
История генетики микроорганизмов кратко
Обновлено: 11.05.2024
Генетика бактерий. Геном бактерии. Генотип бактериальной клетки. Фенотип бактериальной клетки. Генетический материал бактерий.
В наши дни приоритетным направлением естествознания можно считать молекулярную биологию. Она тесно связана с микробиологией и в известном смысле является её детищем, так как в качестве основных моделей использует бактерии и вирусы, а одно из основных направлений молекулярной биологии — молекулярная генетика — долгое время являлась не чем иным, как генетикой бактерий и бактериофагов.
Изучение генетики бактерий имеет также и несомненный прикладной интерес, например в плане установления механизмов передачи патогенных свойств и устойчивости к лекарственным препаратам.
Бактерии — удобная модель для генетических исследований. Их отличает: относительная простота строения генома, позволяющая выявлять мутанты с частотой 10 -9 и ниже; гаплоидность, исключающая явление доминантности; половая дифференциация в виде донорских и реципиентных клеток; наличие обособленных, и интегрированных фрагментов ДНК (плазмид, транспозонов и т.д.); лёгкость культивирования и возможность получения популяций, содержащих миллиарды микробных тел.
Как и у других организмов, совокупность генов бактериальной клетки — геном — определяет её свойства и признаки (генотип). Фенотип бактериальной клетки — результат взаимодействий между бактерией и окружающей средой — также контролирует геном (так как сами признаки закодированы в бактериальных генах).
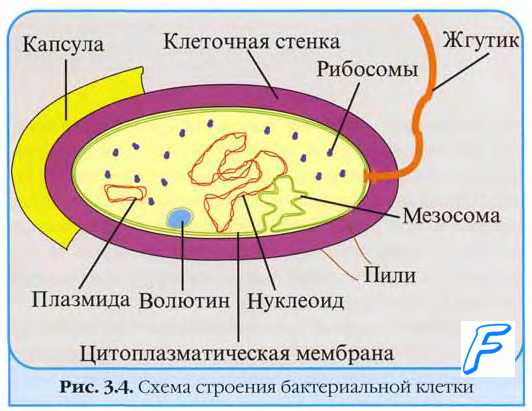
Генетический материал бактерий
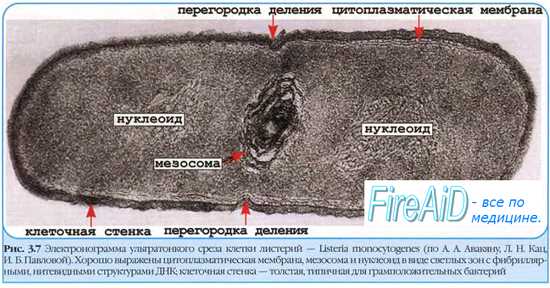
Ядерные структуры бактерий имеют характерное строение, отличающее их от ядер эукарио-тических клеток; их образуют так называемые хроматиновые тельца, или нуклеоиды, лишённые оболочки и включающие в себя почти всю ДНК бактерии.
• Ядерные структуры можно наблюдать в фазово-контрастный микроскоп, где они выглядят как менее плотные участки цитоплазмы. Для их выявления в фиксированных мазках предложена реакция Фёльгена-Россенбёка.
• В растущих бактериальных клетках нуклеоиды активно делятся, их количество иногда достигает 2-4.
Прокариотический геном
У бактерий обычно имеется одна замкнутая кольцевидная хромосома, содержащая до 4000 отдельных генов, необходимых для поддержания жизнедеятельности и размножения бактерий, то есть бактериальная клетка гаплоидна, а удвоение хромосомы обычно сопровождается её делением.
• Некоторые виды (например, Brucella melitensis) стабильно содержат две кольцевые хромосомы, другие (Leptospira interrogans) — одну кольцевую хромосому и одну большую плазмиду, третьи — одну линейную хромосому (Streptomyces ambofaciens), то есть обладают сложными геномами.
• Бактериальная хромосома содержит до 5*10 6 пар оснований. Для сравнения: геном человека составляет 2,9*10 9 пар оснований. Длина бактериальной хромосомы в развёрнутом состоянии составляет около 1 мм (Escherichia coli).
Некоторые бактерии содержат внехромосомные молекулы ДНК (плазмиды) и мобильные элементы (либо плазмидные, либо хромосомные).
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
В 40‑50‑е годы XX в. из общей микробиологии выделилось самостоятельное направление – генетика микроорганизмов – с наиболее важным разделом – генетикой бактерий. Основой исключительно быстрого развития этой области исследований послужили доказательства мутационной природы изменчивости микроорганизмов, обнаружение у них различных форм генетической рекомбинации и роли ДНК в их наследственных свойствах, достоинства микроорганизмов как объектов генетических исследований. Генетика микроорганизмов приобрела особое значение в связи с решением кардинальных проблем молекулярной биологии. Исследования структуры ДНК, ее роли в процессах биосинтеза белка и регуляции внутриклеточных обменных процессов, разработка проблем направленной изменчивости и специфичности мутагенеза ведутся в основном на микроорганизмах.

Георгий Адамович Надсон. 1867–1940.
До оформления генетики микроорганизмов в самостоятельную отрасль знания генетики и микробиологи работали разобщенно. Микробиологи были далеки от генетического истолкования наблюдавшихся ими фактов изменчивости и наследственности микробов, а генетики либо вообще не интересовались бактериологией, либо скептически относились к возможности применения генетических принципов к таким примитивно организованным формам жизни, какими им представлялись бактерии.
Первый этап в развитии генетики бактерий составили исследования, экспериментально доказывающие сходство природы и механизмов передачи потомству наследственных признаков у бактерий и высших организмов.
До выделения генетики бактерий в самостоятельную дисциплину существовало несколько точек зрения на популяционную изменчивость бактерий. Согласно одной из них, все изменения, вызванные внешней средой, являются непосредственно адаптивными. Это воззрение основывалось на представлении о наличии у бактерий повышенной врожденной пластичности, позволяющей им адекватно реагировать на разнообразные условия внешней среды. Длительному сохранению в микробиологии этого взгляда способствовал тот факт, что работа с бактериями велась, как правило, на больших популяциях, быстро размножающихся бесполым путем, что затрудняло контроль за изменчивостью отдельных особей. Вследствие этого изменения наследуемых признаков в единичных клетках оставались не обнаруженными до тех пор, пока такие измененные клетки не размножались в достаточно большом количестве, чтобы образовать целую популяцию.
Согласно другой точке зрения, изменчивость бактерий является результатом длительных модификаций, не затрагивающих наследственной основы организма и проявляющихся в течение многих поколений после того, как вызвавший их фактор перестал действовать. Третья точка зрения сводилась к признанию полицикличности в развитии бактерий, проявлением которой служат изменения их признаков. Эти взгляды означали реставрацию идеи полиморфизма, утверждавшей наличие в мире микробов безграничной изменчивости.
В период с 1910 по 1940 г. были описаны различные виды наследственной изменчивости. В 1921 г. французский микробиолог П. де Крюи описал изменение ряда свойств у одной из патогенных бактерий, возникающее, по его мнению, в результате расщепления признаков под влиянием неблагоприятных условий среды. Этот вид изменчивости он назвал диссоциацией. В 1925 г. в Советском Союзе Г.А. Надсон и Г.С. Филиппов впервые получили мутационные (по их терминологии, сальтационные) стойкие изменения признаков у дрожжевых и плесневых грибков, подвергнув их облучению рентгеновыми лучами. Это был первый случай индуцированной мутации, вызванной физическим мутагенным фактором. Первые данные о характере мутаций позволили провести параллель между природой изменчивости у бактерий и высших организмов. Это стало возможным также после того, как были разработаны методы, которые дали возможность отличать изменения отдельной клетки от изменчивости целой популяции.

Причины различий в объяснении фактов изменчивости у микробов носили преимущественно методологический характер. Из‑за отсутствия единой методики эксперимента данные разных авторов оставались несопоставимыми. Трудности в разграничении фенотипа и генотипа приводили зачастую к отрицанию различий между адаптацией и мутацией. Случаи морфологической изменчивости, требующие длительного наблюдения, нередко объясняли наличием сложных циклов развития. Длительное время отсутствовали и единые методы генетического анализа, в частности, принцип отбора мутантов.
Значительное упорядочение представлений о природе изменчивости у микробов было связано с разработкой методов генетического анализа у высших организмов и утверждением представлений о сходстве механизмов изменчивости и наследственности у всех живых организмов.
Генетика бактерии.
Важным событием в изучении природы мутаций было открытие биохимических (ауксотрофных) мутантов, т. е. микробов, потерявших в результате мутации способность самостоятельно синтезировать те или иные метаболиты и потому нуждающихся во включении этих метаболитов в питательную среду. Ауксотрофные мутанты были впервые обнаружены Дж. Бидлом и Э. Тейтумом в 1941 г. у плесневого грибка Neurospora crassa при облучении культуры ультрафиолетовыми лучами. Вслед за тем Дж. Ледерберг и Э. Тейтум (1946) обнаружили у отдельных мутантных клеток Escherichia coli утрату способности синтезировать некоторые аминокислоты и витамины.
Восстановление биосинтетической активности у ауксотрофов (или реверсия к прототрофности) было впервые описано Ф. Райяном и Дж. Ледербергом в 1948 г. у Neurospora crassa. Это явление рассматривалось как результат либо реверсной мутации в том же локусе, либо мутации в другом локусе хромосомы, сцепленном с локусом, затронутым прямой мутацией (супрессорная мутация). Последующее изучение явления обратного мутирования у биохимических мутантов позволило уточнить механизмы этого процесса. Было показано, что ревертирование обязано не истинным обратным мутациям, а главным образом супрессорным мутациям, возникающим в другом месте генома и приводящим к восстановлению дикого фенотипа. Важный материал был получен также при изучении генетических факторов, регулирующих обмен углеводов, а в связи с этим и механизмов, контролирующих образование ферментов, а также мутирование морфологических признаков клеток, антигенных и вирулентных свойств. Данные по изучению этих мутантов в значительной мере взаимосвязаны (П. де Крюи, 1921; А. Александрини, 1931; В. Браун 40‑е годы).
Для раскрытия молекулярной сущности мутагенеза как главного механизма изменения наследственной информации решающее значение имела расшифровка в 1953 г. структуры молекулы ДНК Дж. Уотсоном и Ф. Криком (Нобелевская премия, 1962) (см. главу 23). Это фундаментальное открытие заложило основу изучения механизмов передачи наследственной информации у бактерий с помощью методов молекулярной биологии. Речь идет об исследовании трансформации, трансдукции, конъюгации и лизогенной конверсии.
В 40‑50‑е годы XX в. из общей микробиологии выделилось самостоятельное направление – генетика микроорганизмов – с наиболее важным разделом – генетикой бактерий. Основой исключительно быстрого развития этой области исследований послужили доказательства мутационной природы изменчивости микроорганизмов, обнаружение у них различных форм генетической рекомбинации и роли ДНК в их наследственных свойствах, достоинства микроорганизмов как объектов генетических исследований. Генетика микроорганизмов приобрела особое значение в связи с решением кардинальных проблем молекулярной биологии. Исследования структуры ДНК, ее роли в процессах биосинтеза белка и регуляции внутриклеточных обменных процессов, разработка проблем направленной изменчивости и специфичности мутагенеза ведутся в основном на микроорганизмах.
Георгий Адамович Надсон. 1867–1940.
До оформления генетики микроорганизмов в самостоятельную отрасль знания генетики и микробиологи работали разобщенно. Микробиологи были далеки от генетического истолкования наблюдавшихся ими фактов изменчивости и наследственности микробов, а генетики либо вообще не интересовались бактериологией, либо скептически относились к возможности применения генетических принципов к таким примитивно организованным формам жизни, какими им представлялись бактерии.
Первый этап в развитии генетики бактерий составили исследования, экспериментально доказывающие сходство природы и механизмов передачи потомству наследственных признаков у бактерий и высших организмов.
До выделения генетики бактерий в самостоятельную дисциплину существовало несколько точек зрения на популяционную изменчивость бактерий. Согласно одной из них, все изменения, вызванные внешней средой, являются непосредственно адаптивными. Это воззрение основывалось на представлении о наличии у бактерий повышенной врожденной пластичности, позволяющей им адекватно реагировать на разнообразные условия внешней среды. Длительному сохранению в микробиологии этого взгляда способствовал тот факт, что работа с бактериями велась, как правило, на больших популяциях, быстро размножающихся бесполым путем, что затрудняло контроль за изменчивостью отдельных особей. Вследствие этого изменения наследуемых признаков в единичных клетках оставались не обнаруженными до тех пор, пока такие измененные клетки не размножались в достаточно большом количестве, чтобы образовать целую популяцию.
Согласно другой точке зрения, изменчивость бактерий является результатом длительных модификаций, не затрагивающих наследственной основы организма и проявляющихся в течение многих поколений после того, как вызвавший их фактор перестал действовать. Третья точка зрения сводилась к признанию полицикличности в развитии бактерий, проявлением которой служат изменения их признаков. Эти взгляды означали реставрацию идеи полиморфизма, утверждавшей наличие в мире микробов безграничной изменчивости.
В период с 1910 по 1940 г. были описаны различные виды наследственной изменчивости. В 1921 г. французский микробиолог П. де Крюи описал изменение ряда свойств у одной из патогенных бактерий, возникающее, по его мнению, в результате расщепления признаков под влиянием неблагоприятных условий среды. Этот вид изменчивости он назвал диссоциацией. В 1925 г. в Советском Союзе Г.А. Надсон и Г.С. Филиппов впервые получили мутационные (по их терминологии, сальтационные) стойкие изменения признаков у дрожжевых и плесневых грибков, подвергнув их облучению рентгеновыми лучами. Это был первый случай индуцированной мутации, вызванной физическим мутагенным фактором. Первые данные о характере мутаций позволили провести параллель между природой изменчивости у бактерий и высших организмов. Это стало возможным также после того, как были разработаны методы, которые дали возможность отличать изменения отдельной клетки от изменчивости целой популяции.
Причины различий в объяснении фактов изменчивости у микробов носили преимущественно методологический характер. Из‑за отсутствия единой методики эксперимента данные разных авторов оставались несопоставимыми. Трудности в разграничении фенотипа и генотипа приводили зачастую к отрицанию различий между адаптацией и мутацией. Случаи морфологической изменчивости, требующие длительного наблюдения, нередко объясняли наличием сложных циклов развития. Длительное время отсутствовали и единые методы генетического анализа, в частности, принцип отбора мутантов.
Значительное упорядочение представлений о природе изменчивости у микробов было связано с разработкой методов генетического анализа у высших организмов и утверждением представлений о сходстве механизмов изменчивости и наследственности у всех живых организмов.
Генетика бактерии.
Важным событием в изучении природы мутаций было открытие биохимических (ауксотрофных) мутантов, т. е. микробов, потерявших в результате мутации способность самостоятельно синтезировать те или иные метаболиты и потому нуждающихся во включении этих метаболитов в питательную среду. Ауксотрофные мутанты были впервые обнаружены Дж. Бидлом и Э. Тейтумом в 1941 г. у плесневого грибка Neurospora crassa при облучении культуры ультрафиолетовыми лучами. Вслед за тем Дж. Ледерберг и Э. Тейтум (1946) обнаружили у отдельных мутантных клеток Escherichia coli утрату способности синтезировать некоторые аминокислоты и витамины.
Восстановление биосинтетической активности у ауксотрофов (или реверсия к прототрофности) было впервые описано Ф. Райяном и Дж. Ледербергом в 1948 г. у Neurospora crassa. Это явление рассматривалось как результат либо реверсной мутации в том же локусе, либо мутации в другом локусе хромосомы, сцепленном с локусом, затронутым прямой мутацией (супрессорная мутация). Последующее изучение явления обратного мутирования у биохимических мутантов позволило уточнить механизмы этого процесса. Было показано, что ревертирование обязано не истинным обратным мутациям, а главным образом супрессорным мутациям, возникающим в другом месте генома и приводящим к восстановлению дикого фенотипа. Важный материал был получен также при изучении генетических факторов, регулирующих обмен углеводов, а в связи с этим и механизмов, контролирующих образование ферментов, а также мутирование морфологических признаков клеток, антигенных и вирулентных свойств. Данные по изучению этих мутантов в значительной мере взаимосвязаны (П. де Крюи, 1921; А. Александрини, 1931; В. Браун 40‑е годы).
Для раскрытия молекулярной сущности мутагенеза как главного механизма изменения наследственной информации решающее значение имела расшифровка в 1953 г. структуры молекулы ДНК Дж. Уотсоном и Ф. Криком (Нобелевская премия, 1962) (см. главу 23). Это фундаментальное открытие заложило основу изучения механизмов передачи наследственной информации у бактерий с помощью методов молекулярной биологии. Речь идет об исследовании трансформации, трансдукции, конъюгации и лизогенной конверсии.
До определенного времени ученые полагали, что у микроорганизмов нет ядерного аппарата. Такое мнение бытовало приблизительно до 30-х 20 века. По этой причине не изучались и вопросы наследственности и изменчивости микроорганизмов.
Изобретение электронного микроскопа стало поворотной точкой в изучении микроорганизмов. А в целом появление этого инструмента позволило увидеть субмикроскопическую структуру клетки.
Начало 40-х годов связано с тем, что ученые-генетики обратили пристальное внимание на микроорганизмы. Полноценными объектами микроскопических исследований стали бактерии, вирусы и микроскопические грибы. Появляется даже целая отрасль микробиологии, которая получила название генетики микроорганизмов.
Генетика микроорганизмов представляет собой раздел общей генетики, предмет изучения которой — разнообразные микроорганизмы, такие как вирусы, бактерии и микроскопические грибы, а также особенности их наследственности и изменчивости.
Отличительная характеристика микроорганизмов — наличие гаплоидного набора хромосом или кольцевая молекула ДНК. За счет этого обеспечивается возможность мутаций уже в первом поколении потомков.
Начало микробиологических генетических исследований
Изучение субмикроскопической структуры клеток микроорганизмов позволило решить множество проблем генетики. Первым доказательством, что молекула ДНК является материальным носителем наследственности, стали результаты опытов американских ученых-генетиков О. Т. Эйвери, К. Мак-Леод и М. Маккарти. Они проводили опыты на пневмококках, а в частности на хлебной плесени. Ученые сформулировали положение, что синтез одной полипептидной цепи (одного белка) программирует один ген.
Новый всплеск изучения микроорганизмов с точки зрения генетики случился после доказательства универсальности закономерностей мутационного процесса на примере кишечной палочки. Этим занимались американские микробиологи С. Лурия и М. Дельброк. Они сделали вывод, что бактерии подчиняются мутационным закономерностям.
Все это поспособствовало появлению нового принципа изучения изменчивости у бактерий.
Это принцип получил название клонального анализа. В его основе лежит тщательное исследование потомства одной клетки, которая становится родоначальницей клона.
Изучение бактерий
Большие исследования провели американские генетики Джошуа и Эстер Ледерберги. Они доказали, что мутации у бактерий возникают вне зависимости от условий их культивирования.
Также учеными был разработан метод отпечатков, позволяющий сильно упростить приемы отбора микроорганизмов с необходимыми свойствами для последующих изучений. В больших популяциях клеток бактерий наблюдаются неупорядоченные (спонтанные) мутации — к такому выводу пришли ученые.
В 1946 году появилось доказательство того, что бактериям свойственен половой процесс. Также были открыты такие явления как конъюгация хромосом и рекомбинация генов, перенос генетической информации от одной бактериальной клетки к другой с помощью бактериофага.
Синтез того или иного белка зависит от того, с какого нуклеотида начался этот процесс считывания.
Изучение фагов
В процессе изучения особенностей взаимоотношений между бактериями и бактериофагами, американским ученым удалось открыть трансдукцию и рекомбинацию у фагов.
Под трансдукцией понимают перенос генов между бактериальными клетками с помощью фагов.
Все перечисленное выше позволило заниматься изучением вопросом наследственности на уровне молекул — т. н. молекулярный уровень организации материи.
Немецкие микробиологи изучали молекулу РНК. Каждая группа микроорганизмов получила свою методику исследования.
Генетика грибов и водорослей
Половой процесс низших грибов и водорослей отличается от полового процесса прочих организмов. Изучение этих групп организмов привело к разработке нового метода — тетрадного анализа.
В ходе исследования этих организмов ученые занимались разработкой методики объединения ядер генетически различных штаммов микроорганизмов.
В дальнейшем с помощью этих методов ученые смогут:
- вывести организмы с определенными качествами;
- разработать новые поколения антибиотиков и биологически активных веществ;
- бороться с многочисленными заболеваниями растений, животных и людей.
Тем не менее генная инженерия требует к себе основательного и осторожного подхода. Пока еще не совсем понятно, какие могут появиться в природе и в человеческом организме генетически модифицированные организмы.
Генетика микроорганизмов как наука, специфика исследований в микробиологии
Наследственностью называется свойство организмов передавать потомкам особенности строения, физиологические свойства и характер индивидуального развития.
Наследственностью люди интересовались очень давно. С развитием сельского хозяйства сформировалась прикладная наука селекция, которая занималась созданием и формированием новых пород животных и сортов растений. Но объяснить механизмы передачи признаков потомкам селекционеры не могли.
Г. Мендель установил дискретность (отдельность) наследственных факторов и разработал гибридологический метод изучения наследственности.
Дискретность наследственности состоит в том, что отдельные свойства и признаки организма развиваются под контролем наследственных факторов, которые при слиянии гамет и образовании зиготы не смешиваются, а при формировании новых гамет наследуются независимо друг от друга.
Значение открытий Г. Менделя оценили только после того, как его результаты были подтверждены в \(1900\) г. тремя биологами независимо друг от друга: Х. де Фризом в Голландии, К. Корренсом в Германии и Э. Чермаком в Австрии. Этот год считается годом возникновения науки генетики.
Менделевские законы наследственности заложили основу теории гена, а генетика превратилась в быстро развивающуюся отрасль биологии.
В \(1901\)–\(1903\) гг. де Фриз выдвинул мутационную теорию изменчивости, которая сыграла большую роль в дальнейшем развитии генетики.
Второй этап развития генетики — изучение закономерностей наследования признаков на хромосомном уровне.
Была установлена взаимосвязь между менделевскими законами наследования и распределением хромосом в процессе клеточного деления (митоз) и созревания половых клеток (мейоз).
Изучение строения клетки привело к уточнению строения, формы и количества хромосом и помогло установить, что гены — это участки хромосом.
В \(1910\)–\(1911\) гг. американский генетик Т. Г. Морган и его сотрудники провели исследования закономерностей наследования на мушках дрозофилах. Они установили, что гены расположены в хромосомах в линейном порядке и образуют группы сцепления.
В \(1953\) г. Ф. Крик и Дж. Уотсон создали модель молекулы ДНК в виде двойной спирали и объяснили способность ДНК к самоудвоению. Стал понятен механизм изменчивости: любые отклонения в структуре гена, однажды возникнув, в дальнейшем воспроизводятся в дочерних нитях ДНК.
Эти положения были подтверждены экспериментами. Уточнилось понятие гена, был расшифрован генетический код и изучен механизм биосинтеза. Были разработаны методы искусственного получения мутаций и с их помощью созданы новые ценные сорта растений и штаммы микроорганизмов.
В последние десятилетия сформировалась генная инженерия — система приёмов, позволяющих синтезировать новый ген или выделить его из одного организма и ввести в генетический аппарат другого организма.
В последнее десятилетие \(20\) века были расшифрованы геномы многих простых организмов. В начала \(21\) века (\(2003\) г.) был завершён проект по расшифровке генома человека.
На сегодняшний день существуют базы данных геномов многих организмов. Наличие такой базы данных человека имеет большое значение в предупреждении и исследовании многих заболеваний.
Читайте также: