
Мионевральный синапс это в анатомии кратко
Обновлено: 03.07.2024
Основным является сессионный cookie, обычно называемый MoodleSession. Вы должны разрешить использование этого файла cookie в своем браузере, чтобы обеспечить непрерывность и оставаться в системе при просмотре сайта. Когда вы выходите из системы или закрываете браузер, этот файл cookie уничтожается (в вашем браузере и на сервере).
Другой файл cookie предназначен исключительно для удобства, его обычно называют MOODLEID или аналогичным. Он просто запоминает ваше имя пользователя в браузере. Это означает, что когда вы возвращаетесь на этот сайт, поле имени пользователя на странице входа в систему уже заполнено для вас. Отказ от этого файла cookie безопасен - вам нужно будет просто вводить свое имя пользователя при каждом входе в систему.
Область контакта между двумя нейронами называют синапсом.
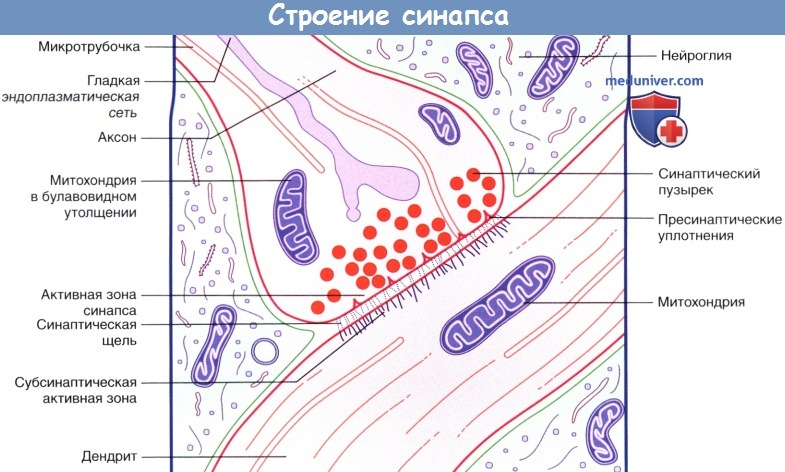
Внутреннее строение аксодендритического синапса.
а) Электрические синапсы. Электрические синапсы в нервной системе млекопитающих встречаются редко. Они образованы щелевидными контактами (нексусами) между дендритами или сомами соприкасающихся нейронов, которые соединяются с помощью цитоплазматических каналов диаметром 1,5 нм. Процесс передачи сигнала происходит без синаптической задержки и без участия медиаторов.
Посредством электрических синапсов возможно распространение электротонических потенциалов от одного нейрона к другому. Вследствие тесного синаптического контакта модуляция проведения сигнала невозможна. Задача этих синапсов — осуществление одновременного возбуждения нейронов, выполняющих одинаковую функцию. Примером служат нейроны дыхательного центра продолговатого мозга, которые во время вдоха синхронно генерируют импульсы. Кроме того, примером могут служить нейронные цепи, управляющие саккадами, при которых точка фиксации взора перемещается от одного объекта внимания к другому.
б) Химические синапсы. Большинство синапсов нервной системы — химические. Функционирование таких синапсов зависит от высвобождения медиаторов. Классический химический синапс представлен пресинаптической мембраной, синаптической щелью и постсинаптической мембраной. Пресинаптическая мембрана — часть булавовидного расширения нервного окончания клетки, передающей сигнал, а постсинаптическая мембрана — часть клетки, получающей сигнал.
Медиатор высвобождается из булавовидного расширения посредством экзоцитоза, проходит через синаптическую щель и связывается с рецепторами на постсинаптической мембране. Под постсинаптической мембраной расположена субсинаптическая активная зона, в которой после активации рецепторов постсинаптической мембраны происходят разнообразные биохимические процессы.
В булавовидном расширении расположены содержащие медиаторы синаптические пузырьки, а также большое количество митохондрий и цистерны гладкой эндоплазматической сети. Применение традиционных методик фиксации при исследовании клеток позволяет различить на пресинаптической мембране пресинаптические уплотнения, ограничивающие активные зоны синапса, к которым при помощи микротрубочек направляются синаптические пузырьки.
Аксодендритический синапс.
Срез препарата спинного мозга: синапс между концевым участком дендрита и, предположительно, двигательным нейроном.
Наличие округлых синаптических пузырьков и постсинаптического уплотнения характерно для возбуждающих синапсов.
Срез дендрита проведен в поперечном направлении, о чем свидетельствует наличие множества микротрубочек.
Кроме того, видны некоторые нейрофиламенты. Участок синапса окружен протоплазматическим астроцитом. Процессы, происходящие в нервных окончаниях двух типов.
(А) Синаптическая передача небольших молекул (например, глутамата).
(1) Транспортные пузырьки, содержащие мембранные белки синаптических пузырьков, направляются вдоль микротрубочек к плазматической мембране булавовидного утолщения.
В это же время происходит перенос молекул ферментов и глутамата путем медленного транспорта.
(2) Мембранные белки пузырьков выходят из плазматической мембраны и формируют синаптические пузырьки.
(3) Глутамат погружается в синаптические пузырьки; происходит накопление медиатора.
(4) Пузырьки, содержащие глутамат, подходят к пресинаптической мембране.
(5) В результате деполяризации происходит экзоцитоз медиатора из частично разрушенных пузырьков.
(6) Высвобождающийся медиатор распространяется диффузно в области синаптической щели и активирует специфические рецепторы на постсинаптической мембране.
(7) Мембраны синаптических пузырьков транспортируются обратно в клетку путем эндоцитоза.
(8) Происходит частичный обратный захват глутамата в клетку для повторного использования.
(Б) Передача нейропептидов (например, субстанции Р), осуществляющаяся одновременно с синаптической передачей (например, глутамата).
Совместная передача этих веществ происходит в центральных нервных окончаниях униполярных нейронов, обеспечивающих болевую чувствительность.
(1) Синтезированные в комплексе Гольджи (в области перикариона) пузырьки и предшественники пептидов (пропептиды) транспортируются к булавовидному расширению путем быстрого транспорта.
(2) При их попадании в область булавовидного утолщения завершается процесс формирования молекулы пептида, и пузырьки транспортируются к плазматической мембране.
(3) Деполяризация мембраны и перенос содержимого пузырьков в межклеточное пространство путем экзоцитоза.
(4) Одновременно с этим происходит высвобождение глутамата.
1. Активация рецепторов. Молекулы медиаторов проходят через синаптическую щель и активируют рецепторные белки, расположенные парами на постсинаптической мембране. Активация рецепторов запускает ионные процессы, которые приводят к деполяризации постсинаптической мембраны (возбуждающее постсинаптическое действие) или гиперполяризации постсинаптической мембраны (тормозящее постсинаптическое действие). Изменение электротонуса передается в сому в виде затухающего по мере распространения электротонического потенциала, за счет которого происходит изменение потенциала покоя в начальном сегменте аксона.
Ионные процессы подробно описаны в отдельной статье на сайте. При преобладании возбуждающих постсинаптических потенциалов начальный сегмент аксона деполяризуется до порогового уровня и генерирует потенциал действия.
Наиболее распространенный возбуждающий медиатор ЦНС — глутамат, а тормозной — гамма-аминомасляная кислота (ГАМК). В периферической нервной системе медиатором для двигательных нейронов поперечно-полосатой мускулатуры служит ацетилхолин, а для чувствительных нейронов — глутамат.
Последовательность процессов, происходящих в глутаматергических синапсах, показана на рисунке ниже. При передаче глутамата совместно с другими пептидами высвобождение пептидов осуществляется внесинаптическим путем.
Большинство чувствительных нейронов помимо глутамата выделяет и другие пептиды (один или несколько), высвобождающиеся в различных участках нейрона; однако основная функция этих пептидов — модуляция (повышение или снижение) эффективности синаптической передачи глутамата.
Кроме того, нейротрансмиссия может происходить путем диффузной внесинаптической передачи сигнала, характерной для моноаминергических нейронов (нейронов, использующих биогенные амины для обеспечения нейротрансмиссии). Выделяют две разновидности моноаминергических нейронов. В одних нейронах осуществляется синтез катехоламинов (норадреналина или дофамина) из аминокислоты тирозина, а в других — серотонина из аминокислоты триптофана. Например, дофамин высвобождается как в синаптической области, так и из варикозных утолщений аксона, в которых также происходит синтез этого нейромедиатора.
Дофамин проникает в межклеточную жидкость ЦНС и до момента деградации способен активировать специфические рецепторы на расстоянии до 100 мкм. Моноаминергические нейроны присутствуют во многих структурах ЦНС; нарушение передачи импульса этими нейронами приводит к различным заболеваниям, среди которых выделяют болезнь Паркинсона, шизофрению и глубокую депрессию.
Оксид азота (газообразная молекула) также участвует в диффузной нейропередаче в глутаматергической системе нейронов. Избыточное влияние оксида азота оказывает цитотоксическое действие, особенно в тех участках, кровоснабжение которых нарушено за счет тромбоза артерий. Глутамат также является потенциально цитотоксическим нейромедиатором.
Существует три вида химических межнейрональных взаимодействий: синаптическое (например, глутаматергическое), внесинаптическое (пептидергическое) и диффузное (например, моноаминергическое, серотонинергическое).
Химические синапсы классифицируют по анатомическому строению на аксодендритические, аксосоматические, аксоаксональные и дендро-дендритические. Синапс представлен пре- и постсинаптическими мембранами, синаптической щелью и субсинаптической активной зоной.
Электрические синапсы обеспечивают одновременную активацию целых групп нейронов, образуя между ними электрические связи за счет щелевидных контактов (нексусов).
Диффузная нейротрансмиссия в головном мозге.
Аксоны глутаматергического (1) и дофаминергического (2) нейронов образуют плотные синаптические контакты с отростком звездчатого нейрона (3) полосатого тела.
Дофамин высвобождается не только из пресинаптической области, но и из варикозного утолщения аксона, откуда диффузно распространяется в межклеточное пространство и активирует дофаминовые рецепторы дендритного ствола и стенки перицита капилляра. Растормаживание.
(А) Возбуждающий нейрон 1 активирует тормозной нейрон 2, который в свою очередь затормаживает нейрон 3.
(Б) Появление второго тормозного нейрона (2б) оказывает противоположное влияние на нейрон 3, поскольку происходит торможение нейрона 2б.
Спонтанно-активный нейрон 3 генерирует сигналы в условиях отсутствия тормозных влияний.
3. Торможение и растормаживание. Функционирование спонтанно-активных нейронов сдерживается под влиянием тормозных нейронов (обычно, ГАМКергических). Деятельность тормозных нейронов, в свою очередь, может быть ингибирована воздействующими на них другими тормозными нейронами, в результате чего происходит растормаживание клетки-мишени. Процесс растормаживания — важная особенность нейрональной активности в базальных ганглиях.
4. Редкие виды химических синапсов. Выделяют два типа аксоаксональных синапсов. В обоих случаях булавовидное утолщение образует тормозной нейрон. Синапсы первого типа образуются в области начального сегмента аксона и передают мощное ингибирующее влияние тормозного нейрона. Синапсы второго типа образуются между булавовидным утолщением тормозного нейрона и булавовидными утолщениями возбуждающих нейронов, что приводит к угнетению высвобождения медиаторов. Этот процесс получил название пресинаптического торможения. В этом плане традиционный синапс обеспечивает постсинаптичсекое торможение.
Дендро-дендритические (Д-Д) синапсы образуются между дендритными шипиками дендритов смежных шипиковых нейронов. Их задача — не генерирование нервного импульса, а изменение электротонуса клетки-мишени. В последовательных Д-Д-синапсах синаптические пузырьки располагаются только в одном дендритном шипике, а в реципрокном Д-Д-синапсе— в обоих. Возбуждающие Д-Д-синапсы изображены на рисунке ниже. Тормозные Д-Д-синапсы широко представлены в переключающих ядрах таламуса.
Кроме того, выделяют немногочисленные сомато-дендритические и сомато-соматические синапсы.
Аксоаксональные синапсы коры головного мозга.
Стрелками указано направление проведения импульсов. (1) Пресинаптическое и (2) постсинаптическое торможение спинномозгового нейрона, направляющегося к головному мозгу.
Стрелками указано направление проведения импульсов (возможно торможение переключательного нейрона под действием тормозных влияний). Возбуждающие дендро-дендритические синапсы. Изображены дендриты трех нейронов.
Реципрокный синапс (справа). Стрелками указано направление распространения электрото-нических волн.
Синапс-контакт между 2-умя возбудимыми клетками,цитоплазма каждой из которых окружена отдельной электрогенной мембраной.
Мионевральный (нервно-мышечный) синапс – образован аксоном мотонейрона и мышечной клеткой.
Нервный импульс возникает в тригерной зоне нейрона, по аксону направляется к иннервируемой мышце, достигает терминали аксона и при этом деполяризует пресинаптическую мембрану.
После этого открываются натриевые и кальциевые каналы, и ионы Ca из среды, окружающей синапс, входят внутрь терминали аксона. При этом процессе броуновское движение везикул упорядочивается по направления к пресинаптической мембране. Ионы Ca стимулируют движение везикул. Достигая пресинаптическую мембрану, везикулы разрываются, и освобождается ацетилхолин. Синаптическая щель заполнена жидкостью, которая по составу напоминает плазму крови, через нее происходит диффузия АХ с пресинаптической мембраны на постсинаптическую, но ее скорость очень мала. Кроме того, диффузия возможна еще и по фиброзным нитям, которые находятся в синаптической щели. После диффузии АХ начинает взаимодействовать с хеморецепторами (ХР) и холинэстеразой (ХЭ), которые находятся на постсинаптической мембране.
Холинорецептор выполняет рецепторную функцию, а холинэстераза выполняетферментативную функцию.
Затем происходит суммация МПКП. В результате суммации образуется ВПСП – возбуждающий постсинаптический потенциал. Постсинаптическая мембрана за счет ВПСП заряжается отрицательно, а на участке, где нет синапса (мышечного волокна), заряд положительный. Возникает разность потенциалов, образуется потенциал действия, который перемещается по проводящей системе мышечного волокна.
В состоянии относительного физиологического покоя синапс находятся в фоновой биоэлектрической активности. Ее значение заключается в том, что она повышает готовность синапса к проведению нервного импульса тем самым значительно облегчает передачу нервного возбуждения по синапсу.
Мионевральный синапс обладает следующими основными свойствами.
1. Синапс проводит возбуждение только в одном направлении – в направлении от пресинаптической мембраны к постсинаптической.
2. В синапсе имеет место синаптическая задержка возбуждения, т. е. скорость проведения возбуждения по синапсу значительно меньше, чем по нервному волокну. Это связано с определенной продолжительностью времени, необходимого для выделения медиатора и взаимодействия его с рецепторами.
3. В синапсе отмечается облегчение проведения каждого последующего возбуждения, что, по всей вероятности, связано с накоплением медиатора в синаптической щели.
4. При длительном возбуждении синапса в нем может наблюдаться снижение чувствительности рецепторов к медиатору, обусловленное закрытием части натриевых каналов, за счет включения системы инактивации.
5. В синапсах быстро развивается процесс утомления, связанный с быстрым метаболическим истощением запасов медиатора в везикулах пресинаптических утолщений.
Синапс-контакт между 2-умя возбудимыми клетками,цитоплазма каждой из которых окружена отдельной электрогенной мембраной.
Мионевральный (нервно-мышечный) синапс – образован аксоном мотонейрона и мышечной клеткой.
Нервный импульс возникает в тригерной зоне нейрона, по аксону направляется к иннервируемой мышце, достигает терминали аксона и при этом деполяризует пресинаптическую мембрану.
После этого открываются натриевые и кальциевые каналы, и ионы Ca из среды, окружающей синапс, входят внутрь терминали аксона. При этом процессе броуновское движение везикул упорядочивается по направления к пресинаптической мембране. Ионы Ca стимулируют движение везикул. Достигая пресинаптическую мембрану, везикулы разрываются, и освобождается ацетилхолин. Синаптическая щель заполнена жидкостью, которая по составу напоминает плазму крови, через нее происходит диффузия АХ с пресинаптической мембраны на постсинаптическую, но ее скорость очень мала. Кроме того, диффузия возможна еще и по фиброзным нитям, которые находятся в синаптической щели. После диффузии АХ начинает взаимодействовать с хеморецепторами (ХР) и холинэстеразой (ХЭ), которые находятся на постсинаптической мембране.

Холинорецептор выполняет рецепторную функцию, а холинэстераза выполняетферментативную функцию.
Затем происходит суммация МПКП. В результате суммации образуется ВПСП – возбуждающий постсинаптический потенциал. Постсинаптическая мембрана за счет ВПСП заряжается отрицательно, а на участке, где нет синапса (мышечного волокна), заряд положительный. Возникает разность потенциалов, образуется потенциал действия, который перемещается по проводящей системе мышечного волокна.
В состоянии относительного физиологического покоя синапс находятся в фоновой биоэлектрической активности. Ее значение заключается в том, что она повышает готовность синапса к проведению нервного импульса тем самым значительно облегчает передачу нервного возбуждения по синапсу.
Мионевральный синапс обладает следующими основными свойствами.
1. Синапс проводит возбуждение только в одном направлении – в направлении от пресинаптической мембраны к постсинаптической.
2. В синапсе имеет место синаптическая задержка возбуждения, т. е. скорость проведения возбуждения по синапсу значительно меньше, чем по нервному волокну. Это связано с определенной продолжительностью времени, необходимого для выделения медиатора и взаимодействия его с рецепторами.
3. В синапсе отмечается облегчение проведения каждого последующего возбуждения, что, по всей вероятности, связано с накоплением медиатора в синаптической щели.
4. При длительном возбуждении синапса в нем может наблюдаться снижение чувствительности рецепторов к медиатору, обусловленное закрытием части натриевых каналов, за счет включения системы инактивации.
5. В синапсах быстро развивается процесс утомления, связанный с быстрым метаболическим истощением запасов медиатора в везикулах пресинаптических утолщений.
Мионевральный (нервно-мышечный) синапс – образован аксоном мотонейрона и мышечной клеткой.
Нервный импульс возникает в тригерной зоне нейрона, по аксону направляется к иннервируемой мышце, достигает терминали аксона и при этом деполяризует пресинаптическую мембрану. После этого открываются натриевые и кальциевые каналы, и ионы Ca из среды, окружающей синапс, входят внутрь терминали аксона. При этом процессе броуновское движение везикул упорядочивается по направления к пресинаптической мембране. Ионы Ca стимулируют движение везикул. Достигая пресинаптическую мембрану, везикулы разрываются, и освобождается ацетилхолин (4 иона Ca высвобождают 1 квант ацетилхолина). Синаптическая щель заполнена жидкостью, которая по составу напоминает плазму крови, через нее происходит диффузия АХ с пресинаптической мембраны на постсинаптическую, но ее скорость очень мала. Кроме того, диффузия возможна еще и по фиброзным нитям, которые находятся в синаптической щели. После диффузии АХ начинает взаимодействовать с хеморецепторами (ХР) и холинэстеразой (ХЭ), которые находятся на постсинаптической мембране.
Холинорецептор выполняет рецепторную функцию, а холинэстераза выполняет ферментативную функцию. На постсинаптической мембране они расположены следующим образом:
ХР + АХ = МПКП – миниатюрные потенциалы концевой пластины.
Затем происходит суммация МПКП. В результате суммации образуется ВПСП – возбуждающий постсинаптический потенциал. Постсинаптическая мембрана за счет ВПСП заряжается отрицательно, а на участке, где нет синапса (мышечного волокна), заряд положительный. Возникает разность потенциалов, образуется потенциал действия, который перемещается по проводящей системе мышечного волокна.
ХЭ + АХ = разрушение АХ до холина и уксусной кислоты.
В состоянии относительного физиологического покоя синапс находятся в фоновой биоэлектрической активности. Ее значение заключается в том, что она повышает готовность синапса к проведению нервного импульса. В состоянии покоя 1–2 пузырька в терминале аксона могут случайно подойти к пресинаптической мембране, в результате чего вступят с ней в контакт. Везикула при контакте с пресинаптической мембраной лопается, и ее содержимое в виде 1 кванта АХ поступает в синаптическую щель, попадая при этом на постсинаптическую мембрану, где будет образовываться МПКН.
Данный текст является ознакомительным фрагментом.
Продолжение на ЛитРес
Правила передачи и хранения продуктов
Правила передачи и хранения продуктов Больным разрешается передавать, если не назначена диета: печенье, молоко, кефир, масло, сыр, яйца, фрукты, варенье.К передаче не допускаются скоропортящаяся колбаса, острые рыбные и мясные консервы, торты, пирожные, сельдь, рыба.
ГЛАВА 3. МЕХАНИЗМЫ И ПУТИ ПЕРЕДАЧИ ИНФЕКЦИИ
ГЛАВА 3. МЕХАНИЗМЫ И ПУТИ ПЕРЕДАЧИ ИНФЕКЦИИ Для каждой инфекционной болезни имеется свой путь передачи микроорганизмов, который сформировался в процессе эволюции и является основным способом сохранения возбудителя как вида.Существуют три фазы перехода возбудителя из
2. Механизмы проведения возбуждения по нервному волокну. Законы проведения возбуждения по нервному волокну
2. Механизмы проведения возбуждения по нервному волокну. Законы проведения возбуждения по нервному волокну Механизм проведения возбуждения по нервным волокнам зависит от их типа. Существуют два типа нервных волокон: миелиновые и безмиелиновые.Процессы метаболизма в
Этиология опухолей (на примере рака молочной железы)
Этиология опухолей (на примере рака молочной железы) Предшественницей раковой клетки в организме всегда является нормальная клетка какой-либо ткани. Факторы (агенты), способные вызвать превращение (трансформацию) нормальной клетки в опухолевую, называются
Перечень разрешенных для передачи продуктов
Дороги, которые ВИЧ выбирает (пути передачи инфекции)
Механизм передачи звуковых колебаний
Механизм передачи звуковых колебаний Звуковые колебания, воздействуя на систему слуховых косточек среднего уха, приводят к колебательным движениям мембраны овального окна, которая, прогибаясь, вызывает волнообразные перемещения перилимфы в вестибулярной и через
Кармический феномен передачи проблем и болезней детям
Антон Ноймайр Музыка и медицина. На примере немецкой романтики
ДУХОВНАЯ МАСТЕРСКАЯ. ЛИНИЯ ПЕРЕДАЧИ
ДУХОВНАЯ МАСТЕРСКАЯ. ЛИНИЯ ПЕРЕДАЧИ 1. Dr. Mikao Usui2. Dr. Chijiro Haiashi3. Mrs. Takata4. Phillis Lei Furumoto5. Carol Farmer6. Leath Smith7. William L. Rand (Usui Tibet Reiki)8. Hazel Raven9. Craham Raven10. Padma С. Т. Gara11. Mark O. Gara12. Inna Higham-Tchoumakova13. Valery N. Govyadinov14. Sergey Postankevich15. Igor
Принцип передачи усилия на кость
Принцип передачи усилия на кость Прямое воздействие на кость включает в себя два аспекта: физический и энергетический. (Деление на аспекты необходимо исключительно для педагогических целей.) К кости прикладывают заметное физическое усилие, приводящее к ее деформации.
Закон передачи
Закон передачи Из Нравственного Закона следуют и законы морали, и законы этики, и масса других законов. Сейчас мы с вами рассмотрим закон Передачи. Он говорит о том, что:Радуясь успехам и благополучию других людей, вы умножаете свои силы и процветание.Сострадая ошибкам и
Читайте также: