосферу, это значит теряется для растения, или вновь может связаться в С3-цикле.
Предполагают, что образующийся НАДН может быть связан с электрон-транспортной цепью дыхания, что приводит к синтезу АТФ при его окислении.
Серин из митохондрий транспортируется вновь в периксисому, где после дезаминирования превращается в оксипируват, потом в глицерат, и наконец, в ФГК. ФГК – конечный продукт этого цикла может поступать в С3-цикл. Цикл превращения гликолата в триозофосфат(ФГК) называется гликолатным. Образовавшийся аммиак не выделяется в окружающую среду, а связывается в виде глутамина.
Так как образуются кислоты, содержащие 2 атома углерода, то этот цикл получил название С2-цикла. С другой стороны, так как во время этих превращений происходит поглощение кислорода и выделение СО2, то этот процесс получил название фотодыхания.
Фотодыхание (или световое дыхание) – это индуцированное светом поглощение О2 и выделение СО2, которое наблюдается в зеленых клетках. Интенсивность фотодыхания увеличивается при повышении освещенности.
Хотя органеллой фотодыхания считается периксисома, в этом цикле непосредственное участие принимают хлоропласты и митохондрии. Интенсивное фотодыхание может снизить продуктивность С3-растений на 30 %, так как при фотодыхании происходит не поглощение СО2, а его потери. Из каждых 10 молекул риболозо-1,5-дифосфата, образовавшегося в С3-цикле и включившихся в фотодыхание, одна разрушается:

Низкую интенсивность фотодыхания в С4-растениях объясняют тем, что в них высокая концентрация СО2 в клетках-обкладки. В этих растениях РДФ-карбоксилаза находится только в клетках-обкладки, где и происходит процесс.
Фотодыхание рассматривают как одну из необходимых функций зеленого растения. Во-первых, фотодыхание, как и С3-цикл, САМ-цикл и другие процессы, является источником метаболитов для разных синтезов. Во-вторых, что при образовании серина и глицина восстанавливается митохондриальная НАД + . В-третьих, выявлена связь фотодыхания с ассимиляцией азота. И, наконец, появилась теория, в соответствии с которой излишки продуктов цикла Кальвина служат сигналом, который увеличивает фотосинтез. Таким образом, соотношение фотосинтеза и фотодыхания выполняют регуляторную роль в растениях.
Как мы видели, в фотосинтезирующих клетках в световых реакциях (гл. 19) образуется O2 (из Н2O), а в светонезависимых реакциях потребляется СО2, поэтому при фотосинтезе происходит изменение газового состава окружающей среды — поглощение СO2 и высвобождение O2.
В темноте растения осуществляют также митохондриальное дыхание — окисление субстратов до СO2 и восстановление O2 до Н2O. Однако у растений происходит и другой процесс, при котором, как и при митохондриальном дыхании, потребляется O2 и выделяется СO2, и который, как фотосинтез, управляется светом. Этот процесс фотодыхания — дорогостоящая реакция фотосинтеза по причине недостаточной специфичности фермента рубиско. В следующем разделе мы опишем эту побочную реакцию и стратегии, которые используются растениями для минимизации ее метаболических последствий.
Фотодыхание — следствие оксигеназной активности рубиско
Рубиско не имеет абсолютной специфичности в отношении СO2 как субстрата. Молекулярный кислород (O2) конкурирует с СO2 за активный центр, и через каждые три-четыре оборота рубиско катализирует одну конденсацию O2 с рибулозо-1,5-бисфосфатом с образованием 3-фосфоглицерата и 2-фосфогликолата (рис. 20-20), метаболически бесполезного продукта. В этом и проявляется оксигеназная активность этого фермента, которая отражена в его полном названии: рибулозо-1,5-бисфосфат- карбоксилаза/оксигеназа. В результате реакции с O2 не происходит фиксации углерода, и в итоге такая реакция оказывается обузой для клетки; усвоение атомов углерода из 2-фосфогликолата требует от клетки много энергии и высвобождает некоторое количество ранее фиксированного углекислого газа.
энергии. Температурная зависимость относительной растворимости O2 и СO2 ([O2] / [СO2]) круто возрастает при высоких температурах. Кроме того, сродство рубиско к СO2уменьшается с увеличением температуры, усиливая вероятность катализа расточительной оксигеназной реакции. А так как СO2 потребляется в реакциях ассимиляции, соотношение между O2 и СO2 в воздушном пространстве, которое окружает листья, увеличивается, что еще больше способствует оксигеназной реакции.
Рис. 20-20. Оисигеназная активность рубиско. Рубиско может включать O2 вместо СO2 в рибулозо-1,5-бисфосфат. Образующийся нестабильный интермедиат распадается на 2-фосфогликолат (он перерабатывается, как показано на рис. 20-21) и 3-фосфоглицерат, который может включаться в цикл Кальвина.

Усвоение фосфоглицерата обходится клетке весьма дорого
Гликолатный путь осуществляет превращение двух молекул 2-фосфогликолата в молекулу серина (3 атома углерода) и молекулу СO2 (рис. 20-21). В хлоропласте фосфатаза переводит 2-фосфогликолат в гликолат, который экспортируется в пероксисому. Там гликолат окисляется молекулярным кислородом, и получившийся альдегид (глиоксилат) подвергается трансаминированию с образованием глицина. В качестве побочного продукта реакции окисления гликолата образуется пероксид водорода, который распадается под действием пероксидазы пероксисомы. Глицин транспортируется из пероксисомы в митохондрию, где подвергается окислительному декарбоксилированию под действием глициндекарбоксилазного комплекса. Этот фермент похож по структуре и механизму действия на митохондриальные ферменты, с которыми мы уже сталкивались: пируватдегидрогеназный и α-кетоглутаратдегидрогеназный комплексы (гл. 16). Глициндекарбоксилазный комплекс окисляет глицин до СO2 и NН3 с одновременным восстановлением NАD+ до NАDН и переносом оставшегося атома углерода от глицина на кофактор тетрагидрофолат (рис. 20-22). Получившаяся одноуглеродная единица затем переносится серин-гидроксиметилтрансферазой, синтезирующей серин, с тетрагидрофолата на второй глицин. Итоговая реакция, катализируемая глициндекарбоксилазным комплексом и серин- гидроксиметилтрансферазой приведена ниже.
2 Глицин + NАD + + Н2O —> серин + СO2 + NН3 + NАDН + Н +
Рис. 20-21. Гликолатный путь. Путь усвоения 2-фос- фоглицерата (выделен розовым) через его превращение в серин и в конечном счете в 3-фосфоглицерат, в котором участвуют три компартмента клетки. Гликолат, образующийся при дефосфорилировании 2-фосфогликолата в хлоропластах, окисляется в пероксисомах до глиоксилата и затем трансаминируется до глицина. В митохондриях две молекулы глицина конденсируются с образованием серина, и СO2 высвобождается в процессе фотодыхания (выделен зеленым). Эта реакция катализируется глициндекарбоксилазой, ферментом, присутствующим в высоких концентрациях в митохондриях С3-растений (см. текст). Серин превращается в гидроксипируват и затем в глицерат в пероксисомах; глицерат попадает в хлоропласты, фосфорилируется и включается в цикл Кальвина. Кислород (окрашен голубым) потребляется в процессе фотодыхания в две стадии.
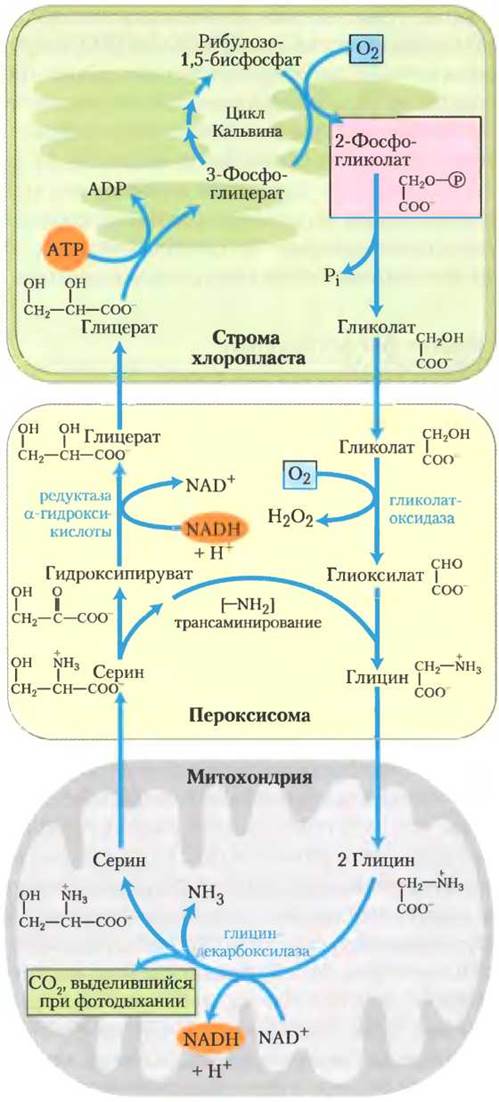
При ярком солнечном свете утечка через гликолатный путь может быть очень высокой. Растение производит таким способом в 5 раз больше СO2, чем обычно образуется в результате всех окислительных реакций в цикле трикарбоновых кислот. Для обеспечения этого потока митохондрии растений содержат потрясающе огромное количество глициндекарбоксилазного комплекса: в листьях гороха и шпината четыре белка этого комплекса составляют половину всех белков митохондриального матрикса! В нефотосинтезирующих частях растения, таких как картофельные клубни, митохондрии содержат глициндекарбоксилазный комплекс в очень низкой концентрации.
Совместная активность рубиско-оксигеназы и гликолатного пути потребляет O2 и производит СO2 — откуда и название фотодыхание. Возможно, этот путь лучше назвать цикл фотосинтетического окисления углерода или С2-путь; эти названия не вызывают ассоциации с дыханием в митохондрии. В отличие от митохондриального дыхания фотодыхание не сохраняет энергию и может ингибировать образование чистой биомассы почти на 50%. Эта неэффективность ведет к эволюционным адаптациям, особенно у растений, которые развиваются в теплом климате.
У С4-растений фиксация СO2 и активность рубиско пространственно разделены
У многих растений, которые растут в тропиках, а также теплолюбивых культур, происходящих из тропиков, таких как кукуруза, сахарный тростник и сорго, развился механизм, благодаря которому они избегают проблемы невыгодного фотодыхания. Стадии включения СO2 в трехуглеродный 3-фосфоглицерат предшествует несколько стадий, в том числе временная фиксация СO2 в четырехуглеродное соединение. Растения, использующие этот путь, называются С4-растениями, а сам процесс ассимиляции — С4-метаболизмом, или С4- путем. Растения, которые фиксируют СO2 по пути, где на первой стадии происходит реакция конденсация СO2 с рибулозо-1,5-бисфосфатом с образованием 3-фосфоглицерата, называются С3-растениями.
С4-Растения обычно развиваются при большой интенсивности света и высокой температуре. Их отличает несколько важных характеристик: высокая скорость фотосинтеза, большая скорость роста, низкий уровень фотодыхания, малая скорость потери влаги и специализированная структура листа. В листьях С4-растений в фотосинтезе участвуют два типа клеток: мезофилл и клетки обкладки (рис. 20-23, а). Существуют три варианта С4-метаболизма, расшифрованные в 1960-х гг. Маршаллом Хэтчем и Роджером Слэком (рис. 20-23, б).
Рис. 20-22. Глициндекарбоксилазная система. Глициндекарбоксилаза митохондрий растений представляет собой комплекс из четырех типов субъединиц со стехиометрией P4H27T9L2. С белком Н ковалентно связан остаток липоевой кислоты, которая может подвергаться обратимому окислению. (1) — образование шиффова основания между пиридоксальфосфатом (PLP от англ. pyridoxal phosphate) и глицином, катализируемое белком Р (названным так потому, что он образует связь с PLP). (2) — белок Р катализирует окислительное декарбоксилирование глицина, высвобождая СО2; оставшаяся группа метиламина связана с одной из -SH-групп восстановленной липоевой кислоты. (3) — белок Т (который использует тетрагидрофолат (H4F) в качестве кофактора) высвобождает NH3 из метиламина и переносит оставшийся одноуглеродный фрагмент на тетрагидрофолат, образуя N 5, N10 -метилентетрагидрофолат. (4) — белок L окисляет две -SH- группы липоевой кислоты до дисульфидной связи, (5) передавая электроны через FAD на NAD + и завершая таким образом цикл. N 5 , N 10 -Метилентетрагидрофолат, образованный в этом процессе, используется серин-гидроксиметилтрансферазой для превращения молекулы глицина в серин, в результате регенерируется тетрагидрофолат, необходимый для реакции, катализируемой белком Т. L-Субъединица глициндекарбоксилазы идентична дигидролипоилдегидрогеназе (Е3) пируватдегидрогеназы и α-кетоглутаратдегидрогеназы (см. рис. 16-6).

У тропических растений первый интермедиат, в который включается 14 СО2, — четырехуглеродное соединение оксалоацетат. Эта реакция происходит в цитозоле клеток мезофилла листьев и катализируется фосфоенолпируват- карбоксилазой, субстратом для которой служит НСО3 - , а не СО2. Затем оксалоацетат либо восстанавливается до малата с расходом NADPH (см. рис. 20-26, б), либо превращается в аспартат в реакции трансаминирования:
Оксалоацетат + α-аминокислота —> L-аспартат + α-кетокислота
Малат или аспартат, синтезированный в клетках мезофилла, проникает в соседние клетки обкладки через плазмодесмы — образованные белками каналы, которые соединяют две растительные клетки и осуществляют движение метаболитов и даже малых белков между клетками. В клетках обкладки малат окисляется и декарбоксил ируется с образованием пирувата и СО2 под действием NADP-зависимой декарбоксилирующей малатдегидрогеназы (иногда называемой малик-ферментом, или яблочным ферментом) с восстановлением NADP + . У растений, использующих аспартат в качестве переносчика СО2, аспартат, поступивший в клетки обкладки, трансаминируется с образованием оксалоацетата и восстанавливается до малата, после чего СО2 высвобождается малатдегидро- геназой или ФЕП-карбоксикиназой. Как показали эксперименты с образцами, меченными 14 С, молекула СО2, высвобождающаяся в клетках обкладки, — это та же самая молекула СО2, которая изначально находилась в оксалоаце- тате в клетках мезофилла. Эта молекула СО2 снова фиксируется, теперь с помощью рубиско, в той же самой реакции, которая происходит у С3-растениях: происходит включение СО2в положение С-1 молекулы 3-фосфоглицерата.
Пируват, образовавшийся при декарбокси- лировании малата в клетках обкладки, транспортируется обратно, в клетки мезофилла, где превращается в фосфоенолпируват в необычной реакции, катализируемой пируват-фосфатдикиназой (рис. 20-23, б). Этот фермент называется дикиназой, потому что он, используя одну молекулу АТР, одновременно фосфорилирует две разные молекулы: пируват до фосфоенолпирувата и фосфат до пирофосфата. Впоследствии пирофосфат гидролизуется до фосфата, поэтому получается, что для регенерации фосфоенолпирувата используются две высокоэнергетические фосфатные группы АТР. Теперь этот фосфоенолпируват готов для включения следующей молекулы СО2 в клетки мезофилла.
Рис. 20-23. Ассимиляция углерода у С4-растений. В растениях тропических районов преобладает С4-путъ, проходящий в клетках мезофилла и клетках обкладки, а — электронная микрофотография, показывающая хлоропласта мезофилла и прилегающих клеток обкладки. Клетки обкладки содержат гранулы крахмала. Видны плазмодесмы, соединяющие две клетки. б — С4-путь ассимиляции СO2, который происходит с участием четырехуглеродного интермедиата.


ФЕП-карбоксилаза имеет высокое сродство к НСО3- (который в водном окружении образуется из СО2) и фиксирует СО2 более эффективно, чем рубиско. В отличие от рубиско этот фермент не использует О2 в качестве альтернативного субстрата, поэтому в этой реакции нет конкуренции между СО2 и О2. Таким образом, ФЕП- карбоксилазная реакция служит для фиксации СО2 в форме малата. Высвобождением СО2 из малата в клетках обкладки достигается высокая локальная концентрация углекислого газа для функционирования рубиско с почти максимальной скоростью и подавления ее оксигеназной активности.
В клетках обкладки СО2 включается в 3-фосфоглицерат, поэтому другие реакции цикла Кальвина происходят так, как было описано ранее. Таким образом, у С4-растений клетки мезофилла осуществляют ассимиляцию СО2 по С4- пути, а клетки обкладки синтезируют крахмал и сахарозу по С3-пути.
В С4-пути три фермента, которые регулируются светом, становятся более активными в дневное время суток. Малатдегидрогеназа активируется через тиоредоксин-зависимый восстановительный механизм, показанный на рис. 20-19; ФЕП-карбоксилаза активируется фосфорилированием серинового остатка; а пируват-фосфатдикиназа активируется дефосфорилированием. В последних двух случаях детали инициации светом процессов фосфорилирования или дефосфорилирования неизвестны.
У CAM-растений фиксация СО2 и активность рубиско разделены во времени
Суккулентные растения, такие как кактусы и ананас, которые в природе произрастают в очень жарком и очень засушливом климате, осуществляют другой вариант фотосинтетической фиксации СО2, уменьшающий потери воды через устьица, через которые СО2 и О2 должны попадать в ткань листа. Вместо пространственного разделения процессов начального захвата СО2 и его фиксации рубиско (как это делают С4-растения) они разобщают эти процессы во времени. Ночью, когда воздух холоднее и более влажный, устьица открыты и пропускают СО2, который затем фиксируется на оксалоацетате ФЕП-карбоксилазой. Оксалоацетат восстанавливается до малата и запасается в вакуолях с целью защиты ферментов цитозоля и пластид от низких значений pH, возникающих из-за диссоциации яблочной кислоты. В дневное время устьица закрыты, что предотвращает потерю воды, которая происходила бы из-за высоких дневных температур, и углерод, включенный ночью в малат, высвобождается NADP-зависимой малатдегидрогеназой в виде СО2. Этот СО2 далее вовлекается ферментом рубиско в цикл Кальвина. Так как этот метод фиксации СО2 впервые был открыт в многолетних толстянковых растениях из семейства Crassulaceae, он был назван crassulacean add metabolism (CAM-метаболизм), а соответствующие растения названы САМ-растениями.
Краткое содержание раздела 20.2 Фотодыхание, С4- и САМ-пути
■ Когда рубиско использует в качестве субстрата О2, а не СО2, образующийся 2-фосфогликолат распадается по кислород-зависимому пути. В результате увеличивается расход О2 — фотодыхание, или, более правильно, цикл фотосинтетического окисления углерода или С2-путь. 2-Фосфогликолат превращается в глиоксилат, затем в глицин, а после этого в серин под действием ферментов стромы хлоропластов, пероксисом и митохондрий.
■ У С4-растений путь ассимиляции углерода сводит к минимуму возможность фотодыхания: в клетках мезофилла СО2 включается в четырехуглеродное соединение, которое попадает в клетки обкладки и высвобождает СО2 в высоких концентрациях. Свободный СО2 фиксируется рубиско и другие реакции цикла Кальвина происходят так же, как и у С3-растений.
■ У CAM-растений в темноте СО2 соединяется с малатом и хранится в вакуолях до наступления дня, когда устьица закрыты (для уменьшения потери влаги), и малат становится источником СО2 для рубиско.
Биологическая библиотека - материалы для студентов, учителей, учеников и их родителей.
Наш сайт не претендует на авторство размещенных материалов. Мы только конвертируем в удобный формат материалы, которые находятся в открытом доступе и присланные нашими посетителями.
Если вы являетесь обладателем авторского права на любой размещенный у нас материал и намерены удалить его или получить ссылки на место коммерческого размещения материалов, обратитесь для согласования к администратору сайта.
Разрешается копировать материалы с обязательной гипертекстовой ссылкой на сайт, будьте благодарными мы затратили много усилий чтобы привести информацию в удобный вид.
Читайте также: