
Полигибридное скрещивание это в биологии кратко
Обновлено: 02.07.2024
В этой статье читатель ознакомиться с информацией о том, что являет собой процесс скрещивания, селекция, а также будут рассмотрены их различные формы, особенности протекания, гибридное и дигибридное скрещивание и значение селекции в природе и жизни человека.
Что такое скрещивание
Скрещивание – это вариант из списка методов селекции представителей животного и растительного мира. Процесс искусственного или естественного объединения генотипов, различающихся между собой наследственной информацией, которую они несут. Происходит скрещивания при оплодотворении.
Существует довольно обильное количество способов скрещивания живых организмов, и если говорить о растениях, то выделяют следующие:
- Скрещивание, носящее внутривидовой характер, то есть протекание передачи признаков между особями одного вида.
- Диаллельное скрещивание – при котором линии испытуемых или даже целые сорта скрещиваются во всевозможных комбинационных вариантах.
- Насыщающее скрещивание – это процесс многократного повторения скрещивания гибридов имеющих исходную форму предка с целью возврата признака.
- Топкросс и беккросс отличающиеся друг от друга тем, что первый тип используется для скрещивания одних сортов с другими, а второй процесс протекает между поколения F1.
- Инконгруентное скрещивание – как правило, межвидовое или межродовое скрещивание организмов с несоответствующим хромосомным числом или различие в их наборе.
- Конгруэнтное скрещивание, характерное для организмов с совместным хромосомным набором.
- Ступенчатое скрещивание – представляет собой процесс последовательного использования определенного количества родительских форм.
- Простое скрещивание.

Дигибридный способ скрещивания
Дигибридное скрещивание – это процесс, при котором происходит скрещивание организмов, имеющих различия по альтернативным признакам в двух парах, например, форма цветка и его окрас. По третьему закону Грегора Менделя при скрещивании такого типа различные гены аллельных пар, находящиеся в разных гомологичных хромосомах, наследуют свои признаки в независимом порядке один от другого.

Представители царств Растений, Животных, Грибов и Микроорганизмов носят различия в своем генетическом материале, и впоследствии они имеют разнообразные признаки. Процесс анализа наследования пары признаков базируется на изучении наследственности, которую несет каждая пара признака, при этом не стоит обращать внимание на иные пары. После отслеживания наследуемых качеств организма остается только сравнить и объединить результаты наблюдений.
Дигибридное скрещивание, как и полигибридное, зависит от расположения гена, а именно важно положение информации в одной или разных хромосомах. Прекрасно иллюстрирует процесс дигибридного скрещивания эксперимент Г. Менделя, в котором он использовал гомозиготные гороховые растения, носящие отличие между собой одновременно в двух признаках. Первое различие заключалось в желтом и зеленом цвете плодов, а второе - в их поверхностной характеристике: желтый горох был гладкий, а зеленый - морщинистый.
Гибриды всего первого поколения получились желтыми и гладкими, а это указывало на доминирующую способность данного признака. Из всего этого можно сделать вывод, что родительские организмы носили генотип AABB (доминантный) и aabb (рецессивный), а первое гибридное поколение стало гетерозиготным: AaBb. Второе поколение гибридов получало всевозможные вариации наследования признаков с расщеплением 9:3:3:1. Больше всего было гладких и желтых, а меньше всего - морщинистых и зеленых гороховых семян.
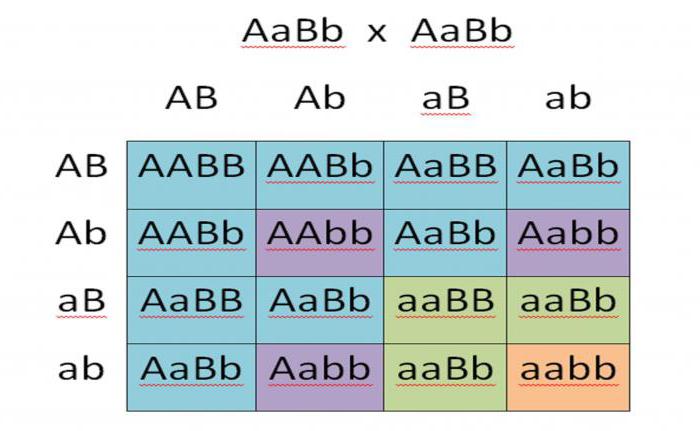
Скрещивание дигибридным образом носит значимый селекционный характер и является очень важным явлением в природе и человеческой деятельности.
Полигибридный способ скрещивания
Полигибридное скрещивание растений представляет собой процесс скрещивания форм, которые отличаются одна от другой одновременно по трем и более парам определенных признаков. В данном способе скрещивания гетерозиготные особи по n признаку могут создать гаметы 2 n типа. F2 может образовать потомство с 3 n генотипом.
Определив частоту воспроизведения конкретного генотипа в родительском потомстве, различающегося каким-либо числом генов, которые наследуются независимо, можно просчитать шанс проявления конкретного генотипа всех генных пар по отдельности, после чего полученные варианты перемножаются.
Например, вам необходимо сделать расчет на частоту AabbCc генотипа при скрещивании с AaBbcc. Можно произвести расчет либо по решетке Пеннета, либо обычным умножением, а именно Aa*Aa=1/2, bb*Bb=1/4 и Cc*Cc=1/2, и как следствие получаем вероятность проявления представителей вида с AabbCc генотипом 1/2*1/4*1/2=1/16.
Селекция
Селекция – это отдел науки, изучающий методы, которые позволят создать новые или улучшенные уже существующие виды живых организмов. Скрещивание растений, животных, выведение штаммов среди микроорганизмов – всем этим занимается именно данная наука. Она использует множество различных способов скрещивания, например, дигибридное скрещивание или полигибридное.

Необходимость селекции
Значение селекции как растений, так и животных заключается в том, что это явление позволяет создавать различные новые формы живых существ, имеющих более устойчивые способности к различным негативным воздействиям, например, лучше переносить перепады низких и высоких температур, будут хуже подвергаться нападкам паразитических насекомых. Растения могут давать гораздо больше урожая и в более короткие сроки, лучше бороться с болезнями и быть устойчивыми к ним. Среди животных можно выводить формы, имеющие конкретные необходимые признаки, и избавляться от дефектов породы.

Заключение
Теперь, ознакомившись с предоставленной информацией, можно четко сформулировать, что представляет собой дигибридное скрещивание, основные особенности различных видов скрещивания растений, способы селекционного воспроизведения новых сортов. А также можно определить роль скрещивания для человека как потребителя растительной продукции.
Рассуждая аналогично, можно представить расщепление при три- и полигибридном скрещивании, т.е. когда родители различаются по аллелям трех и более генов, а в F1 образуются три- и полигетерозиготы. Соотношение генотипических и фенотипических классов в F2 три- и полигибридных скрещиваний, а также число типов гамет (и число фенотипов) у гибридов F1 определяются простыми формулами: при моногибридном скрещивании число типов гамет равно 2, при дигибридном 4(2 2 ), а при полигибридном – 2 n ; число генотипов равно соответственно 3, 9(3 2 ) и 3 n .
Полигибридное скрещивание – это скрещивание организмов, анализируемых по трем и более парам альтернативных признаков. Механизм наследования двух, трех и многих пар признаков, определяемых генами, лежащими в разных негомологичных хромосомах, в принципе не отличается от механизма наследования одной пары признаков. В основе этих скрещиваний лежит одна и та же закономерность.
Опираясь на независимость наследования разных пар аллелей, можно также любые сложные расщепления представить как произведение от соответствующего числа независимых моногибридных скрещиваний. Общая формула определения фенотипических классов при полигибридном скрещивании имеет вид (3:1) n , где п равно числу пар признаков, по которым идет расщепление. Для моногибрида эта формула соответственно имеет вид (3:1); дигибрида – 9:3:3:1 или (3:1) 2 ; тригибрида – (3:1) 3 . Расщепление по генотипу имеет вид (1:2; 1) n , где п – число расщепляющихся пар аллелей.
Известно, что каждый организм гетерозиготен по многим генам. Если предположить, что человек, у которого отдельные пары хромосом содержат не одну, а сотни пар аллелей, гетерозиготен хотя бы по 20 генам, то число типов гамет у такой полигетерозиготы составит 2 20 = 1 048 576. Эта цифра дает определенное представление о потенциальных возможностях комбинативной изменчивости. Поэтому каждый человек обладает неповторимой индивидуальностью. На Земле нет двух людей, совершенно одинаковых по наследственности, за исключением однояйцевых близнецов.
Таким образом, третий закон Менделя (закон независимого наследования признаков) еще раз демонстрирует дискретный характер генетического материала. Это проявляется в независимом комбинировании аллелей разных генов и в их независимом действии – фенотипическом выражении.
13.2. Дигибридное и полигибридное скрещивание. Независимое наследование
Дигибридное скрещивание – это скрещивание родительских особей, различающихся по двум парам альтернативных признаков и, соответственно, по двум парам аллельных генов.
Полигибридное скрещивание – это скрещивание особей, различающихся по нескольким парам альтернативных признаков и, соответственно, по нескольким парам аллельных генов.
Георг Мендель скрещивал растения гороха, отличающиеся по окраске семян (желтые и зеленые) и по характеру поверхности семян (гладкие и морщинистые). Скрещивая чистые линии гороха с желтыми гладкими семенами с чистыми линиями, имеющими зеленые морщинистые семена, он получил гибриды первого поколения с желтыми гладкими семенами (доминантные признаки). Затем Мендель скрестил гибриды первого поколения между собой и получил четыре фенотипических класса в соотношении 9: 3: 3: 1, т. е. в результате во втором поколении появилось два новых сочетания признаков: желтые морщинистые и зеленые гладкие. Для каждой пары признаков отмечалось отношение 3: 1, характерное для моногибридного скрещивания: во втором поколении получилось 3/4 гладких и 1/4 морщинистых семян и 3/4 желтых и 1/4 зеленых семян. Следовательно, две пары признаков объединяются у гибридов первого поколения, а затем разделяются и становятся независимыми друг от друга.
На основе этих наблюдений был сформулирован третий закон Менделя.
Третий закон Менделя
Закон о независимом наследовании: расщепление по каждой паре признаков идет независимо от других пар признаков. В чистом виде этот закон справедлив только для генов, локализованных в разных хромосомах, и частично соблюдается для генов, расположенных в одной хромосоме, но на значительном расстоянии друг от друга.
Опыты Менделя легли в основу новой науки – генетики. Генетика – это наука, изучающая наследственность и изменчивость.
Успеху исследований Менделя способствовали следующие условия:
1. Удачный выбор объекта исследования – гороха. Когда Менделю предложили повторить свои наблюдения на ястре-бинке, этом вездесущем сорняке, он не смог этого сделать.
2. Проведение анализа наследования отдельных пар признаков в потомстве скрещиваемых растений, отличающихся по одной, двум или трем парам альтернативных признаков. Велся учет отдельно по каждой паре этих признаков после каждого скрещивания.
3. Мендель не только зафиксировал полученные результаты, но и провел их математический анализ.
Мендель сформулировал также закон чистоты гамет, согласно которому гамета чиста от второго аллельного гена (альтернативного признака), т. е. ген дискретен и не смешивается с другими генами.
При моногибридном скрещивании в случае полного доминирования у гетерозиготных гибридов первого поколения проявляется только доминантный аллель, однако рецессивный аллель не теряется и не смешивается с доминантным. Среди гибридов второго поколения и рецессивный, и доминантный аллель может проявиться в своем – чистом – виде, т. е. в гомозиготном состоянии. В итоге гаметы, образуемые такой гетерозиготой, являются чистыми, т. е. гамета А не содержит ничего от аллели а, гамета а – чиста от А.
На клеточном уровне основой дискретности аллелей является их локализация в разных хромосомах каждой гомологичной пары, а дискретности генов – их расположение в разных локусах хромосом.
Анализ наследования одной аллельной пары в моногибридном скрещивании позволил Менделю и его последователям понять наследование двух и более пар признаков при дигибридном и полигибридном скрещиваниях.
Из всего ранее изложенного по этому поводу можно сделать следующие выводы:
- альтернативные признаки определяются аллелями гена, находящимися в гомологичных хромосомах;
- в каждой паре аллелей одна может быть доминантной, другая — рецессивной;
- во втором поколении моногибридного скрещивания происходит расщепление по доминантным и рецессивным признакам в определенном отношении; при полном доминировании расщепление по фенотипу 3:1, при неполном — 1:2:1, последнее отношение соответствует расщеплению по генотипу. Расщепление в F2 открыло принципиально важное явление, а именно: признаки скрещивающихся форм не пропадают в поколениях, а сохраняются;
- при дигибридном скрещивании обе аллельные пары, находящиеся в негомологичных парах хромосом, наследуются независимо друг от друга и вступают во все возможные комбинации с равной вероятностью, давая расщепление во втором поколении по фенотипу 9:3:3: 1 и по генотипу 1:2:2:4:1:2:1:2:1;
- явление расщепления и независимой комбинации разных пар аллелей генов обусловлено тем, что эти гены находятся в негомологичных хромосомах, которые случайно комбинируются при расхождении к полюсам в анафазе I мейоза.
Эти основные положения, установленные генетическим и цитологическим методами, позволяют сделать следующий шаг в анализе закономерностей наследования: выяснить их общее значение при полигибридном скрещивании.
Рассмотрим, например, тригибридное скрещивание . Можно заранее сказать, что основные закономерности наследования, установленные при моногибридном и дигибридном скрещиваниях, являются общими и для наследования при тригибридном скрещивании. Мендель для такого рода скрещивания избрал следующие три пары альтернативных признаков семян: гладкая — морщинистая форма семян, желтая — зеленая окраска семядолей, а также серо-коричневая и неокрашенная кожура, причем первыми в парах названы доминантные признаки, а вторыми — рецессивные.
В этом скрещивании материнское растение было гомозиготным по всем трем доминантным признакам, т. е. имело генотип ААВВСС, а отцовское — гомозиготно по трем рецессивным aabbcc. Все три гена находятся в трех негомологичных хромосомах. Тригибрид первого поколения имел структуру A/a B/b C/c. По внешнему виду семена тригибрида полностью походили на материнские, т. е. имели все три доминантных признака: гладкую форму, желтую окраску семядолей и серо-коричневую окраску семенной кожуры.
При указанной степени гетерозиготности гибриды F1 образуют уже восемь сортов гамет как женских, так и мужских, а именно: ABC, AbC, АВс, аВС, Abe, аВс, аbС и abc. При оплодотворении в результате сочетания восьми сортов женских и восьми сортов мужских гамет во втором поколении образуется 64 комбинации. В этом можно легко убедиться, воспользовавшись либо решеткой Пеннета, либо математическим способом перемножения вероятностей, как это мы делали раньше при дигибридном скрещивании. В результате подсчета обнаруживается, что у тригибрида расщепление в F2 по фенотипу включает 8 классов в определенном числовом отношении: 27 : 9 : 9 : 9 : 3 : 3 : 3 : 1.
Очевидно, для того чтобы выявить все 8 классов расщепления по фенотипу, необходимо иметь достаточно большое число растений в F2. И Мендель отмечает, что из всех опытов опыт с тригибридом потребовал наибольшего количества времени и труда. Чтобы убедиться в том, что тригибрид действительно образует 8 типов гамет в равных количествах, можно воспользоваться известным нам методом анализирующего скрещивания, который не требует столь большого числа потомков, как это необходимо для обнаружения восьми фенотипических классов в F2. Скрещивая тригибрид с формой, гомозиготной по всем трем рецессивным признакам (A/a B/b C/c x a/a b/b c/c), мы получим расщепление в отношении 1 : 1 : 1 : 1 : 1 : 1 : 1 : 1.
Если рассчитать расщепление в F2 по фенотипу отдельно для каждой пары альтернативных признаков, то расщепление в каждом случае окажется равным 3:1. Это исходное отношение, как мы знаем, обеспечивается точным цитологическим механизмом независимого расхождения пар хромосом в мейозе. Именно это числовое отношение в расщеплении по каждой паре признаков позволило Менделю и в тригибридном скрещивании подтвердить принцип независимого сочетания признаков в потомстве гибрида.
Принцип независимого поведения разных пар альтернативных признаков в расщеплении по фенотипу в F2 при полном доминировании может быть выражен формулой (3 + 1) n , где n — число пар альтернативных признаков.
Исходя из этой формулы, можно высчитать число ожидаемых классов в расщеплении по фенотипу при любом числе пар признаков, взятых в скрещивание:
- моногибридное скрещивание (3+1) 1 = 3 : 1, т. е. 2 класса,
- дигибридное скрещивание (3+1) 2 = 9: 3 : 3 : 1, т. е. 4 класса,
- тригибридное скрещивание (3 + 1) 3 = 27 : 9 : 9 : 9 : 3 : 3 : 3 : 1, т. е. 8 классов и т. д.
Иначе, говоря, число фенотипических классов при расщеплении в F2 может быть выражено формулой 2 n , где основание 2 указывает на парность генов (аллельность), находящихся в одной паре гомологичных хромосом, а степень n — число аллельных пар в негомологичных хромосомах, по которым различаются взятые в скрещивание родительские формы. Поэтому при моногибридном скрещивании число классов расщепления по фенотипу равно 2 1 = 2, при дигибридном 2 2 = 4, тригибридном 2 3 = 8, тетрагибридном 2 4 = 16 и т. д.
Таким же образом можно рассчитать число типов гамет, образующихся у любого полигибрида первого поколения (F1), и число комбинаций гамет, дающих различные генотипы в F2.
- у моногибрида A/a образуются два сорта гамет, или 2 1
- у дигибрида A/a B/b — четыре, или 2 2
- у тригибрида A/a B/b C/c — восемь сортов гамет, или 2 3
Следовательно, число различных типов гамет, образуемых гибридом F1, также может быть выражено формулой 2 n , где n — число пар аллельных генов, по которым различаются скрещиваемые формы.
Так как при моногибридном скрещивании у гибрида F1 образуются два сорта как женских, так и мужских гамет, то очевидно, что при этом возможно образование 4 комбинаций: 1А/А : 2А/a : 1a/a, т.е. 4 1 .
При дигибридном скрещивании таких сочетаний будет 4 2 = 16, при тригибридном 4 3 = 64 и т. д., т. е. число возможных комбинаций гамет выражается формулой 4 n , где основание 4 отражает число возможных комбинаций мужских и женских гамет в моногибридном скрещивании, а n — число пар аллелей.
Число генотипических классов рассчитывают при любом числе аллельных пар, по которым различаются родительские формы, взятые в скрещивание. В потомстве моногибрида расщепление по генотипу включает три класса: 1А/А : 2А/a : 1a/a.
При дигибридном скрещивании в F2 число генотипов соответствует 9 классам:
При тригибридном скрещивании число генотипических классов будет равно 3 3 .
Итак, число генотипических классов можно определить по формуле 3 n , где n — число аллельных пар. Таким образом, зная число пар аллелей при полигибридном скрещивании, можно рассчитать число типов гамет, образующихся у гибрида F1 число их сочетаний при оплодотворении, а также число фенотипических и генотипических классов.
Следует еще раз подчеркнуть, что все эти расчеты правомочны при том важном условии, если гены находятся в негомологичных хромосомах. Однако известно, что число последних для каждого Вида организмов является относительно небольшим и постоянным. Так, у гороха имеется всего 7 пар гомологичных хромосом, у человека — 23, у мушки дрозофилы — 4 и т. д. Следовательно, возможно одновременное независимое наследование лишь стольких генов, сколько пар гомологичных хромосом имеется у организмов Данного вида. Поскольку у гороха имеется 7 пар хромосом (n = 7), то у гибрида возможно независимое сочетание не более семи одновременно взятых в скрещивание пар признаков. При этом в потомстве гибридного растения гороха в F2 при полном доминировании возможно появление 2 7 различных фенотипических классов и 3 7 генотипических классов. Общее число возможных сочетаний гамет составит 4 7 . У классического для генетических исследований объекта — дрозофилы по независимо наследующимся парам признаков возможно только тетрагибридное скрещивание.
Очевидно, что если в скрещивание будет взято число аллельных пар, большее, чем имеющееся у данного организма число пар гомологичных хромосом, то третий закон Менделя не осуществится. В этом случае вступят в действие другие закономерности.
На первый взгляд это ограничение закона независимого сочетания генов может создать впечатление ограниченности наследственной изменчивости в силу небольшого числа возможных комбинаций гамет. Рассмотрим для примера возможный размах комбинативной изменчивости, возникающей в силу свободной комбинации гамет у человека. Допустим, что в каждой из 23 пар хромосом человека имеется только по одной паре аллелей: A/a, B/b, C/c, D/d, E/e и т.д.
При таком расчете число различных сортов гамет выразится величиной 2 23 , число классов расщепления по фенотипу — 2 23 , по генотипу — 3 23 , а число возможных комбинаций гамет — 4 23 , или 2 46 .
Для иллюстрации приводится число типов гамет и их сочетаний у индивидуумов, гетерозиготных по разному числу генов, от 1 до 23.
Подсчеты показывают, насколько велико число комбинаций гамет, которое может обеспечить наследственную изменчивость у человека, если различие одновременно касается 23 пар генов. Надо заметить, что в одном эякуляте мужчины число сперматозоидов колеблется от 2х10 9 до 5×10 9 . Следовательно, даже при таком огромном количестве сперматозоидов в одном эякуляте далеко не все указанные выше типы гамет могут встретиться.
Эти иллюстративные вычисления отражают лишь незначительную долю той изменчивости, которая в действительности возможна при свободном сочетании гамет. Во-первых, в каждой паре гомологичных хромосом может быть более одного гена. Во-вторых, при рассмотрении расщепления мы исходили из того, что разные пары аллелей в своем определении признаков не взаимодействуют между собой; на самом же деле это не так: в процессе индивидуального развития организма гены взаимодействуют. Существует и ряд других причин, которые могут приводить к увеличению изменчивости генотипов и фенотипов.
Несмотря на указанные моменты и тот факт, что каждый организм получает половину хромосом от матери, а половину от отца, наследственное сходство между родителями и потомством надежно обеспечивается. Комбинативная изменчивость представляет собой перекомбинацию наследственных возможностей родителей. На ограниченном числе потомков она не может быть выявлена. Лишь на большом числе потомков комбинативная изменчивость реализуется с большой полнотой.
В заключение изложения классической схемы анализа наследования признаков и свойств организмов следует сказать о том, в какой мере этот анализ можно и следует применять. Закономерности наследования, установленные Менделем на горохе, являются общими для всех организмов, имеющих половой процесс. Однако проявление их может видоизменяться в зависимости от объекта и влияния факторов внешней среды. Каждое закономерное явление природы осуществляется при определенных условиях. Если эти условия не способствуют его проявлению, то оно или совсем не проявится, или проявление его изменится. Но если сам метод обнаружения закономерного явления объективен, то с его помощью можно вскрыть новый характер проявления закономерности при измененных условиях. В этом сила научного метода.
Положение о независимом наследовании разных пар аллелей и признаков было подтверждено Менделем при изучении наследования трех пар признаков у гороха. Он скрещивал сорт растения с круглыми семенами (А), желтыми семядолями (В) и серо-коричневой кожурой семян (С) с сортом, форма семян которого морщинистая (о), семядоли зеленые (/>), семенная кожура белая (с). Материнское растение имело генотип ААВВСС, отцовское — ааЪЪсс. Поскольку родители гомозиготны по всем трем парам признаков, у них образуется по одному типу гамет: ABC и аЪс. Гибриды Fi будут иметь генотип АаВЬСс (тригетерозигота). При тригетерозиготности три пары разных аллелей находятся в трех разных парах гомологичных хромосом. В результате независимого сочетания хромосом (значит, и аллелей) из разных пар у гибрида Fi образуется восемь сортов гамет: ABC, ABc, АЬС, аВС, Abe, аВс, аЬс, аЬс. При самоопылении в результате случайного сочетания гамет в F2 получается 64 комбинации, включающие 8 фенотипов.
Мендель установил, что расщепление по фенотипу при три-гибридном скрещивании представляет собой сочетание трех независимых моногибридных расщеплений. Чем больше признаков, по которым отличаются взятые для скрещивания особи, тем сложнее расщепление и сильнее возрастает комбинативная изменчивость. Число возможных комбинаций гамет и число классов по фенотипу можно определить, пользуясь таблицей 3.
3. Количество фенотипов и генотипов в F2 при скрещивании родителей,
отличающихся разным числом пар признаков
(при полном доминировании)
| Число гамет, | Число | Число классов в F2 | Пропорция | ||
| Число пар признаков | образующихся в¥2 | возможных комбинаций гамет | по фенотипу | по генотипу | полных рецессивов в F2 |
| 2 3 4 п | 2 ! =2 2 2 =4 2 3 =8 2 4 =1б 2 я | 4 1= 4 4 2 =1б 4 3 =64 4 4 =25б 4" | 2 1 =2 2 2 =4 2 3 =8 2 4 =16 2" | 3 ! =3 3 2 =9 3 3 =27 3 4 =81 3 я | 1/4 1/16 1/64 1/256 1/4" |
, Для того чтобы понять, почему в пределах популяции каждого вида животных наблюдается такое большое разнообразие в типе телосложения, размерах, продуктивности и т. д., можно произвести простые расчеты при помощи формулы 2 я . Цифра 2 показывает, что набор хромосом диплоидный, и — гаплоидное число хромосом у определенного вида животных. Если отец и мать гетерозиготны только по одной какой-то паре аллельных генов, расположенных в каждой паре хромосом, то при полном доминировании каждого из признаков число определяемых этими аллелями возможных различных фенотипов у их потомков будет: у крупного рогатого скота 2 30 , или более миллиарда, у свиней 2 19 , или более 500 тыс., и т. д. Но животные, очевидно, могут отличаться друг от друга и по большему числу пар аллелей. Поэтому потенциальные возможности комбинативной изменчивости огромны, и становится понятным, почему в природе не встречается абсолютно похожих особей, за исключением однояйцевых близнецов.
Огромной заслугой Менделя является то, что в процессе своей работы он не только установил закономерности наследования признаков, но и открыл основные принципы (законы) наследственности:
1) дискретной (генной) наследственной детерминации признаков.
Этот принцип лежит в основе теории гена;
2) относительного постоянства наследственной единицы (гена);
3) аллельного состояния гена (доминантность и рецессивность).
ВЗАИМОДЕЙСТВИЕ НЕАЛЛЕЛЬНЫХ ГЕНОВ
Иногда на формирование признака влияют две или несколько пар неаллельных генов. Проявление признака в этом случае зависит от характера их взаимодействия в процессе развития организма. В первом поколении появляется новый признак, которого не было у исходных родительских форм, и соотношение фенотипов во втором поколении будет иным. Рассмотрим несколько примеров различных типов взаимодействия генов.

Новообразование. Новообразованием называется такой тип взаимодействия генов, когда при их сочетании в одном организме развивается совершенно новая форма признака. Известно, что у кур гены розовидного и стручковидного гребня не являются аллельными. И стручковидный, и розовидный гребень доминирует над листовидным. При скрещивании кур породы виан-дот, имеющих розовидный гребень (RRcc), с петухами породы брама со стручковидным гребнем (rrCQ у потомков первого поколения (RrCc) в результате взаимодействия двух доминантных генов Л и С развивается новая форма гребня — ореховидная (рис. 11). Скрещивание потомков Fi между собой ведет к получению в F2 четырех разных фенотипов в соотношении: 9 — с генами Ли Сс гребнем ореховидной формы, ЪЯсс — с гребнем розовидной формы, ЗггС — со стручковидным гребнем и irrvc
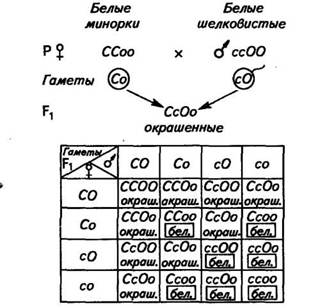
Комплементарное взаимодействие генов. В том случае, когда признак образуется при наличии двух доминантных неаллельных генов, каждый из которых не имеет самостоятельного фенотипи-ческого выражения, гены обозначают как комплементарные. Например, при скрещивании белых минорок с белыми шелковистыми курами первое поколение получается окрашенным. Для развития окраски необходимо, чтобы в организме синтезировались тирозин — предшественник меланина и фермент тирозингидрок-силаза, без которого пигмент не образуется. Обычно способность синтезировать какое-либо вещество доминирует над неспособностью к его образованию. Белые минорки имеют генотип ССоо. Они способны синтезировать тирозин, необходимый для образования
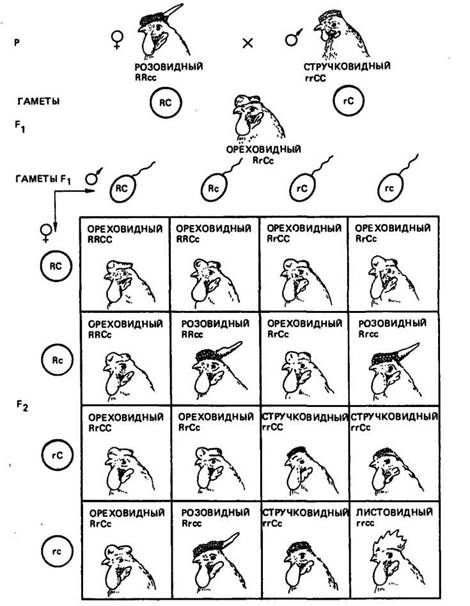
Рис. 11. Наследование формы гребня у кур при взаимодействии двух пар генов
пигмента, но не способны синтезировать фермент тирозингид-роксилазу, превращающий это вещество в пигмент. Белые шелковистые куры имеют генотип ссОО. Они не способны синтезировать тирозин, но обладают способностью синтезировать фермент. При спаривании таких кур между собой
пигмента в результате включения в генотип птиц Fi обоих доминантных генов — С (обусловливающего синтез тирозина) и О (обусловливающего синтез фермента). Во втором поколении окрашенных птиц ожидается 9 частей (СО..), а белых — 7 частей (С.оо—3, ссО..—Ъ и ссоо—1).
Комплементарным взаимодействием генов обусловлен, очевидно, особый тип паралича задних конечностей у помесных собак, полученных от скрещивания датского дога с сенбернаром. Генетический анализ, проведенный Стокардом, показал, что при чистопородном разведении как у датских догов, так и у сенбернаров паралич не наблюдается. В то же время из 57 помесей Fi, полученных от реципрокных скрещиваний указанных пород и доживших до 3-месячного возраста, только 3 или 4 не имели такого дефекта. Среди помесей F2 из 66 щенков, доживших до 3 мес, была парализована почти треть. Болезнь проявляется внезапно в возрасте около 3 мес. Тяжесть болезни может быть различной: от слабой парализованности до полной утраты способности к самостоятельному передвижению. Аналогичное заболевание встречается у некоторых помесных собак-ищеек.
Эпистаз. При этом типе взаимодействия доминантный ген одной пары аллелей, например ген С, подавляет действие другого неаллельного доминантного гена В. При генотипе ССВВ проявляются признаки, обусловленные геном С. Ген, подавляющий развитие другого признака, называется эпистатичным, а подавляемый — гипостатичным. Например, у лошади серая доминирующая масть, связанная с ранним поседением, перекрывает все другие масти. При скрещивании серой лошади генотипа ССВВ с масти. При скрещивании серой лошади генотипа ССВВ с рыжей генотипа ccbb в Fi все потомки будут серыми с генотипом СсВЬ. При скрещивании Fi между собой в F2 наблюдается расщепление по фенотипу: 12 серых, 3 вороные и 1 рыжая. Аллель серой масти (С) перекрывает действие других независимых генов окраски. Все лошади, имеющие в генотипе аллель С, будут серыми. Если аллель С отсутствует, при наличии в генотипе аллеля В лошадь будет вороной (ссВВ, ссВЬ), и лошадь с генотипом ccbb, двойным рецессивом будет рыжей окраски (рис. 12).
Полимерия. При полимерии, или полимерном (полигенном) наследовании, на один и тот же признак влияют несколько разных, но сходно действующих неаллельных генов. Каждый из них усиливает развитие признака. Такие однозначно действующие гены называются аддитивными. Впервые этот тип взаимодействия генов установлен Нильсоном-Эле при изучении наследования окраски чешуи овса и зерен пшеницы.
Рассмотрим пример наследования окраски зерен пшеницы при взаимодействии двух пар полимерных генов. Различают две основные окраски зерен: красную и белую. Полимерные гены, действующие на один и тот же признак, обозначают одинаковой буквой. Разные аллельные пары обозначают цифрами внизу букв. Исходя из этого, генотип пшеницы с темно-красным зерном будет А1А1А2А2, с белым зерном — аммяг. У первого родителя образуются гаметы Auii, у второго — а&ь В результате потомки Fi будут иметь генотип AiaA&i (двойная гетерозигота) и промежуточную окраску зерен — светло-красную, так как имеется два доминантных гена (Ai и А2), влияющих на проявление признака. Потомки первого поколения образуют по четыре сорта гамет (А1А2, Ajd2, аАг, aia2) и при спаривании между собой дадут F2, в котором расщепление по фенотипу и генотипу будет таким: из 16 частей 1 часть темно-красных (А1А1А2А2), четыре красных
Мендель установил, что расщепление по фенотипу при три-гибридном скрещивании представляет собой сочетание трех независимых моногибридных расщеплений. Чем больше признаков, по которым отличаются взятые для скрещивания особи, тем сложнее расщепление и сильнее возрастает комбинативная изменчивость. Число возможных комбинаций гамет и число классов по фенотипу можно определить, пользуясь таблицей 3.
3. Количество фенотипов и генотипов в F2 при скрещивании родителей,
отличающихся разным числом пар признаков
(при полном доминировании)
| Число гамет, | Число | Число классов в F2 | Пропорция | ||
| Число пар признаков | образующихся в¥2 | возможных комбинаций гамет | по фенотипу | по генотипу | полных рецессивов в F2 |
| 2 3 4 п | 2 ! =2 2 2 =4 2 3 =8 2 4 =1б 2 я | 4 1= 4 4 2 =1б 4 3 =64 4 4 =25б 4" | 2 1 =2 2 2 =4 2 3 =8 2 4 =16 2" | 3 ! =3 3 2 =9 3 3 =27 3 4 =81 3 я | 1/4 1/16 1/64 1/256 1/4" |
, Для того чтобы понять, почему в пределах популяции каждого вида животных наблюдается такое большое разнообразие в типе телосложения, размерах, продуктивности и т. д., можно произвести простые расчеты при помощи формулы 2 я . Цифра 2 показывает, что набор хромосом диплоидный, и — гаплоидное число хромосом у определенного вида животных. Если отец и мать гетерозиготны только по одной какой-то паре аллельных генов, расположенных в каждой паре хромосом, то при полном доминировании каждого из признаков число определяемых этими аллелями возможных различных фенотипов у их потомков будет: у крупного рогатого скота 2 30 , или более миллиарда, у свиней 2 19 , или более 500 тыс., и т. д. Но животные, очевидно, могут отличаться друг от друга и по большему числу пар аллелей. Поэтому потенциальные возможности комбинативной изменчивости огромны, и становится понятным, почему в природе не встречается абсолютно похожих особей, за исключением однояйцевых близнецов.
Огромной заслугой Менделя является то, что в процессе своей работы он не только установил закономерности наследования признаков, но и открыл основные принципы (законы) наследственности:
1) дискретной (генной) наследственной детерминации признаков.
Этот принцип лежит в основе теории гена;
2) относительного постоянства наследственной единицы (гена);
3) аллельного состояния гена (доминантность и рецессивность).
ВЗАИМОДЕЙСТВИЕ НЕАЛЛЕЛЬНЫХ ГЕНОВ
Иногда на формирование признака влияют две или несколько пар неаллельных генов. Проявление признака в этом случае зависит от характера их взаимодействия в процессе развития организма. В первом поколении появляется новый признак, которого не было у исходных родительских форм, и соотношение фенотипов во втором поколении будет иным. Рассмотрим несколько примеров различных типов взаимодействия генов.
Новообразование. Новообразованием называется такой тип взаимодействия генов, когда при их сочетании в одном организме развивается совершенно новая форма признака. Известно, что у кур гены розовидного и стручковидного гребня не являются аллельными. И стручковидный, и розовидный гребень доминирует над листовидным. При скрещивании кур породы виан-дот, имеющих розовидный гребень (RRcc), с петухами породы брама со стручковидным гребнем (rrCQ у потомков первого поколения (RrCc) в результате взаимодействия двух доминантных генов Л и С развивается новая форма гребня — ореховидная (рис. 11). Скрещивание потомков Fi между собой ведет к получению в F2 четырех разных фенотипов в соотношении: 9 — с генами Ли Сс гребнем ореховидной формы, ЪЯсс — с гребнем розовидной формы, ЗггС — со стручковидным гребнем и irrvc
Читайте также: