
Ресинтез атф в ходе гликолиза кратко
Обновлено: 05.07.2024
АТФ в процессе сокращения поставляет необходимую энергию для образования актомиозинового комплекса, а в процессе расслабления мышцы — обеспечивает энергией активный транспорт ионов кальция в ретикулум. Для поддержания сократительной функции мышцы концентрация АТФ в ней должна находиться на постоянном уровне от 2 до 5 ммоль/кг.
Поэтому при мышечной деятельности аденозинтрифосфорная кислота должна восстанавливаться с той же скоростью, с какой расщепляется в процессе сокращения, что осуществляется отдельными биохимическими механизмами ее ресинтеза.
Энергетические источники ресинтеза АТФ в скелетных мышцах и других тканях — богатые энергией фосфатсодержащие вещества. Они присутствуют в тканях (креатинфосфат, аденозиндифосфат) или образуются в процессе катаболизма гликогена, жирных кислот и других энергетических субстратов. Кроме того, в результате аэробного окисления различных веществ возникают энергии протонного градиента на мембране митохондрий.
Ресинтез аденозинтрифосфата может осуществляться в реакциях без участия кислорода (анаэробные механизмы) или с его участием (аэробный механизм). В обычных условиях ресинтез АТФ в мышцах происходит преимущественно аэробным путем. При напряженной физической работе, когда доставка кислорода к мышцам затруднена, включаются и анаэробные механизмы ресинтеза АТФ. В скелетных мышцах человека выявлены три вида анаэробных и один путь аэробного восстановления аденозинтрифосфата.
К анаэробным механизмам относятся креатинфосфокиназный (фосфогенный или алактатный), гликолитический (лактатный) и миокиназный механизмы.
Аэробный механизм ресинтеза АТФ заключается в окислительном фосфорилировании, протекающем в митохондриях, количество которых в скелетных мышцах при аэробных тренировках существенно увеличивается. Энергетическими субстратами аэробного окисления служат: глюкоза, жирные кислоты, частично аминокислоты, а также промежуточные метаболиты гликолиза (молочная кислота) и окисления жирных кислот (кетоновые тела).
Каждый механизм имеет разные энергетические возможности, которые оцениваются по следующим критериям: максимальная мощность, скорость развертывания, метаболическая емкость и эффективность.
Максимальная мощность — это наибольшая скорость образования АТФ в данном метаболическом процессе. Она лимитирует предельную интенсивность работы, выполняемой за счет используемого механизма.
Скорость развертывания — время достижения максимальной мощности данного пути ресинтеза адено-зинтрифосфата от начала работы.
Метаболическая емкость — общее количество АТФ, которое может быть получено в используемом механизме ресинтеза АТФ за счет величины запасов энергетических субстратов. Емкость лимитирует объем выполняемой работы. Метаболическая эффективность — это та часть энергии, которая накапливается в макроэргических связях аденозинт-рифосфата. Она определяет экономичность выполняемой работы и оценивается общим значением коэффициента полезного действия, представляющего отношение всей полезно затраченной энергии к ее общему количеству, выделенному при текущем метаболическом процессе.
Общий коэффициент полезного действия при преобразовании энергии метаболических процессов в механическую работу зависит от двух показателей:
- эффективности фосфорилирования;
- эффективности хемомеханического сопряжения (эффективности преобразования АТФ в механическую работу).
Эффективность хемомеханического сопряжения в процессах аэробного и анаэробного метаболизма примерно одинакова и составляет 50%.
Эффективность фосфорилирования наивысшая в алактатном анаэробном процессе — около 80%, и наименьшая в анаэробном гликолизе — в среднем 44%. В аэробном же процессе она составляет примерно 60%.
Таким образом, анаэробные механизмы имеют большую максимальную мощность и эффективность образования АТФ, но короткое время удержания и небольшую емкость, из-за малых запасов энергетических субстратов. Например, максимальная мощность креатинфосфокиназной реакции развивается уже на 0,5—0,7 с интенсивной работы и поддерживается 10-15 с у нетренированных людей идо 25-30 су высокотренированных спортсменов и составляет 3,8 кДж/кг в минуту.
Гликолитический механизм ресинтеза АТФ отличается невысокой эффективностью. Большая часть энергии остается в молекулах образующейся молочной кислоты. Концентрация последней находится в прямой зависимости от мощности и продолжительности работы, и может быть выделена только путем аэробного окисления.
Гликолиз — это основной путь энергообразования в упражнениях субмаксимальной мощности, предельная продолжительность которых составляет от 30 с до 2,5 мин (бег на средние дистанции, плавание на 100 и 200 м и др.).
Гликолитический механизм энергообразования служит биохимической основой специальной скоростной выносливости организма.
Миокиназная реакция происходит в мышцах при значительном увеличении концентрации АДФ в саркоплазме. Такая ситуация возникает при выраженном мышечном утомлении, когда другие пути ресинтеза уже не возможны.
Таким образом, анаэробные механизмы являются основными в энергообеспечении кратковременных упражнений высокой интенсивности.
При адаптации к интенсивным нагрузкам повышается активность ферментов анаэробных механизмов и запасов энергетических механизмов: содержание креатинфосфата в скелетных мышцах может увеличиваться в 1,5—2 раза, а содержание гликогена — почти в 3 раза.
Этот путь ресинтеза, так же как и креатинфосфатный, относится к анаэробным способам образования АТФ. Источником энергии, необходимой для ресинтеза АТФ, в данном случае является мышечный гликоген, концентрация которого в саркоплазме колеблется в пределах 0,2-3%. При анаэробном распаде гликогена от его молекулы под воздействием фермента фосфорилазы поочередно отщепляются концевые остатки глюкозы в форме глюкозо-1-фосфата. Далее молекулы глюкозо-1-фосфата через ряд последовательных стадий (их всего 10) превращаются в молочную кислоту (лактат), которая по своему химическому составу является как бы половинкой молекулы глюкозы. В процессе анаэробного распада гликогена до молочной кислоты, называемого гликолизом, образуются промежуточные продукты, содержащие фосфатную группу с макроэргической связью, которая легко переносится на АДФ с образованием АТФ.
Итоговое уравнение анаэробного расщепления гликогена (гликолиза) имеет следующий вид:
Все ферменты гликолиза находятся в саркоплазме мышечных клеток.
Гликолизу может также подвергаться глюкоза, поступающая в мыщцы из кровяного русла. Анаэробный распад глюкозы протекает по Уравнению:
Регуляция скорости гликолиза осуществляется путем изменения активности двух ферментов: фосфорилазы и фосфофруктокиназы. Фосфорилаза катализирует первую реакцию распада гликогена - отщепление от него глюкозо-1-фосфата. Этот фермент активируется адреналином, АМФ и ионами кальция, а ингибируется глюкозо-6-фосфатом и избытком АТФ. Второй регуляторный фермент гликолиза - фосфо- фруктокиназа - активируется АДФ и особенно АМФ, а тормозится избытком АТФ и лимонной кислотой (лимонная кислота - промежуточный метаболит цикла трикарбоновых кислот). Наличие таких регуля- торных механизмов приводит к тому, что в покое гликолиз протекает очень медленно, при интенсивной мышечной работе его скорость резко возрастает и может увеличиваться по сравнению с уровнем покоя почти в 2000 раз, причем повышение скорости гликолиза может наблюдаться уже в предстартовом состоянии за счет выделения адреналина.
Количественные критерии гликолитического пути ресинтеза АТФ:
Максимальная мощность - 750-850 кал/мин-кг, что примерно вдвое выше соответствующего показателя тканевого дыхания. Высокое значение максимальной мощности гликолиза объясняется содержанием в мышечных клетках большого запаса гликогена, наличием механизмов активации ключевых ферментов, приводящих к значительному росту скорости гликолиза (в 2000 раз!), отсутствием потребности в кислороде.
Время развертывания - 20-30 с. Это обусловлено тем, что все участники гликолиза (гликоген и ферменты) находятся в саркоплазме миоцитов, а также возможностью активации ферментов гликолиза. Как уже отмечалось, фосфорилаза - фермент, запускающий гликолиз, - активируется адреналином, который выделяется в кровь непосредственно перед началом работы. Ионы кальция, концентрация которых в саркоплазме повышается примерно в 1000 раз под воздействием двигательного нервного импульса, также являются мощными активаторами фосфорилазы.
Время работы с максимальной мощностью - 2-3 мин. Существуют две основные причины такой небольшой величины этого критерия. Во-первых, гликолиз протекает с высокой скоростью, что быстро приводит к уменьшению в мышцах концентрации гликогена и, следовательно, к последующему снижению скорости его распада. Во-вторых, в процессе гликолиза образуется молочная кислота (лактат), накопление которой приводит к повышению кислотности внутри мышечных клеток. В условиях повышенной кислотности снижается каталитическая активность ферментов, в том числе ферментов гликолиза, что также ведет к уменьшению скорости этого пути ресинтеза АТФ.
Гликолитический способ образования АТФ имеет ряд преимуществ перед аэробным путем. Он быстрее выходит на максимальную мощность (за 20-30 с, в то время как аэробный путь - за 3-4 мин), цме ет более высокую величину максимальной мощности (в 2 раза больше, чем у тканевого дыхания) и не требует участия митохондрий и кислорода.

Однако у этого пути есть и существенные недостатки. Этот процесс малоэкономичен. Распад до лактата одного остатка глюкозы, отщепленного от гликогена, дает только 3 молекулы АТФ, тогда как при аэробном окислении гликогена до воды и углекислого газа образуется 39 молекул АТФ в расчете на один остаток глюкозы. Такая неэкономичность в сочетании с большой скоростью быстро приводит к исчерпанию запасов гликогена.
Другой серьезный недостаток гликолитического пути ресинтеза АТФ - образование и накопление лактата, являющегося конечным продуктом этого процесса. Повышение концентрации лактата в мышечных волокнах вызывает сдвиг рН в кислую сторону, при этом происходят конформационные изменения мышечных белков, приводящие к снижению их функциональной активности. Таким образом, накопление молочной кислоты в мышечных клетках существенно нарушает их нормальное функционирование и ведет к развитию утомления.
При снижении интенсивности физической работы, а также в промежутках отдыха во время тренировки образовавшийся лактат может частично выходить из мышечных клеток в лимфу или кровь, что делает возможным повторное включение гликолиза.
Еще один метод оценки скорости гликолиза, фиксирующий последствия образования и накопления молочной кислоты, - это определение щелочного резерва крови. Щелочной резерв крови - это щелочные компоненты всех буферных систем крови. При поступлении во время мышечной работы в кровь молочной кислоты она вначале нейтрализуется путем взаимодействия с буферными системами крови (с их щелочными компонентами), и поэтому происходит снижение щелочного резерва крови.
Оценить вклад гликолиза в энергообеспечение выполненной физической работы можно также путем определения лактата в моче. В покое в моче лактат практически отсутствует. После тренировки, особенно с использованием интенсивных упражнений, с мочой выделяются большие количества молочной кислоты. При этом надо учесть, что в процессе тренировки гликолиз включается многократно и поэтому анализ мочи дает информацию о суммарном вкладе гликолитического пути ресинтеза в обеспечение энергией всех нагрузок, выполненных за время тренировки.
По величине лактатного кислородного долга можно судить о возможностях гликолитического пути ресинтеза АТФ. Так, величина лактатного долга свидетельствует о метаболической емкости гликолиза, а его максимальная мощность может быть оценена по отношению величины лактатного долга к времени выполнения предельной нагрузки субмаксимальной мощности.
В результате систематических тренировок с использованием субмаксимальных нагрузок в мышечных клетках повышается концентрация гликогена и увеличивается активность ферментов гликолиза. У высокотренированных спортсменов наблюдается развитие резистентности (нечувствительности) тканей и крови к снижению рН, и поэтому они сравнительно легко переносят сдвиг водородного показателя крови до 7 0 и ниже.
Аденилаткиназная (миокиназная) реакция
Аденилаткиназная (или миокиназная) реакция протекает в мышечных клетках в условиях значительного накопления в них АДФ, что обычно наблюдается при наступлении утомления. Аденилаткиназная реакция ускоряется ферментом аденилаткиназой (миокиназой), который находится в саркоплазме миоцитов. В ходе этой реакции одна молекула АДФ передает свою фосфатную группу на другую АДФ, в результате образуется АТФ и АМФ:
АДФ + АДФ ----------- — АТФ + АМФ
Длительное время этот путь образования АТФ рассматривался как аварийный механизм, обеспечивающий синтез АТФ в условиях, когда другие способы получения АТФ становятся неэффективными. Кроме того, считалось, что аденилаткиназная реакция ведет к уменьшению в миоцитах общего количества адениловых нуклеотидов, так как образующийся в этой реакции АМФ может дезаминироваться и превращаться в инозиновую кислоту:
АМФ ------- Инозиновая кислота + NH3
Однако в настоящее время этой реакции отводят не энергетическую, а регуляторную роль. Это связано с тем, что АМФ является мощным активатором ферментов распада углеводов - фосфорилазы и фосфо- фруктокиназы, участвующих как в анаэробном расщеплении гликогена и глюкозы до молочной кислоты, так и в их аэробном окислении до воды и углекислого газа. Оказалось также, что превращение АМФ в инозиновую кислоту имеет положительное значение для мышечной деятельности. Образующийся в результате дезаминирования аммиак может нейтрализовать молочную кислоту и тем самым предупреждать наступление изменений в миоцитах, связанных с ее накоплением (сдвиг РН, изменение конформации белков, снижение активности ферментов и ДР-)- При этом общее содержание адениловых нуклеотидов (АТФ, АДФ, АМФ) в клетках не изменяется, так как инозиновая кислота при взаимодействии с одной из аминокислот - аспарагиновой кислотой снова превращается в АМФ.
В табл. 7 приведены величины критериев описанных выше путей Ресинтеза АТФ.
СООТНОШЕНИЕ МЕЖДУ РАЗЛИЧНЫМИ ПУТЯМИ РЕСИНТЕЗА АТФ ПРИ МЫШЕЧНОЙ РАБОТЕ
При любой мышечной работе функционируют все три пути ресинтеза АТФ, но включаются они последовательно. В первые секунды работы ресинтез АТФ идет за счет креатинфосфатной реакции, затем включается гликолиз и, наконец, по мере продолжения работы на смену гликолизу приходит тканевое дыхание (рис. 19).
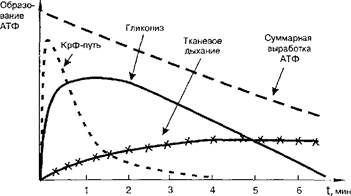 Рис. 19. Включение путей ресинтеза АТФ при выполнении физической работы |
Из рисунка видно, что переход энергообеспечения мышечной деятельности с анаэробных путей на аэробный ведет к уменьшению суммарной выработки АТФ за единицу времени, что находит отражение в снижении мощности выполняемой работы.
г
(гликолиз, лактатный)
Этот путь ресинтеза, так же как и креатинфосфатный, относится к анаэробным способам образования АТФ. Источником энергии, необходимой для ресинтеза АТФ, в данном случае является мышечный гликоген, концентрация которого в саркоплазме колеблется в пределах 0,2-3%. При анаэробном распаде гликогена от его молекулы под воздействием фермента фосфорилазы поочередно отщепляются концевые остатки глюкозы в форме глюкозо-1-фосфата. Далее молекулы глюкозо-1-фосфата через ряд последовательных стадий (их всего 10) превращаются в молочную кислоту (лактат), которая по своему химическому составу является как бы половинкой молекулы глюкозы. В процессе анаэробного распада гликогена до молочной кислоты, называемого гликолизом, образуются промежуточные продукты, содержащие фосфатную группу с макроэргической связью, которая легко переносится на АДФ с образованием АТФ.
Итоговое уравнение анаэробного расщепления гликогена (гликолиза) имеет следующий вид:
Все ферменты гликолиза находятся в саркоплазме мышечных клеток.
Гликолизу может также подвергаться глюкоза, поступающая в мыщцы из кровяного русла. Анаэробный распад глюкозы протекает по Уравнению:
Регуляция скорости гликолиза осуществляется путем изменения активности двух ферментов: фосфорилазы и фосфофруктокиназы. Фосфорилаза катализирует первую реакцию распада гликогена - отщепление от него глюкозо-1-фосфата. Этот фермент активируется адреналином, АМФ и ионами кальция, а ингибируется глюкозо-6-фосфатом и избытком АТФ. Второй регуляторный фермент гликолиза - фосфо- фруктокиназа - активируется АДФ и особенно АМФ, а тормозится избытком АТФ и лимонной кислотой (лимонная кислота - промежуточный метаболит цикла трикарбоновых кислот). Наличие таких регуля- торных механизмов приводит к тому, что в покое гликолиз протекает очень медленно, при интенсивной мышечной работе его скорость резко возрастает и может увеличиваться по сравнению с уровнем покоя почти в 2000 раз, причем повышение скорости гликолиза может наблюдаться уже в предстартовом состоянии за счет выделения адреналина.
Количественные критерии гликолитического пути ресинтеза АТФ:
Максимальная мощность - 750-850 кал/мин-кг, что примерно вдвое выше соответствующего показателя тканевого дыхания. Высокое значение максимальной мощности гликолиза объясняется содержанием в мышечных клетках большого запаса гликогена, наличием механизмов активации ключевых ферментов, приводящих к значительному росту скорости гликолиза (в 2000 раз!), отсутствием потребности в кислороде.
Время развертывания - 20-30 с. Это обусловлено тем, что все участники гликолиза (гликоген и ферменты) находятся в саркоплазме миоцитов, а также возможностью активации ферментов гликолиза. Как уже отмечалось, фосфорилаза - фермент, запускающий гликолиз, - активируется адреналином, который выделяется в кровь непосредственно перед началом работы. Ионы кальция, концентрация которых в саркоплазме повышается примерно в 1000 раз под воздействием двигательного нервного импульса, также являются мощными активаторами фосфорилазы.
Время работы с максимальной мощностью - 2-3 мин. Существуют две основные причины такой небольшой величины этого критерия. Во-первых, гликолиз протекает с высокой скоростью, что быстро приводит к уменьшению в мышцах концентрации гликогена и, следовательно, к последующему снижению скорости его распада. Во-вторых, в процессе гликолиза образуется молочная кислота (лактат), накопление которой приводит к повышению кислотности внутри мышечных клеток. В условиях повышенной кислотности снижается каталитическая активность ферментов, в том числе ферментов гликолиза, что также ведет к уменьшению скорости этого пути ресинтеза АТФ.
Гликолитический способ образования АТФ имеет ряд преимуществ перед аэробным путем. Он быстрее выходит на максимальную мощность (за 20-30 с, в то время как аэробный путь - за 3-4 мин), цме ет более высокую величину максимальной мощности (в 2 раза больше, чем у тканевого дыхания) и не требует участия митохондрий и кислорода.
Однако у этого пути есть и существенные недостатки. Этот процесс малоэкономичен. Распад до лактата одного остатка глюкозы, отщепленного от гликогена, дает только 3 молекулы АТФ, тогда как при аэробном окислении гликогена до воды и углекислого газа образуется 39 молекул АТФ в расчете на один остаток глюкозы. Такая неэкономичность в сочетании с большой скоростью быстро приводит к исчерпанию запасов гликогена.
Другой серьезный недостаток гликолитического пути ресинтеза АТФ - образование и накопление лактата, являющегося конечным продуктом этого процесса. Повышение концентрации лактата в мышечных волокнах вызывает сдвиг рН в кислую сторону, при этом происходят конформационные изменения мышечных белков, приводящие к снижению их функциональной активности. Таким образом, накопление молочной кислоты в мышечных клетках существенно нарушает их нормальное функционирование и ведет к развитию утомления.
При снижении интенсивности физической работы, а также в промежутках отдыха во время тренировки образовавшийся лактат может частично выходить из мышечных клеток в лимфу или кровь, что делает возможным повторное включение гликолиза.
Еще один метод оценки скорости гликолиза, фиксирующий последствия образования и накопления молочной кислоты, - это определение щелочного резерва крови. Щелочной резерв крови - это щелочные компоненты всех буферных систем крови. При поступлении во время мышечной работы в кровь молочной кислоты она вначале нейтрализуется путем взаимодействия с буферными системами крови (с их щелочными компонентами), и поэтому происходит снижение щелочного резерва крови.
Оценить вклад гликолиза в энергообеспечение выполненной физической работы можно также путем определения лактата в моче. В покое в моче лактат практически отсутствует. После тренировки, особенно с использованием интенсивных упражнений, с мочой выделяются большие количества молочной кислоты. При этом надо учесть, что в процессе тренировки гликолиз включается многократно и поэтому анализ мочи дает информацию о суммарном вкладе гликолитического пути ресинтеза в обеспечение энергией всех нагрузок, выполненных за время тренировки.
По величине лактатного кислородного долга можно судить о возможностях гликолитического пути ресинтеза АТФ. Так, величина лактатного долга свидетельствует о метаболической емкости гликолиза, а его максимальная мощность может быть оценена по отношению величины лактатного долга к времени выполнения предельной нагрузки субмаксимальной мощности.
В результате систематических тренировок с использованием субмаксимальных нагрузок в мышечных клетках повышается концентрация гликогена и увеличивается активность ферментов гликолиза. У высокотренированных спортсменов наблюдается развитие резистентности (нечувствительности) тканей и крови к снижению рН, и поэтому они сравнительно легко переносят сдвиг водородного показателя крови до 7 0 и ниже.
Аденилаткиназная (миокиназная) реакция
Аденилаткиназная (или миокиназная) реакция протекает в мышечных клетках в условиях значительного накопления в них АДФ, что обычно наблюдается при наступлении утомления. Аденилаткиназная реакция ускоряется ферментом аденилаткиназой (миокиназой), который находится в саркоплазме миоцитов. В ходе этой реакции одна молекула АДФ передает свою фосфатную группу на другую АДФ, в результате образуется АТФ и АМФ:
АДФ + АДФ ----------- — АТФ + АМФ
Длительное время этот путь образования АТФ рассматривался как аварийный механизм, обеспечивающий синтез АТФ в условиях, когда другие способы получения АТФ становятся неэффективными. Кроме того, считалось, что аденилаткиназная реакция ведет к уменьшению в миоцитах общего количества адениловых нуклеотидов, так как образующийся в этой реакции АМФ может дезаминироваться и превращаться в инозиновую кислоту:
АМФ ------- Инозиновая кислота + NH3
Однако в настоящее время этой реакции отводят не энергетическую, а регуляторную роль. Это связано с тем, что АМФ является мощным активатором ферментов распада углеводов - фосфорилазы и фосфо- фруктокиназы, участвующих как в анаэробном расщеплении гликогена и глюкозы до молочной кислоты, так и в их аэробном окислении до воды и углекислого газа. Оказалось также, что превращение АМФ в инозиновую кислоту имеет положительное значение для мышечной деятельности. Образующийся в результате дезаминирования аммиак может нейтрализовать молочную кислоту и тем самым предупреждать наступление изменений в миоцитах, связанных с ее накоплением (сдвиг РН, изменение конформации белков, снижение активности ферментов и ДР-)- При этом общее содержание адениловых нуклеотидов (АТФ, АДФ, АМФ) в клетках не изменяется, так как инозиновая кислота при взаимодействии с одной из аминокислот - аспарагиновой кислотой снова превращается в АМФ.
В табл. 7 приведены величины критериев описанных выше путей Ресинтеза АТФ.
СООТНОШЕНИЕ МЕЖДУ РАЗЛИЧНЫМИ ПУТЯМИ РЕСИНТЕЗА АТФ ПРИ МЫШЕЧНОЙ РАБОТЕ
При любой мышечной работе функционируют все три пути ресинтеза АТФ, но включаются они последовательно. В первые секунды работы ресинтез АТФ идет за счет креатинфосфатной реакции, затем включается гликолиз и, наконец, по мере продолжения работы на смену гликолизу приходит тканевое дыхание (рис. 19).
| Рис. 19. Включение путей ресинтеза АТФ при выполнении физической работы |
Из рисунка видно, что переход энергообеспечения мышечной деятельности с анаэробных путей на аэробный ведет к уменьшению суммарной выработки АТФ за единицу времени, что находит отражение в снижении мощности выполняемой работы.
Другим важнейшим анаэробным механизмом ресинтеза АТФ является гликолиз. Гликолизом называют начальный этап расщепления углеводов (гликогена и глюкозы), завершающийся образованием пировиноградной или молочной кислоты. При адекватном потребностям снабжении организма кислородом пировиноградная кислота подвергается дальнейшим превращениям с его участием, приводящим к образованию конечных продуктов обмена: СО2 и Н2О. В условиях напряженной мышечной деятельности, когда имеет место неадекватное потребностям снабжение организма кислородом, могут значительно усиливаться анаэробные превращения углеводов, завершающиеся образованием молочной кислоты. В силу этого анаэробный гликолиз, приводящий к накоплению молочной кислоты, можно рассматривать как самостоятельный процесс ресинтеза АТФ.
В цепи реакций гликолиза есть две окислительные стадии, сопряженные с накоплением энергии в молекулах АТФ. Связанное с этими стадиями образование АТФ не только покрывает расход энергии (в виде АТФ) на начальных этапах гликолиза, но и образует дополнительное количество АТФ, используемое на выполнение работы.
Энергетическая эффективность гликолиза невысока. Расщепление 1 моля глюкозы до молочной кислоты сопровождается ресинтезом 2 молей АТФ. Для сравнения укажем, что расщепление 1 моля глюкозы в аэробных превращениях до образования конечных продуктов обмена (СО2 и Н2О) обеспечивает ресинтез 38 молей АТФ.
Анаэробное расщепление гликогена характеризуется большей энергетической эффективностью, чем расщепление глюкозы. Выход АТФ в этом случае составляет 3 моля АТФ на моль глюкозных остатков гликогена. Более высокая энергетическая эффективность анаэробного расщепления гликогена связана с тем, что он является более энергоемким веществом, чем глюкоза, и начальное его фосфорилирование осуществляется в реакции со свободной фосфорной кислотой, тогда как для начального фосфорилирования глюкозы требуются затраты АТФ.
Ферментные системы гликолиза локализованы на мембранах цитоплазматической сети (в мышечных волокнах – на мембранах саркоплазматического ретикулума). Сам же процесс гликолиза протекает в цитоплазме (саркоплазме) мышечных клеток.
Гликолиз уступает креатинфосфокиназной реакции по скорости развертывания и по развиваемой максимальной мощности. Скорость развертывания гликолиза связана с интенсивностью выполняемого упражнения и зависит от тренированности спортсмена. У тренированных спортсменов гликолиз достигает максимальной мощности через 20-40 сек после начала интенсивного упражнения.
Максимальная мощность гликолиза приблизительно в 1,5 раза ниже мощности креатинфосфокиназной реакции, но в 1,5-3 раза выше мощности аэробного процесса. У тренированных спортсменов она может достигать 750 кал/(кг.мин).
Скорость развертывания и максимальная мощность гликолиза в первую очередь определяется количеством и активностью его ключевого фермента – фосфофруктокиназы. Под влиянием систематической тренировки, связанной с использованием упражнений, в энергообеспечении которых участвует гликолиз, оба эти параметра (мощность и скорость развертывания) претерпевают значительные изменения. То есть, происходит более быстрое развертывание и повышается мощность гликолиза.
Гликолиз имеет значительное превосходство перед креатинфосфатной реакцией по метаболической емкости. Прямые измерения метаболической емкости гликолиза невозможны, т.к. гликолиз обеспечивает энергией работу только в комплексе с другими энергопреобразующими процессами: креатинфосфокиназной реакцией и аэробным окислением. По расчетам итальянского исследователя Р. Маргариа у тренированного спортсмена гликолиз мог бы обеспечить работу субмаксимальной (ниже максимальной) интенсивности в течении 40 сек при условии выключения из работы других процессов, обеспечивающих энергией мышечную деятельность.
Косвенные данные, основанные на определении общего количества образованной при работе молочной кислоты, свидетельствуют о том, что максимальная емкость гликолиза в 5-7 раз превышает емкость креатинфосфатного механизма.
Емкость гликолиза определяется целым рядом свойств организма, среди которых первостепенное значение имеют запасы гликогена в мышцах, активность ферментов гликолиза, компенсаторные возможности организма и волевые качества, обеспечивающие способность продолжать работу в условиях очень быстро нарастающих изменений во внутренней среде организма. Высокая значимость компенсаторных возможностей для протекания гликолиза при работе связана с образованием молочной кислоты. Молочная кислота оказывает сильное воздействие на организм. Накапливаясь в значительных количествах в мышечных волокнах, она вызывает повышение осмотического давления, в результате чего волокна набухают, сдавливая нервные окончания. Набухшие мышечные волокна теряют эластичность, в них происходит рассогласование взаимного расположения внутриклеточных структурных компонентов. В результате сдавливания нервных окончаний (нервно-мышечных синапсов) могут возникнуть болевые ощущения, ощущения тяжести.
Еще более существенное влияние на работоспособность оказывает вызываемый молочной кислотой сдвиг реакции внутренней среды в кислую сторону, что воздействует на белки, в частности, сократительные, ухудшая их сократительную способность. Под влиянием изменений реакции внутренней среды снижается активность многих ферментов, в частности ферментов, катализирующих реакции энергетического обмена. В результате замедляются процессы ресинтеза АТФ, что приводит к снижению работоспособности.
Компенсаторные возможности организма реализуются через нейтрализацию части образованной молочной кислоты буферными системами (бикарбонатной, белковой), а также через повышенную устойчивость ферментов к сдвигам реакции внутренней среды в кислую сторону, вызываемым молочной кислотой. Под влиянием систематической тренировки с использованием упражнений, приводящих к накоплению молочной кислоты, повышаются возможности указанных компонентов компенсаторной системы. Повышается буферная емкость (способность нейтрализовать молочную и другие кислоты) и устойчивость ферментов к изменениям рН. Последнее связано с тем, что каждый фермент в организме существует в нескольких модификациях, проявляющих свою наивысшую активность в несколько разных условиях. Систематическое накопление молочной кислоты под влиянием выполнения соответствующих упражнений, вызывает усиленный синтез модификаций ферментов, более устойчивых к изменениям рН.
Влияние молочной кислоты на обменные процессы проявляется не только на уровне мышечной ткани. Обладая высокой диффузионной способностью, молочная кислота выходит в кровь. Это снижает ее концентрацию в мышечной ткани и степень воздействия на мышечную ткань, а также способствует устранению части молочной кислоты даже по ходу выполнения работы. Так, интенсивно работающее в этот период сердце извлекает часть молочной кислоты из крови и использует ее в качестве источника энергии. Могут использовать молочную кислоту в качестве источника энергии мышцы, работающие с невысокой интенсивностью.
С другой стороны, вышедшая в кровь молочная кислота оказывает воздействие на некоторые органы и ткани. Так, влияя на рецепторы, расположенные в кровеносных сосудах, она вызывает чрезмерное усиление внешнего дыхания, что отвлекает часть поступающего в организм кислорода на обеспечение энергией работы дыхательных мышц.
Гликолиз обладает невысокой энергетической эффективностью, так как большая часть энергии, заключенной в химических связях углеводов, не освобождается в ходе гликолиза, а сохраняется в молочной кислоте. Если при полном окислении (до СО2 и Н2О) одного моля глюкозы (180г) освобождается 2880 кДж энергии, то в ходе гликолиза только 196 кДж. Из этого количества в молекулах АТФ аккумулируется около 61 кДж (приблизительно 31%). Из-за невысокой эффективности участие гликолиза в энергетической обеспечении работы приводит к быстрому истощению запасов гликогена в работающих мышцах.
У нетренированных лиц мощность работы, соответствующая уровню ПАНО, невысока. Она вызывает повышение частоты сердечных сокращений приблизительно до 130 уд/мин. У тренированных спортсменов ПАНО проявляет себя при более высокой мощности упражнения. Частота сердечных сокращений при этом может доходить до 160-170 уд/мин и более.
Гликолиз участвует в энергетическом обеспечении как циклических, так и ациклических видов спорта, в частности, спортивных единоборств, некоторых спортивных игр. Особенно велика его роль в тех видах спорта, где высокая интенсивность работы сочетается со статическим режимом деятельности мышц: вольная и греко-римская борьба, хоккей с шайбой, некоторые гимнастические упражнения и др.
Следует, однако, подчеркнуть, что фосфофруктокиназа менее чувствительна к изменениям отношения [АТФ] / [АДФ], чем ферменты аэробной фазы превращений углеводов. Резкое увеличение ее активности (и, следовательно, скорости анаэробного этапа превращений углеводов) происходит только в условиях интенсивной работы, когда аэробные реакции не справляются с ресинтезом АТФ и происходит значительное снижение отношения [АТФ] / [АДФ].
Другим важным регулятором скорости гликолиза является фермент фосфорилаза. Ведущая роль в активации этого фермента принадлежит гормонам адреналину и глюкагону. Активация фосфорилазы может осуществляться уже в предстартовом состоянии, характеризующимся высокой гормональной активностью.
Дано определение ресинтеза АТФ. Описаны основные пути ресинтеза АТФ в мышечных волокнах: креатинфосфатный, гликолитический, миокиназный и тканевое дыхание. Описаны количественные критерии путей ресинтеза АТФ, соотношение между различными путями ресинтеза АТФ при мышечной работе, а также между путями ресинтеза АТФ и зонами относительной мощности.

Ресинтез АТФ в мышечных волокнах
Определение
Ресинтез АТФ – синтез АТФ из различных энергетических субстратов во время физической работы в мышечных волокнах.
Формула ресинтеза АТФ выглядит следующим образом:
Пути ресинтеза АТФ
Ресинтез АТФ может осуществляться двумя путями:
- с участием кислорода (аэробный путь).
- без участия кислорода (анаэробный путь);
Аэробный путь (тканевое дыхание, аэробное или окислительное фосфорилирование) – основной способ образования АТФ в мышечных волокнах. Он протекает в митохондриях мышечных волокон. В результате тканевого дыхания выделяется 39 молекул АТФ. Окисляемое вещество распадается до углекислого газа и воды.
Анаэробный ресинтез АТФ
Анаэробный ресинтез АТФ в мышечных волокнах возможен посредством нескольких механизмов:
-
– ресинтез АТФ из креатинфосфата; – ресинтез АТФ из гликогена мышц;
- Миокиназный (аденилаткиназный) ресинтез АТФ – ресинтез АТФ из АДФ при значительном накоплении в мышечных волокнах АДФ. Рассматривается как аварийный механизм, обеспечивающий ресинтез АТФ, когда другие пути ресинтеза АТФ невозможны.
Количественные критерии путей ресинтеза АТФ
Существуют количественные критерии путей ресинтеза АТФ. К ним можно отнести: максимальную мощность, время развертывания, время сохранения или поддержания максимальной мощности, метаболическую ёмкость (табл. 1).
- Максимальная мощность – максимальное количество АТФ, которое может образоваться в единицу времени при функционировании данного пути ресинтеза АТФ.
- Время развертывания – минимальная длительность, необходимая для выхода ресинтеза АТФ на свою максимальную мощность.
- Время сохранения или поддержания максимальной скорости – длительность функционирования данного пути ресинтеза АТФ с максимальной мощностью.
- Метаболическая ёмкость – количество АТФ, которое может образоваться во время мышечной работы за счёт данного пути ресинтеза АТФ.
Таблица 1. Количественные критерии основных путей ресинтеза АТФ (С.С. Михайлов, 2009)
Соотношение между различными путями ресинтеза АТФ
При любой мышечной работе функционируют все три основных механизма ресинтеза АТФ, но включаются они последовательно. В первые секунды ресинтез АТФ осуществляется за счет креатинфосфатной реакции, затем включается гликолиз. По мере продолжения работы на смену гликолизу приходит тканевое дыхание (рис.1). Эта смена механизмов ресинтеза АТФ приводит к уменьшению суммарной выработки АТФ.
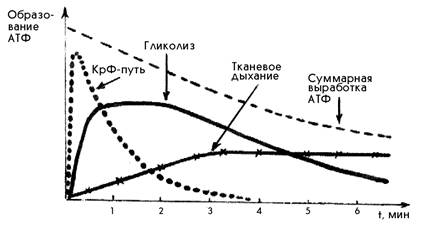
Рис.1. Включение путей ресинтеза АТФ при выполнении физической работы (С.С. Михайлов, 2009)
Пути ресинтеза АТФ и зоны относительной мощности
В.С. Фарфель приводит следующее соотношение мощности работы и основной системы энергообеспечения (табл.2)
Таблица 2. Зоны мощности работы и основная система энергообеспечения (В.С. Фарфель)
| Мощность работы | Основная система энергообеспечения | Типичное время работы |
| Максимальная | Креатинфосфатная реакция | до 20 с |
| Субмаксимальная | Гликолиз | до 5 мин. |
| Большая | Гликолиз+ тканевое дыхание | до 30 мин. |
| Умеренная | Тканевое дыхание | Более 30 мин. |
J.T. Cramer (2008) приводит несколько иное соотношение зон мощности и основных систем энергообеспечения (табл.3)
Таблица 3. — Зоны относительной мощности и основная ситема энергообеспечения (J.T. Cramer, 2008)
| % от максимальной мощности работы | Основная система энергообеспечения | Время работы |
| 90-100 | Креатинфосфатная реакция | 5-10 с |
| 75-90 | Гликолиз | 15-30 с |
| 30-75 | Гликолиз+ тканевое дыхание | 1-3 мин. |
| 20-30 | Тканевое дыхание | Более 3 мин. |
Литература
- Михайлов С.С. Спортивная биохимия. – М.: Советский спорт, 2009.– 348 с.
- Волков Н.И., Несен Э.Н., Осипенко А.А., Корсун С.Н. Биохимия мышечной деятельности.- Киев: Олимпийская литература, 2000.- 504 с.
Похожие записи:
Тест времени реакции на сигнал
Представлена программа расчета времени реакции на сигнал, предназначенная для использования в учебных целях, например на занятиях по…
Саркоплазматическая гипертрофия мышц
Дано определение и описаны механизмы саркоплазматической гипертрофии скелетных мышц. Показано, что этот вид гипертрофии мышц широко…
Классификация типов конституции человека М.В. Черноруцкого
Рассмотрена классификация типов конституции человека, разработанная выдающимся терапевтом М.В. Черноруцким в 1925 году. Классификация типов конституции человека М.В.
Типы гипертрофии скелетных мышц человека
В статье дается классификация различных видов гипертрофии скелетных мышц человека на основе ряда классификационных признаков: времени проявления…
Миомейкер: Мембранный активатор слияния миобластов и образования мышц
Ученые установили, что для образования мышечных волокон необходимо слияние клеток-предшественников, которые называются миобластами. Эти клетки имеют только…
Как только в процессе анаэробной мышечной работы креатинфосфокиназный механизм перестает обеспечивать необходимую скорость восстановления АТФ в мышцах, в энергообеспечение работы вовлекается анаэробный гликолитический механизм ресинтеза АТФ. В процессе гликолиза используются в основном внутримышечные запасы гликогена, а также глюкоза, поступающая из крови. Они постепенно расщепляются до молочной кислоты с участием многих ферментов (рис. 124).
Большинство ферментов гликолиза локализовано в саркоплазме мышечных волокон. Ферменты фосфорилаза и гексокиназа, обеспечивающие начальные реакции гликолиза, легко активируются при повышении концентрации АДФ и неорганического фосфата в саркоплазме. Кроме того, образование активной формы фосфорилазы стимулируется катехоламинами и ионами Са 2+ , уровень которых повышается при мышечном сокращении. Все это способствует быстрому подключению гликолиза к ресинтезу АТФ уже с первых секунд работы, о чем свидетельствует повышение концентрации молочной кислоты в мышцах.
 |
Активации гидролиза способствует также снижение концентрации креатинфосфата в мышцах и накопление АМФ, образующегося в миокиназной реакции ресинтеза АТФ.
Обобщенно процесс гликолиза и гликогенолиза может быть представлен в виде следующих уравнений:
Глюкоза Молочная кислота
Гликоген Молочная кислота
Энергетический баланс гликолиза в случае, когда исходным веществом служит глюкоза, составляет 2 моля АТФ на 1 моль расщепляемых углеводов, а гликогенолиза, когда исходным веществом является гликоген мышц, — 3 моля АТФ на 1 моль расщепляемого глюкозного эквивалента.
Максимальная мощностьгликолиза у хорошо тренированных спортсменов может составлять 3,1 кДж • кг -1 • мин -1 , а у нетренированных людей — 2,5 кДж • кг -1 • мин -1 . Это несколько ниже, чем мощность креатинфосфокиназной реакции, но в 2—3 раза выше мощности аэробного процесса.
На максимальную мощность этот механизм выходит уже на 20—30-й секунде после начала работы, т. е. его скорость развертывания значительно меньше, чем креатинфосфокиназного. К концу 1 -и минуты работы гликолиз становится основным механизмом ресинтеза АТФ. Однако при дальнейшей работе наблюдается снижение активности ключевых ферментов гликолиза под влиянием образующейся молочной кислоты или снижения внутриклеточного рН, что приводит к снижению скорости ресинтеза АТФ в этом механизме.
Метаболическая емкостьгликолиза, определяемая внутримышечными запасами углеводов и зависящая от резервов буферных систем, стабилизирующих величину внутриклеточного рН, обеспечивает поддержание анаэробной работы продолжительностью от 30 с до 2—6 мин.
Общее количество энергии, которое образуется в гликолитическом механизме у нетренированных людей, не превышает 840 Дж • кг -1 , что соответствует концентрации молочной кислоты в крови около 13 ммоль • л -1 , которая для них является граничной. У спортсменов, которые в процессе тренировки используют физические нагрузки анаэробной гликолитической направленности, лактатная емкость составляет 1760—2090 Дж • кг -1 , что соответствует наличию молочной кислоты в крови в пределах 25— 30 ммоль • л -1 . Тем не менее у спортсменов, специализирующихся в видах спорта на выносливость, после выполненной мышечной работы концентрация молочной кислоты в крови не превышает 10—13 ммоль • л -1 и зависит от содержания быстросокращающихся волокон в скелетных мышцах.
Гликолитический механизм ресинтеза АТФ отличается невысокой эффективностью, так как в процессе анаэробного распада 1 моля глюкозы из 2880 кДж энергии, содержащейся в ней, высвобождается только 240 кДж. Большая часть энергии остается в молекулах образующейся молочной кислоты и может быть выделена только путем аэробного окисления.
Из общего количества энергии, выделенной в ходе гликолиза, в доступную для использования форму макроэргических фосфатных связей АТФ преобразуется от 80 до 125 кДж, а остальная энергия выделяется в виде тепла. Исходя из этого метаболическая эффективность гликолиза оценивается значениями КПД порядка 0,35—0,52. Это означает, что примерно половина всей выделяемой энергии превращается в тепло и не может быть использована в работе. В результате при гликолитической работе повышается скорость теплопродукции в работающих мышцах и их температура до 41-42 • С.
Гликолиз - основной путь энергообразования в упражнениях субмаксимальной мощности, предельная продолжительность которых составляет от 30 с до 2,5 мин (бег на средние дистанции, плавание на 100 и 200 м, велосипедные гонки на треке и т. п.).
За счет гликолиза совершаются длительные ускорения по ходу упражнения и на финише дистанции.
Гликолитический механизм энергообразования является биохимической основой специальной скоростной выносливости организма.
Гликолитический путь ресинтеза АТФ сопровождается накоплением молочной кислоты в мышцах, концентрация которой находится в прямой зависимости от мощности и общей продолжительности упражнения.
Молочная кислота в водной среде подвергается диссоциации на ионы и вызывает изменение концентрации водородных ионов и величины рН внутриклеточной среды:
СНз-СНОН-СООН ← → СНз-СНОН-СОо - + Н +
Молочная кислота Ион молочной кислоты (лактат)
Умеренный сдвиг рН в кислую сторону активирует работу ферментов дыхательного цикла в митохондриях и усиливает аэробное энергообразование. Значительное изменение рН среды в мышцах от 7,1 в состоянии покоя до 6,5 при изнеможении угнетает ферменты, регулирующие скорость гликолиза и сокращение мышц. При значении внутримышечного рН 6,4 прекращается расщепление гликогена, что вызывает резкое снижение уровня АТФ и развитие утомления.
Увеличение количества молочной кислоты в саркоплазме мышц сопровождается изменением осмотического давления. При этом вода из межклеточной среды поступает внутрь мышечных волокон, вызывая их набухание, что может привести к сдавливанию нервных окончаний и возникновению болевых ощущений в мышцах.
Молочная кислота может диффундировать через клеточные мембраны по градиенту концентрации и поступать из работающих мышц в кровь. Обычно максимальное накопление молочной кислоты в крови наблюдается через 5—7 мин после работы. Молочная кислота взаимодействует с бикарбонатной буферной системой крови, что приводит к образованию "неметаболического" избытка СО2:
СНз-СНОН-СООН + NаНСОз → СН3-СНОН-СООNа + Н + + НСО3 -
Увеличение концентрации водородных ионов и повышение напряжения С02 в крови способствуют активации дыхательного центра, поэтому при выходе молочной кислоты в кровь резко усиливается легочная вентиляция и поставка кислорода к работающим мышцам. Значительное накопление молочной кислоты, появление избыточного СО2, изменение рН и гипервентиляция легких, отражающие усиление гликолиза в мышцах, обнаруживается при увеличении интенсивности выполняемого упражнения более 50 % максимальной аэробной мощности. Этот уровень нагрузки обозначается как порог анаэробного обмена (ПАНО), или порог лактата (ПЛ). Чем раньше он будет достигнут, тем быстрее вступит в действие гликолиз, сопровождающийся накоплением молочной кислоты и последующим развитием утомления работающих мышц.
Величина ПАНО является важным показателем эффективности процессов энергообразования в мышцах, интенсивности тренирующих нагрузок, роста степени тренированности, который широко используется при биохимическом контроле функционального состояния спортсмена. Наиболее обоснованным является метод построения индивидуальных кривых зависимости концентрации лактата от мощности выполняемой физической работы, которую выражают либо в ваттах (Вт), либо в процентах от МПК. Резкий излом кривой свидетельствует о переходе мышц в анаэробный режим работы, после чего концентрация молочной кислоты с увеличением мощности нагрузки будет увеличиваться за счет повышения интенсивности гликолиза.
У многих спортсменов ПАНО соответствует уровню молочной кислоты, равному 2—4 ммоль • л -1 . С ростом степени тренированности на выносливость порог лактата увеличивается, т. е. наступает при более интенсивной работе.
Конфликтные ситуации в медицинской практике: Наиболее ярким примером конфликта врача и пациента является.
Читайте также: