
Мобильные генетические элементы кратко
Обновлено: 02.07.2024
- Мобильные генетические элементы (МГЭ, англ. Mobile genetic elements, MGE) — последовательности ДНК, которые могут перемещаться внутри генома.
Существует несколько классов мобильных элементов генома, отличающихся по строению и способу перемещения:
* Транспозоны, например, Tn5;
* Инсерционные элементы, например, IS1603;
* Плазмиды, например, половой фактор кишечной палочки (F-плазмида);
* Бактериофаги, например, Mu, интегрирующиеся случайно в участки генома;
Связанные понятия
Повторя́ющиеся после́довательности ДНК (англ. Repetitive DNA) — участки ДНК, включённые в геном, последовательность которых состоит из повторяющихся фрагментов. Выделяют 2 типа таких повторяющихся последовательностей.
Ретротранспозоны (мобильные генетические элементы первого типа, или транспозоны, перемещающиеся через РНК-интермедиаты) — это генетические элементы, которые могут самовоспроизводиться в геноме и являются вездесущими компонентами ДНК многих эукариотических организмов.
Рекомбинация — перераспределение генетического материала (ДНК или РНК) путём разрыва и соединения разных молекул, приводящее к появлению новых комбинаций генов или других нуклеотидных последовательностей. В широком смысле слова включает в себя не только рекомбинацию между молекулами ДНК, но и перекомбинацию (сортировку) генетического материала на уровне целых хромосом или ядер, а также обмен плазмидами между клетками.
Упоминания в литературе
Исследования также показывают, что трихомонада прошла длительный эволюционный путь, на протяжении которого она заимствовала гены прежде всего у прокариот (кишечных бактерий). По этой причине в геноме трихомонады преобладают гены ферментов, участвующих в метаболизме углеводов и аминокислот, а также гены, кодирующие белки, всевозможные мобильные генетические элементы (в частности, встроенные фрагменты вирусных геномов, транспозоны, ретротранспозоны). На практике это привело к тому, что в последнее столетие геном трихомонады значительно увеличился в результате удвоения некоторых крупных фрагментов, иными словами, для пущей приспособленности к паразитическому образу жизни у трихомонады многократно увеличилось количество генов, необходимых для заглатывания отдельных белковых молекул и целых клеток организма хозяина. Это означает, что трихомонада еще не окончательно приспособилась к паразитированию в мочеполовых путях человека.
Связанные понятия (продолжение)
Дупликация (лат. duplicatio — удвоение) — разновидность хромосомных перестроек, при которой участок хромосомы оказывается удвоенным. Может произойти в результате неравного кроссинговера, ошибки при гомологичной рекомбинации, ретротранспозиции.
Горизонтальный перенос генов (ГПГ) — процесс, в котором организм передаёт генетический материал организму-непотомку. В отличие от горизонтального, о вертикальном переносе генов говорят, что при нем организм получает генетический материал от своего предка. В области интересов генетики основное место занимает вертикальный перенос генов. Однако в настоящее время горизонтальному переносу уделяется всё больше внимания.
РНК-интерференция (англ. RNA interference, RNAi) — процесс подавления экспрессии гена на стадии транскрипции, трансляции, деаденилирования или деградации мРНК при помощи малых молекул РНК.
Открытая рамка считывания (англ. Open Reading Frame, ORF) — последовательность нуклеотидов в составе ДНК или РНК, потенциально способная кодировать белок. Основным признаком наличия ORF служит отсутствие стоп-кодонов (в случае РНК — обычно UAA, UGA и UAG) на достаточно длинном участке последовательности после стартового кодона (в подавляющем большинстве случаев — AUG). Поскольку в некоторых случаях стартовый и терминирующие кодоны отличаются от канонических, а также ввиду возможности супрессии (подавления.
Энхансер (англ. enhancer — усилитель, увеличитель) — небольшой участок ДНК, который после связывания с ним факторов транскрипции стимулирует транскрипцию с основных промоторов гена или группы генов. Энхансеры не обязательно находятся в непосредственной близости от генов, активность которых они регулируют, и даже не обязательно располагаются с ними на одной хромосоме. Энхансеры могут располагаться как в 5'-, так и в 3'-положении относительно матричной цепи регулируемого гена и в любой ориентации к.
Консервати́вные после́довательности (англ. conserved sequences) — схожие или идентичные последовательности, встречающиеся в биологических полимерах: нуклеиновых кислотах, первичной и пространственной структурах белков, полисахаридах как в пределах особей разных видов (ортологичные последовательности), так и в пределах одной особи (паралогичные последовательности). Ортологичные последовательности являются подтверждением того, что определённые последовательности могут поддерживаться эволюцией, несмотря.
Гомеобокс (англ. homeobox) — последовательность ДНК, обнаруженная в генах, вовлечённых в регуляцию развития у животных, грибов и растений. Эти гены кодируют факторы транскрипции, которые, как правило, переключают каскады других генов. Гомеобокс состоит приблизительно из 180 пар нуклеотидов и кодирует белковый домен длиной в 60 аминокислот (гомеодомен), который может связывать ДНК.
Подавление экспрессии генов (сайленсинг генов от англ. gene silencing, или в частности, выключение гена) — это общий термин, описывающий эпигенетический процесс регуляции генов. При этом последовательность нуклеотидов не изменяется, а лишь прекращается экспрессия соответствующего гена. Для выключения генов в лабораторных условиях применяют метод нокдауна генов.
Трансду́кция (от лат. transductio — перемещение) — процесс переноса бактериальной ДНК из одной клетки в другую бактериофагом. Общая трансдукция используется в генетике бактерий для картирования генома. К трансдукции способны как умеренные фаги, так и вирулентные, последние, однако, уничтожают популяцию бактерий, поэтому трансдукция с их помощью не имеет большого значения ни в природе, ни при проведении исследований.
Оперон — функциональная единица генома у прокариот, в состав которой входят цистроны (гены, единицы транскрипции), кодирующие совместно или последовательно работающие белки и объединенные под одним (или несколькими) промоторами. Такая функциональная организация позволяет эффективнее регулировать транскрипцию этих генов.
Гено́м — совокупность наследственного материала, заключенного в клетке организма. Геном содержит биологическую информацию, необходимую для построения и поддержания организма. Большинство геномов, в том числе геном человека и геномы всех остальных клеточных форм жизни, построены из ДНК, однако некоторые вирусы имеют геномы из РНК.
Последовательность Шайна — Дальгарно (англ. Shine-Dalgarno sequence, Shine-Dalgarno box) — сайт связывания рибосом на молекуле мРНК прокариот, обычно на расстоянии около 10 нуклеотидов до стартового кодона AUG. Описана австралийскими учёными Джоном Шайном и Линн Дальгарно.Консенсусом является последовательность из шести нуклеотидов AGGAGG; в случае E. coli последовательность Шайна — Дальгарно — AGGAGGU. Комплементарная последовательность CCUCCU, называемая последовательностью анти-Шайна — Дальгарно.
Комплементарная ДНК (кДНК, англ. сDNA) — это ДНК, синтезированная на матрице зрелой мРНК в реакции, катализируемой обратной транскриптазой.
Изоформа белка — любая из нескольких разных форм одного и того же белка. Различные формы белка могут быть образованы связанными генами, или могут возникнуть из того же гена путём альтернативного сплайсинга. Большое количество изоформ вызваны однонуклеотидными полиморфизмами — небольшими генетическими различиями между аллелями одного и того же гена. Это происходит в определенных отдельных местах расположения нуклеотидов на гене.
Транскрипт — молекула РНК, образующаяся в результате транскрипции (экспрессии соответствующего гена или участка ДНК).
Цис-регуляторные элементы (или цис-элементы) — участки ДНК или РНК, регулирующие экспрессию генов, находящихся на той же молекуле (обычно хромосоме).
Конденсины — большие белковые комплексы, которые играют главную роль в расхождении хромосом во время митоза и мейоза.
Метилирование ДНК — это модификация молекулы ДНК без изменения самой нуклеотидной последовательности ДНК, что можно рассматривать как часть эпигенетической составляющей генома.
Эндоге́нные ви́русные элеме́нты (англ. Endogenous viral elements) — последовательности ДНК вирусного происхождения в геноме невирусных организмов, которые присутствуют в клетках зародышевой линии и передаются по наследству. Иногда эндогенные вирусные элементы представлены полными вирусными геномами (провирусами), в других случае они являются фрагментами вирусных геномов. Провирусы могут сохранять потенциальную способность вызывать инфекцию, опосредуя образование новых вирусных частиц. При удвоении.
Гомологи́чные хромосо́мы — пара хромосом приблизительно равной длины, с одинаковым положением центромеры и дающие одинаковую картину при окрашивании. Их гены в соответствующих (идентичных) локусах представляют собой аллельные гены — аллели, то есть кодируют одни и те же белки или РНК. При двуполом размножении одна гомологичная хромосома наследуется организмом от матери, а другая — от отца.
Плазми́ды (англ. plasmids) — небольшие молекулы ДНК, физически обособленные от хромосом и способные к автономной репликации. Главным образом плазмиды встречаются у бактерий, а также у некоторых архей и эукариот (грибов и высших растений). Чаще всего плазмиды представляют собой двухцепочечные кольцевые молекулы. Несмотря на способность к размножению, плазмиды, как и вирусы, не рассматриваются в качестве живых организмов.
Центромера — участок хромосомы, который связывает сестринские хроматиды, играет важную роль в процессе деления клеточного ядра и участвует в контроле экспрессии генов. Характеризуется специфическими последовательностью нуклеотидов и структурой.
Эпигенетическим наследованием называют наследуемые изменения в фенотипе или экспрессии генов, вызываемые механизмами, отличными от изменения последовательности ДНК (приставка эпи- означает в дополнение). Такие изменения могут оставаться видимыми в течение нескольких клеточных поколений или даже нескольких поколений живых существ.
Нуклео́ид (англ. Nucleoid) — неправильной формы зона в цитоплазме прокариотической клетки, в которой находится геномная ДНК и ассоциированные с ней белки. На долю ДНК приходится около 60 % массы нуклеоида; помимо ДНК, нуклеоид содержит РНК и белки. Белки нуклеоида, которые обеспечивают пространственную организацию геномной ДНК, называют нуклеоидными белками или нуклеоид-ассоциированными белками; они не имеют ничего общего с гистонами, упаковывающими ДНК у эукариот. В отличие от гистонов, ДНК-связывающие.
Флуоресце́нтная гибридиза́ция in situ, или метод FISH (англ. fluorescence in situ hybridization — FISH), — цитогенетический метод, который применяют для детекции и определения положения специфической последовательности ДНК на метафазных хромосомах или в интерфазных ядрах in situ. Кроме того, FISH используют для выявления специфических мРНК в образце ткани. В последнем случае метод FISH позволяет установить пространственно-временные особенности экспрессии генов в клетках и тканях.
Вектор (в генетике) — молекула нуклеиновой кислоты, чаще всего ДНК, используемая в генетической инженерии для передачи генетического материала внутрь клетки, в том числе в клетку живого многоклеточного организма in vivo.
Некодирующая ДНК или Мусорная ДНК (англ. Non-coding DNA англ. junk DNA) — части геномной ДНК организмов, которые не кодируют последовательности белков. Некоторые некодирующие ДНК переводятся в функциональные некодирующие РНК-молекулы. Другие функции некодирующей ДНК включают регуляцию последовательностей кодирующих белки, центромер и теломер.
Делеции (от лат. deletio — уничтожение) — хромосомные перестройки, при которых происходит потеря участка хромосомы. Делеция может быть следствием разрыва хромосомы или результатом неравного кроссинговера. По положению утерянного участка хромосомы делеции классифицируют на внутренние (интерстициальные) и концевые (терминальные).
Теломе́ры (от др.-греч. τέλος — конец и μέρος — часть) — концевые участки хромосом. Теломерные участки хромосом характеризуются отсутствием способности к соединению с другими хромосомами или их фрагментами и выполняют защитную функцию.
Амплификация (лат. amplificatio — усиление, увеличение), в молекулярной биологии — процесс образования дополнительных копий участков хромосомной ДНК, как правило, содержащих определённые гены либо сегменты структурного гетерохроматина. Амплификация может быть ответом клеток на селективное воздействие (например, при действии метотрексата). Амплификация — один из механизмов активации онкогенов в процессе развития опухоли, например, онкогена N-myc при развитии нейробластомы. Также амплификация — накопление.
Ми́кроРНК (англ. microRNA, miRNA) — малые некодирующие молекулы РНК длиной 18—25 нуклеотидов (в среднем 22), обнаруженные у растений, животных и некоторых вирусов, принимающие участие в транскрипционной и посттранскипционной регуляции экспрессии генов путём РНК-интерференции. Помимо внутриклеточной обнаружена внеклеточная (циркулирующая) микроРНК.МикроРНК кодируются ядерной ДНК растений и животных и вирусной ДНК у некоторых ДНК-содержащих вирусов.
Нуклеосома — это структурная часть хромосомы, образованная совместной упаковкой нити ДНК с гистоновыми белками H2А, H2B, H3 и H4. Последовательность нуклеосом, соединенная гистоновым белком H1, формирует нуклеофиламент, или иначе нуклеосомную нить.
Ретрови́русы (лат. Retroviridae, от лат. retro — обратный) — семейство РНК-содержащих вирусов, заражающих преимущественно позвоночных. Наиболее известный и активно изучаемый представитель — вирус иммунодефицита человека.
Рекомбинантная структура (англ. Recombinant structure) — гибридная (англ. recombination — рекомбинация) нуклеиновая кислота (ДНК или РНК) или белок, полученные в результате объединения in vitro чужеродных фрагментов и содержащие новые сочетания последовательностей нуклеотидов или аминокислот соответственно.
Точечная мутация — тип мутации в ДНК или РНК, для которого характерна замена одного азотистого основания другим. Термин также применяется и в отношении парных замен, инсерции или делеции одного или нескольких нуклеотидов. Точечные мутации, возникающие в некодирующей ДНК, обычно никак себя не проявляют. Точечный мутант — организм, в генотипе которого произошла точечная мутация.
Соматические клетки (др.-греч. σῶμα — тело) — клетки, составляющие тело (сому) многоклеточных организмов и не принимающие участия в половом размножении. Таким образом, это все клетки, кроме гамет.
Мобильные генетические элементы (прыгающие гены) - участки ДНК, способные к транспозиции, или случайному перемещению, из одного места в другое: в пределах одной молекулы ДНК, из одной ДНК в другую. Не способны к самостоятельной репликации. Размножаются в составе бактериальной хромосомы или плазмид. К подвижным генетическим элементам относятся IS элементы, транспозоны, интегроны.
Транспозиция обеспечивается ферментом – транспозазой. Ген, кодирующий этот фермент, входит в состав всех мобильных генетических элементов. Транспозаза обладает эндонуклеазной и лигазной функцией: она разрезает ДНК по краям мобильного генетического элемента (эндонуклеазная функция) и сшивает его с разрывом ДНК в новом месте (лигазная функция). В некоторых случаях транспозиция сопровождается удвоением мобильных генетических элементов и перемещением копии в другое место.
Инсерционные последовательности, или IS элементы. Это разновидность мобильных генетических элементов, которые не несут в своем составе структурные гены, а только гены, отвечающие за перемещение. Многообразие IS элементов обозначают цифровыми индексами - IS1, IS 6010. Их размер меньше, чем транспозонов, и составляет от 700 до 1800 п.о., но описаны IS элементы более крупных и мелких размеров — 5700 п.о. и 200 п.о. Центральную часть IS элемента занимает ген, кодирующий траспозазу; некоторые IS элементы несут промоторы или репрессоры генов, или их части. На обоих концах IS элемента находятся повторяющиеся последовательности, или палиндромы, размером 10 – 40 п.о., по которым транспозаза распознает его и вырезает. В геноме бактерий присутствует, как правило, небольшое количество их копий: в геноме E. coli IS1 встречается в 6 - l0 копиях, а IS2 – 5 копиях. Транспозиция происходит двумя путями: 1) консервативным: покидая один участок IS элемент встраивается в другой; 2) репликативным: синтезируется копия, которая встраивается в другой участок генома. Встраивание, как правило, осуществляется в участках богатых А/Т.
Значение IS элементов.1) Участвуют в мутационной изменчивости микроорганизмов – инсерциях и делециях. Инсерция IS элементов в бактериальную ДНК приводит к синтезу неполноценного белка. При встраивании некоторых IS элементов по обоим их концам происходит удвоение небольшого участка хромосомы размером 5 - 9 п.о. С меньшей частотой (10 -3 -10 -4 ) IS элементы приводят к делециям в прилегающих генах: покидая ДНК, IS4 вырезает участки хромосомальной ДНК по обоим своим концам.
2. Являются генетическими маркерами вида или рода бактерий. Некоторые IS последовательности специфичны для определенных видов микроорганизмов, что позволяет их использовать для видовой идентификации бактерий.
3. Являются местом распознавания и встраивания плазмид и генно-инженерных векторов. Плазмиды и генно-инженерными векторы встраиваться в бактериальную хромосому в области IS последовательностей.
4. Участвуют в регуляции функций генов – активации или репрессии, так как несут в своем составе промоторы или репрессоры генов или их компоненты. Например, формирование резистентности к метронидазолу у анаэробных микроорганизмов связано с активацией молчащих nim A, B, C, D, E генов в результате встраивания IS элементов, несущих промоторы этих генов.
Транспозоны. Это разновидность мобильных генетических элементов, которые содержат в своем составе один или несколько структурных генов и гены, ответственные за перемещение. Транспозоны обозначают Tn с числовым индексом, например, Tn 4556. Их размер больше IS элементов и составляет 2,500 - 7,000 п.о. На обоих концах Tn находятся прямые или инвертированные повторы, по которым транспозаза распознает их и вырезает. Частота транспозиций Tn сравнима с частотой мутаций. В зависимости от структуры выделяют два класса транспозонов:
1. Сборные(Tn 5, 9, 10, 903 и 1681). Состоят из фенотипического модуля (гены резистентности) и двух располагающихся по краям IS элементов, ориентированных в одном или противоположных направлениях. IS элементы обеспечивают перемещение транспозонов, но могут покидать его и перемещаться самостоятельно.
Tn10, имеющий по краям две копии IS10, подвергается переносу с частотой 10 -7 . Этот транспозон встраивается, преимущественно, в участках с последовательностью GCTNAGC (при встраивании этот участок удваивается), как правило, полностью безошибочно вырезается, но в некоторых случаях в процессе эксцизии может захватывать из бактериальной хромосомы дополнительно 50 п.о.

Бактериофаг m, или мутатор, также относится к комплексным транспозонам. Этот самый сложно организованный транспозон содержит 38 000 п.о. и на его концах находятся инвертированные повторы размером 11 п.о. Он не имеет определенного сайта встраивания в бактериальную ДНК, в процессе вырезания из нее обычно захватывает участок размером 10% своего размера. Бактериофаг m часто используют в генетических исследованиях, так как в естественных условиях он не входит в состав бактериальных геномов и потому легко проводить его детекцию.
Перенос транспозонов осуществляется консервативным или репликативным механизмом. Консервативный перенос происходит путем вырезания транспозона из одного участка и транспозицией в другой без увеличения количества копий, при этом участок ДНК, откуда вырезается транспозон, утрачивает свои функции. При репликативном способе переноса синтезированная копия транспозона перемещается в новое место, при этом механизме увеличивается количество копий.
Функции. Транспозоны участвуют в регуляции активности генов, инактивируя их или активируя. Осуществляют горизонтальный перенос генов, например, вирулентности или резистентности, обуславливая распространение устойчивости к антибиотикам среди микроорганизмов.
Интегроны.Отвечают за сайт-специфическую рекомбинацию. Интегроны - мелкие генетические элементы, содержащие промотор и ген тирозиновой рекомбиназы - int, которая распознает и обеспечивает встраивание в сайт att I бактериальной хромосомы. Не содержат гены, отвечающие за транспозицию. Способны соединяться с кассетами генов, кодирующими резистентность и другие признаки, при этом генетические кассеты должны содержать обеспечивающие подвижность гены и элемент размером 59 п.о.
Мобильные генетические элементы (прыгающие гены) - участки ДНК, способные к транспозиции, или случайному перемещению, из одного места в другое: в пределах одной молекулы ДНК, из одной ДНК в другую. Не способны к самостоятельной репликации. Размножаются в составе бактериальной хромосомы или плазмид. К подвижным генетическим элементам относятся IS элементы, транспозоны, интегроны.
Транспозиция обеспечивается ферментом – транспозазой. Ген, кодирующий этот фермент, входит в состав всех мобильных генетических элементов. Транспозаза обладает эндонуклеазной и лигазной функцией: она разрезает ДНК по краям мобильного генетического элемента (эндонуклеазная функция) и сшивает его с разрывом ДНК в новом месте (лигазная функция). В некоторых случаях транспозиция сопровождается удвоением мобильных генетических элементов и перемещением копии в другое место.
Инсерционные последовательности, или IS элементы. Это разновидность мобильных генетических элементов, которые не несут в своем составе структурные гены, а только гены, отвечающие за перемещение. Многообразие IS элементов обозначают цифровыми индексами - IS1, IS 6010. Их размер меньше, чем транспозонов, и составляет от 700 до 1800 п.о., но описаны IS элементы более крупных и мелких размеров — 5700 п.о. и 200 п.о. Центральную часть IS элемента занимает ген, кодирующий траспозазу; некоторые IS элементы несут промоторы или репрессоры генов, или их части. На обоих концах IS элемента находятся повторяющиеся последовательности, или палиндромы, размером 10 – 40 п.о., по которым транспозаза распознает его и вырезает. В геноме бактерий присутствует, как правило, небольшое количество их копий: в геноме E. coli IS1 встречается в 6 - l0 копиях, а IS2 – 5 копиях. Транспозиция происходит двумя путями: 1) консервативным: покидая один участок IS элемент встраивается в другой; 2) репликативным: синтезируется копия, которая встраивается в другой участок генома. Встраивание, как правило, осуществляется в участках богатых А/Т.
Значение IS элементов.1) Участвуют в мутационной изменчивости микроорганизмов – инсерциях и делециях. Инсерция IS элементов в бактериальную ДНК приводит к синтезу неполноценного белка. При встраивании некоторых IS элементов по обоим их концам происходит удвоение небольшого участка хромосомы размером 5 - 9 п.о. С меньшей частотой (10 -3 -10 -4 ) IS элементы приводят к делециям в прилегающих генах: покидая ДНК, IS4 вырезает участки хромосомальной ДНК по обоим своим концам.
2. Являются генетическими маркерами вида или рода бактерий. Некоторые IS последовательности специфичны для определенных видов микроорганизмов, что позволяет их использовать для видовой идентификации бактерий.
3. Являются местом распознавания и встраивания плазмид и генно-инженерных векторов. Плазмиды и генно-инженерными векторы встраиваться в бактериальную хромосому в области IS последовательностей.
4. Участвуют в регуляции функций генов – активации или репрессии, так как несут в своем составе промоторы или репрессоры генов или их компоненты. Например, формирование резистентности к метронидазолу у анаэробных микроорганизмов связано с активацией молчащих nim A, B, C, D, E генов в результате встраивания IS элементов, несущих промоторы этих генов.
Транспозоны. Это разновидность мобильных генетических элементов, которые содержат в своем составе один или несколько структурных генов и гены, ответственные за перемещение. Транспозоны обозначают Tn с числовым индексом, например, Tn 4556. Их размер больше IS элементов и составляет 2,500 - 7,000 п.о. На обоих концах Tn находятся прямые или инвертированные повторы, по которым транспозаза распознает их и вырезает. Частота транспозиций Tn сравнима с частотой мутаций. В зависимости от структуры выделяют два класса транспозонов:
1. Сборные(Tn 5, 9, 10, 903 и 1681). Состоят из фенотипического модуля (гены резистентности) и двух располагающихся по краям IS элементов, ориентированных в одном или противоположных направлениях. IS элементы обеспечивают перемещение транспозонов, но могут покидать его и перемещаться самостоятельно.
Tn10, имеющий по краям две копии IS10, подвергается переносу с частотой 10 -7 . Этот транспозон встраивается, преимущественно, в участках с последовательностью GCTNAGC (при встраивании этот участок удваивается), как правило, полностью безошибочно вырезается, но в некоторых случаях в процессе эксцизии может захватывать из бактериальной хромосомы дополнительно 50 п.о.
Бактериофаг m, или мутатор, также относится к комплексным транспозонам. Этот самый сложно организованный транспозон содержит 38 000 п.о. и на его концах находятся инвертированные повторы размером 11 п.о. Он не имеет определенного сайта встраивания в бактериальную ДНК, в процессе вырезания из нее обычно захватывает участок размером 10% своего размера. Бактериофаг m часто используют в генетических исследованиях, так как в естественных условиях он не входит в состав бактериальных геномов и потому легко проводить его детекцию.
Перенос транспозонов осуществляется консервативным или репликативным механизмом. Консервативный перенос происходит путем вырезания транспозона из одного участка и транспозицией в другой без увеличения количества копий, при этом участок ДНК, откуда вырезается транспозон, утрачивает свои функции. При репликативном способе переноса синтезированная копия транспозона перемещается в новое место, при этом механизме увеличивается количество копий.
Функции. Транспозоны участвуют в регуляции активности генов, инактивируя их или активируя. Осуществляют горизонтальный перенос генов, например, вирулентности или резистентности, обуславливая распространение устойчивости к антибиотикам среди микроорганизмов.
Интегроны.Отвечают за сайт-специфическую рекомбинацию. Интегроны - мелкие генетические элементы, содержащие промотор и ген тирозиновой рекомбиназы - int, которая распознает и обеспечивает встраивание в сайт att I бактериальной хромосомы. Не содержат гены, отвечающие за транспозицию. Способны соединяться с кассетами генов, кодирующими резистентность и другие признаки, при этом генетические кассеты должны содержать обеспечивающие подвижность гены и элемент размером 59 п.о.

Обзор
Многообразие мобильных генетических элементов
Автор
Редакторы

Организмы и геномы можно таким образом расценивать как секции биосферы, по которым гены глобально циркулируют с различной интенсивностью, и в которые персональные гены и опероны могут включаться, если они предоставляют достаточные преимущества.
K. Jeon и J. Danielli [1]

Основные сокращения
МГЭ мобильные генетические элементы ГГП горизонтальный генетический перенос ОРС открытая рамка считывания, последовательность нуклеотидов между инициирующим и терминирующим кодонами гена ori T место начала переноса цепи плазмидной ДНК при мобилизации ori V место начала репликации (копирования, воспроизводства) плазмидной ДНК rep ген белка-инициатора репликации многих бактериальных плазмид
Вирусы: суперпаразиты и помощники
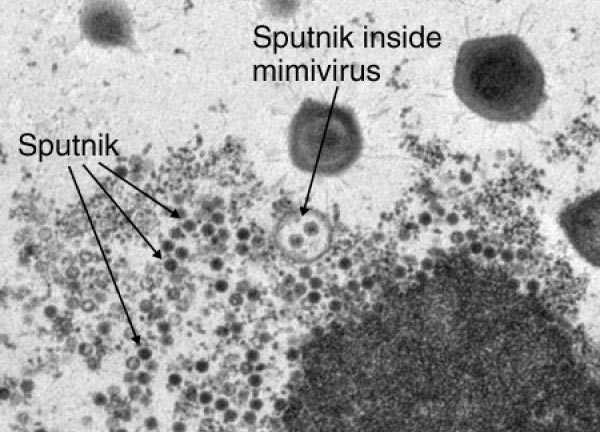
В 2012 году охарактеризовали интегрированную в ДНК мимивирусов форму вирофага и даже новый класс МГЭ — трансповироны, способные встраиваться в ДНК и мимимирусов, и вирофагов [7]. Подобно бактериофагам, переносящим гены от одних бактерий к другим, вирофаги могут играть важную роль в ГГП между разными группами вирусов и их хозяевами.
Многие профаги кодируют факторы вирулентности, трансформируя нетоксигенные бактерии в агентов-убийц: профаг CTXphi из Vibrio cholerae кодирует холерный токсин, а упомянутый выше P22 — ферменты конверсии О-антигена сальмонеллы, позволяя ей уходить от иммунного надзора.
В составе фагов обнаруживают другие МГЭ (например, транспозоны, несущие гены антибиотикорезистентности) и их модули (системы репликации и переноса конъюгативных плазмид). Самый известный пример слияния модулей фагового и плазмидного происхождения — фаг Р1, способный реплицироваться и длительно поддерживаться вне хромосомы, ничем не отличаясь от плазмиды. Профаги разных бактерий детально рассмотрены в обзоре [8].
Плазмиды — маленькие гиганты больших процессов
Плазмиды — внехромосомные двуцепочечные молекулы ДНК, способные к длительному автономному существованию в клетках прокариот и некоторых эукариот. Чаще всего плазмидные ДНК суперскручены и ковалентно замкнуты в кольцо, однако у актиномицетов и спирохет встречаются и линейные формы, что обычно сочетается с линейной организацией хромосом. Размер плазмид обычно варьирует от 0,85 т.п.н. (pRKU1 из Thermotoga petrophila) до 600 т.п.н., но у бруцелл и ризобий описаны мегаплазмиды размером более 1 млн п.н., что иногда делает вопрос их дифференцировки от дополнительных хромосом риторическим [9]. Элиминация мегаплазмид, в отличие от хромосом, обычно не вызывает фатальных для бактериальной клетки последствий, однако вместе с плазмидами могут утратиться такие важные функции, как способность к фиксации азота и формированию симбиотических клубеньков (у ризобий).
На долю плазмидной ДНК может приходиться 1–15% наследственной информации бактериальной клетки, однако известны случаи, когда плазмидами контролируется до 25% (у некоторых Archaea) и даже до 40% информации (2 мегаплазмиды размером около 1,4 и 1,6 млн п.н. у Sinorhizobium meliloti).
Плазмидами часто мобилизуются сосуществующие с ними в одной клетке неконъюгативные МГЭ (но обладающие как минимум сайтом начала переноса oriT, а чаще и генами mob) и даже хромосомы. Мобилизация генов хромосомы возможна в случае интеграции в неё конъюгативной плазмиды. Это возможно, когда в обеих молекулах присутствуют одинаковые инсерционные последовательности (IS-элементы), обеспечивающие гомологичную рекомбинацию (типичный пример — F-фактор E.coli). Однако интеграция может быть и RecA-независимой, если плазмида кодирует тирозиновую интегразу (плазмиды стрептомицетов pSE101, pSAM2 и псевдомонад pKLK106). В этом случае интеграция чаще происходит в гены тРНК, но в некоторых хозяевах эта закономерность нарушается [3].
Транспозоны — универсальные генетические челноки
Транспозоны — МГЭ, перемещающиеся как в пределах одной молекулы ДНК, так и между разными репликонами одного генома (конъюгативные транспозоны — и между геномами). Фланкированы инвертированными повторами, а в центральной части содержат гены, ответственные за перемещение. Транспозоны прокариот подразделяют на IS-элементы, Tn-элементы и Mu-подобные фаги [10].
Интегроны — природные системы клонирования и экспрессии
В процессе перемещения от одного интегрона к другому или от одного сайта в интегроне к другому сайту, генная кассета существует как автономная и неспособная к репликации двунитевая кольцевая молекула ДНК. Кассеты, захватываемые интегронами и суперинтегронами, могут содержать гены факторов патогенности, метаболических путей, детерминанты антибиотико- и дезинфектантоустойчивости или гены рестрикционных ферментов. Интеграза IntI катализирует сайт-специфическую рекомбинацию между сайтами attI и attC, в результате чего происходит интеграция или вырезание кассеты. Множество событий интеграции ведет к образованию мультикассетных рядов, в которых все кассеты фланкированы attC-сайтами. Известны хромосомные суперинтегроны, включающие до 179 генных кассет (у Vibrio cholerae), однако среди клинически значимых бактерий большинство интегронов содержит до 5–8 генных кассет. Наиболее эффективно экспрессируются кассеты, расположенные ближе к промотору, но изменение селективного давления может способствовать перестройкам в составе интегрона.

Рисунок 3. Интегрон — ДНК, улавливающая генные кассеты и распространяемая в составе более высокоорганизованных МГЭ. а — Структура интегрона класса 1. Pint — промотор интегразы, Pant — промотор кассет антибиотикорезистентности. Остальные элементы объяснены в тексте; б — Иерархическая организация МГЭ.
Все интегроны, несущие кассеты антибиотикорезистентности, разделяют на 5 классов на основании гомологии последовательностей кодируемых ими интеграз. Большинство интегронов антибиотикорезистентности относится к классу 1 (часто ассоциированы с Tn21-семейством). Они включают два концевых невариабельных региона, называемых константными последовательностями (constant sequences, CS), и высоковариабельный центральный участок. В одном конце интегрона (5’-CS), обычно находятся intI, attI и промотор, от которого экспрессируются гены кассеты. В другом конце (3’-CS), находится часть гена qacEΔ1, который, будучи интактным, несёт устойчивость к четвертичным аммониевым соединениям. За ним расположен ген sul, определяющий резистентность микроорганизма к сульфаниламидам, и 1–2 гена с неустановленной функцией — orf5 и иногда orf6 (рис. 3, а). Интегроны класса 2 ассоциированы с Tn7-семейством, классов 3 и 5 — с плазмидами, класса 4 — с конъюгативным геномным островом SXT Vibrio cholerae.

Рисунок 4. Схематическое изображение структуры геномного острова в составе бактериальной хромосомы. DR — прямые повторы ДНК хромосомы, фланкирующие ГО; IS — инсерционные элементы.
Геномные острова гонококков (GGI) кодируют систему секреции типа IV (T4SS), родственную плазмидным системам конъюгационного переноса. Посредством T4SS распространяется не только сам элемент и его продукты, но секретируется в окружающую среду и хромосома Neisseria gonorrhoeae, которая затем может трансформировать другие бактерии и участвовать в рекомбинационных событиях.
Главные нарушители границ и их мекка
Из обитателей отстойников, а что самое опасное — из бактерий вод, уже прошедших очистку, — в большом количестве выделяют плазмиды IncP-1-группы несовместимости (в системе классификации плазмид псевдомонад) [12]. Эти относительно небольшие молекулы дарят хозяевам массу селективных преимуществ (от множественной резистентности до биодеградации хлорорганики) и способны распространяться не только среди псевдомонад, но и практически всех грамотрицательных и некоторых грамположительных бактерий. Кроме того, они способны мобилизовать неконъюгативные R-плазмиды (например, IncP-4) к переносу в ещё более широкий спектр бактерий, а также в дрожжи и клеточные линии млекопитающих [17].
Биология плазмид в России и за рубежом. Что же можно возвести на крепком фундаменте?
МОБИ́ЛЬНЫЕ ГЕНЕТИ́ЧЕСКИЕ ЭЛЕМЕ́Н ТЫ (МГЭ, транспозоны), дискретные нуклеотидные фрагменты ДНК с непостоянной локализацией в хромосоме; способны к транспозиции – перемещению из одного участка хромосомы (донорного) в другой (реципиентный). МГЭ присутствуют в геномах всех организмов; их размеры варьируют от нескольких сотен до нескольких тысяч пар нуклеотидов (п. н.). Они могут быть рассеяны по хромосомам или же группироваться в отдельных (гетерохроматиновых) участках хромосом. Для одних МГЭ выбор места перемещения может быть достаточно случаен, для других – ограничен определёнными нуклеотидными последовательностями ДНК в реципиентном сайте.

Что отличает нас от животных? Поэт, романтик и философ скажут, что человека от животных отличает наличие души. Специалист в области высшей нервной деятельности вслед за Павловым заметит, что только у человека есть вторая сигнальная система. Нейробиолог добавит, рассказав нам много об особенностях строения мозга человека: и про увеличение ассоциативных полей неокортекса, и про увеличение височных долей, про центры речи и про многое другое. Антрополог, изучающий кости ископаемых предков Homo sapiens, обязательно скажет, что только у нас есть подбородочный выступ в черепе, нет надбровных дуг, несоразмерно большой мозговой отдел черепа и т.д. (Существует даже красивая теория, что одно из отличий человека от животных – это умение смеяться и плакать.) И все они будут правы. Выходит, искать отличия человека от животных можно в самых разных областях науки, да и жизни вообще. Однако, поскольку все признаки организма определяются его геномом, все эти отличия можно свести к отличиям в геноме. И поиск генетической основы, ответственной за очевидные различия в фенотипах человека и животных, особенно, его ближайших родственников, представляется одной из интереснейших задач современной биологии. Показано, что значительный вклад в структуру и функционирование генома, и, соответственно, в его видоспецифичность, вносят мобильные генетические элементы (МГЭ).
В обсуждаемой статье, посвященной роли МГЭ в функционировании генома человека, К. К. Баскаев и А. А. Буздин из Института биоорганической химии им. акад. М. М. Шемякина и Ю. А. Овчинникова РАН приводят краткий обзор основных генетических отличий человека от его ближайших нынеживущих родственников – шимпанзе, представленных в настоящее время двумя видами: обыкновенный шимпанзе (Pan troglodytes) и карликовый шимпанзе, или бонобо (P. paniscus). Авторы выделяют четыре основные группы генетических различий человека от шимпанзе.
1. Различная хромосомная организация, включающая утраты частей хромосом (делеции), различные вставки (инсерции), удвоения частей хромосом (дупликации) и обмен участками между негомологичными хромосомами (транслокации). Таких хромосомные перестройки происходят сравнительно часто, они имели место и на филогенетической линии, ведущей к человеку, и на линии шимпанзе. Посчитано, что инсерции и делеции, по которым человек отличается от шимпанзе, включают около 150 млн. п.о. Гораздо реже происходят более крупномасштабные события, такие как изменения числа хромосом. На человеческой линии произошло объединение двух предковых хромосом с образованием человеческой 2 хромосомы. У шимпанзе эта хромосома соответствует 12 и 13 хромосомам. Еще одно существенное отличие – перестройки в теломерных и центромерных областях, а так же в некоторых участках Y-хромосомы
2. Вариации в числе копий, положении в геноме и активности общих для человека и шимпанзе последовательностей.
3. Различия в белоккодирующих регионах, результатом которых являются различия в первичной структуре некоторых белков.
4. Видоспецифичные инсерции МГЭ.
Различия между ДНК человека и шимпанзе составляют в среднем около 1,24% и всего примерно 0,5% в белоккодирующих участках. Это значит, что если из всего генома человека наугад выбрать 100 нуклеотидов, то из них, скорее всего, только 1,24 будут не такими, как у шимпанзе. Тем не менее, никто не спутает человека с шимпанзе. Дело в том, что геном организован так, что один маленький эффект – экспрессия белоккодирующего гена - регулируется сложнейшей огромной сетью регуляторных генов. И маленькое изменение в этой пирамиде может радикальным образом увеличить или уменьшить экспрессию какого-то белка.
Особое внимание в обзоре было уделено анализу вклада специфических для человека МГЭ в структуру и функционирование генома человека.
Напомню о принципе строения, функционирования и вкладе МГЭ в геном. Тем, кто все это знает, следующие три абзаца можно пропустить. МГЭ – это фрагмент двунитевой молекулы ДНК, часть генома организма-хозяина, способная к самовоспроизведению, независимо от воспроизведения остального генома.

Рис. 2. МГЭ состоит из центральной части, фланкированной инвертированным повторами. Рисунок из лекций Зинченко В. В. по молекулярной генетики.
Попросту говоря, МГЭ могут перемещаться по геному, встраиваясь в определенные сайты встраивания молекул ДНК-мишеней и обеспечивая тем самым рекомбинацию между ДНК мобильного элемента и ДНК-мишенью. Элементарный МГЭ, или IS-элемент (от Insertion sequence, инсерционная последовательность) состоит из центральной части и концевых инвертированных повторов (рис. 2). Центральная часть кодирует белок транспозазу – ключевой белок транспозиции.
У эукариот, помимо описанных выше IS-элементов, существует два класса транспозонов: транспозоны I класса, присущие только эукариотам, и транспозоны II класса, общие для прокариот и эукариот. Транспозоны II класса, или ДНК-транспозоны, отличаются от описанных IS-элементов лишь тем, что они включает не только ген транспозазы, но и другие, посторонние, гены. Бывают составные ДНК-транспозоны и несоставные, или комплексные, ДНК-транспозоны. Составные ДНК-транспозоны представляют собой два идентичных типичных IS-элемента, окружающих центральную часть, содержащую посторонние гены, и вся эта конструкция вырезается, переносится и вшивается как единое целое. Комплексные ДНК-транспозоны по сути являются IS-элементами, отличие от типичных IS-элементов в том, что у этих транспозонов в центральной части присутствуют посторонние гены. Транспозоны II класса, или ретротранспозоны, присущи только эукариотам. И это, наряду с эндогенными ретровирусами (см. ниже), единственный способный к транспозиции класс МГЭ, найденный у млекопитающих и человека. МГЭ других классов утратили способность автономно воспроизводиться и перемещаться по геному из-за накопившихся мутаций. Главное отличие ретротранспозонов от ДНК-транспозонов в том, что их цикл репродукции включает стадию промежуточной молекулы РНК и стадию обратной транскрипции. Соответственно, в составе ретротранспозонов, как правило, имеется участок, кодирующий обратную транскриптазу, или ревертазу. На ретротранспозоны очень похожи т.н. эндогенные ретровирусы эукариот – ретровирусы, прочно интегрированные в геном клетки-хозяина и утратившие способность к образованию вирионов. Т.е. эндогенные ретровирусы можно рассматривать как МГЭ, имеющие вирусное происхождение, но на настоящей стадии являющиеся в большей степени частью организма-хозяина, чем частью вирусного паразита.
Результатом вставок МГЭ является отключение генов в результате непосредственной вставки МГЭ в их белоккодирующие части и изменение их активности в результате вставки МГЭ в регуляторные участки. Так, например, у человека не работает ген CMP, кодирующий фермент гидроксилазу сиаловой кислоты. Это фермент превращает N-ацетилнейраминовую кислоту в N-гликолилнейраминовую кислоту, которую человек, в отличие от шимпанзе, синтезировать не умеет. Причина отключения гена – вставка в его белоккодирующую часть специфического для человека ретротранспозона из семейства Alu. Другой результат транспозиции – это рекомбинация генома (рис. 4).


Рис. 4. Рекомбинация в результате вставки МГЭ. Слева направо: делеция, инверсия в результате внутримолекулярной рекомбинации; делеция + дупликация в результате рекомбинации между МГЭ гомологичных хромосом. Рисунок можно увеличить нажатием. Рисунок из лекций Зинченко В. В. по молекулярной генетики.
1. МГЭ – субстраты рекомбинации. Основные механизмы МГЭ-зависимой рекомбинации приведены на рис. 4.
2. МГЭ – энхансеры транскрипции. Действительно, известно много случаев, когда МГЭ увеличивает уровень экспрессии гена. Так, например, ретроэлемент семейства Alu является частью энхансерной области (т.е. области, повышающей активность) человеческого гена CD8.
3. МГЭ – транскрипционные промоторы. Оказывается, около 25% человеческих промоторов [0] (сайтов связывания ДНК-зависимой РНК-полимеразы) содержат в своей последовательности ретроэлементы.
4. МГЭ – поставщики альтернативных сайтов сплайсинга. Здесь ключевая роль принадлежит ретротранспозонам Alu. По краям белоккодирующего участка гена всегда есть нетранслируемые участки, в которых присутствие Alu может оказывать существенное влияние на то, что происходит с мРНК после транскрипции. Механизмы этого влияния могут быть разными: влияние на трансляцию, стабильность мРНК и альтернативный сплайсинг. Т.е. МГЭ могут влиять на продукт гена уже после его транскрипции. Примером такого влияния может служить специфическая для человека вставка Alu в ген, связанный с врождённой мышечной дистрофией.
5. МГЭ – доноры сигнала полиаденилирования. Напомню, что полиаденилирование – один из важных этапов созревания мРНК перед трансляцией, состоящий в присоединении 100-200 остатков аденина к определенной сигнальной последовательности – сигналу полиаденилирования. МГЭ человека кодируют свои гены, и эти гены имеют свои сигналы полиаденилирования. Значит, включение МГЭ в гены человека создает альтернативные сайты полиаденилирования.
6. МГЭ – антисмысловые регуляторы транскрипции . Антисмысловые РНК – это РНК, комплементарные мРНК. Показано, что МГЭ, присутствующие в интронах генов, как правило, находятся в антисмысловой ориентации относительно направления транскрипции этого гена. Таким образом, промоторы МГЭ могут управлять транскрипцией РНК, комплементарной участкам мРНК соответствующего гена. Получившаяся антисмысловая РНК, комплементарно связываясь с мРНК, может подавлять ее сплайсинг и трансляцию.
Читайте также: