
Фотосистема 1 и 2 кратко
Обновлено: 02.07.2024
Фотосистема — совокупность ССК, фотохимического реакционного центра и переносчиков электрона. В хлоропластах зеленых растений имеются две фотоактивные пигментные системы: фотосистема I(ФС-1) и фотосистема II (ФС-2). Открыты они были в связи си изучением эффекта существенного усиления фотосинтетической активности хлоропластов при добавлении к дальнему красному свету более коротковолнового. Пониженная интенсивность фотосинтеза на одном лишь длинноволновом свете объясняется тем, что в возбужденное состояние приходят молекулы хлорофилла ФС-1, добавление же более коротковолнового света возбуждает обе системы, что и приводит к резкому усилению фотосинтеза.
ФС-2 содержит 200 молекул хлорофилла a, 100 молекул хлорофилла b, 50 молекул каротиноидов и 2 молекулы феофитина. Реакционный центр ФС-2 представляет собой пигмент-белковый комплекс, расположенный в тилакоидных мембранах и окружённый ССК. В нём находится димер хлорофиллаaс максимумом поглощения при 680 нм (П680). На него,в конечном счёте передаётся энергия кванта света из ССК, в результате чего один из электронов переходит на более высокое энергетическое состояние, связь его с ядром ослабляется и возбуждённая молекула П680становится сильным восстановителем (E0=-0,7 В).
П680 восстанавливает феофитин, в дальнейшем электрон переносится на хиноны, входящие в состав ФС-2 и далее на пластохиноны, транспортируемые в восстановленной форме к b6f комплексу. Одна молекула пластохинона переносит 2 электрона и 2 протона, которые берутся из стромы. Заполнение электронной вакансии в молекуле П680происходит за счёт воды. В состав ФС-2 входит водоокисляющий комплекс, содержащий в активном центре ионы марганца в количестве 4 штук. Для образования одной молекулы кислорода требуется две молекулы воды, дающие 4 электрона. Поэтому процесс проводится в 4 такта и для его полного осуществления требуется 4 кванта света. Комплекс находится со стороны внутри тилакоидного пространства и полученные 4 протона выбрасываются в него.
Таким образом, суммарный результат работы ФС-2 —это окисление 2 молекул воды с помощью 4 квантов света с образованием 4 протонов во внутри тилакоидном пространстве и 2 восстановленных пластохинонов в мембране
В эволюционном отношении ФС-1 более древняя фотосистема. У фотосинтезирующих бактерий эта система не способна использовать воду в качестве донора электронов и является единственной.ФС-1содержит примерно 200 молекул хлорофилла.В реакционном центре первой фотосистемы находится димер хлорофилла a с максимумом поглощения при 700 нм (П700). После возбуждения квантом света он восстанавливает первичный акцептор —хлорофилл a, тот —вторичный (витамин K1или филлохинон), после чего электрон передаётся на ферредоксин, который и восстанавливает НАДФ с помощью фермента ферредоксин-НАДФ-редуктазы. Белок пластоцианин, восстановленный в b6f комплексе, транспортируется к реакционному центру первой фотосистемы со стороны внутритилакоидного пространства и передаёт электрон на окисленный П700
Фотосистема — совокупность ССК, фотохимического реакционного центра и переносчиков электрона. В хлоропластах зеленых растений имеются две фотоактивные пигментные системы: фотосистема I(ФС-1) и фотосистема II (ФС-2). Открыты они были в связи си изучением эффекта существенного усиления фотосинтетической активности хлоропластов при добавлении к дальнему красному свету более коротковолнового. Пониженная интенсивность фотосинтеза на одном лишь длинноволновом свете объясняется тем, что в возбужденное состояние приходят молекулы хлорофилла ФС-1, добавление же более коротковолнового света возбуждает обе системы, что и приводит к резкому усилению фотосинтеза.
ФС-2 содержит 200 молекул хлорофилла a, 100 молекул хлорофилла b, 50 молекул каротиноидов и 2 молекулы феофитина. Реакционный центр ФС-2 представляет собой пигмент-белковый комплекс, расположенный в тилакоидных мембранах и окружённый ССК. В нём находится димер хлорофиллаaс максимумом поглощения при 680 нм (П680). На него,в конечном счёте передаётся энергия кванта света из ССК, в результате чего один из электронов переходит на более высокое энергетическое состояние, связь его с ядром ослабляется и возбуждённая молекула П680становится сильным восстановителем (E0=-0,7 В).

П680 восстанавливает феофитин, в дальнейшем электрон переносится на хиноны, входящие в состав ФС-2 и далее на пластохиноны, транспортируемые в восстановленной форме к b6f комплексу. Одна молекула пластохинона переносит 2 электрона и 2 протона, которые берутся из стромы. Заполнение электронной вакансии в молекуле П680происходит за счёт воды. В состав ФС-2 входит водоокисляющий комплекс, содержащий в активном центре ионы марганца в количестве 4 штук. Для образования одной молекулы кислорода требуется две молекулы воды, дающие 4 электрона. Поэтому процесс проводится в 4 такта и для его полного осуществления требуется 4 кванта света. Комплекс находится со стороны внутри тилакоидного пространства и полученные 4 протона выбрасываются в него.
Таким образом, суммарный результат работы ФС-2 —это окисление 2 молекул воды с помощью 4 квантов света с образованием 4 протонов во внутри тилакоидном пространстве и 2 восстановленных пластохинонов в мембране
В эволюционном отношении ФС-1 более древняя фотосистема. У фотосинтезирующих бактерий эта система не способна использовать воду в качестве донора электронов и является единственной.ФС-1содержит примерно 200 молекул хлорофилла.В реакционном центре первой фотосистемы находится димер хлорофилла a с максимумом поглощения при 700 нм (П700). После возбуждения квантом света он восстанавливает первичный акцептор —хлорофилл a, тот —вторичный (витамин K1или филлохинон), после чего электрон передаётся на ферредоксин, который и восстанавливает НАДФ с помощью фермента ферредоксин-НАДФ-редуктазы. Белок пластоцианин, восстановленный в b6f комплексе, транспортируется к реакционному центру первой фотосистемы со стороны внутритилакоидного пространства и передаёт электрон на окисленный П700
В фотосистемы они являются функциональными единицами фотосинтетического процесса. Они определяются формами их ассоциации и особой организацией фотосинтетических пигментов и белковых комплексов, способных поглощать и преобразовывать световую энергию в процессе, который включает в себя перенос электронов.
Известны два типа фотосистем, названные фотосистемами I и II из-за порядка, в котором они были обнаружены. Фотосистема I имеет очень большое количество хлорофилла. к по сравнению с количеством хлорофилла б, в то время как фотосистема II имеет очень похожее количество обоих фотосинтетических пигментов.
Фотосистемы расположены в тилакоидных мембранах фотосинтезирующих организмов, таких как растения и водоросли. Их также можно найти у цианобактерий.
Хлоропласты
Хлоропласты представляют собой сферические или удлиненные органеллы диаметром около 5 мкм, содержащие фотосинтетические пигменты. Внутри него в клетках растений происходит фотосинтез.
Они окружены двумя внешними мембранами, а внутри содержат мешкообразные структуры, также окруженные двумя мембранами, называемыми тилакоидами.
Тилакоиды уложены друг на друга, образуя набор, который называется грана, а жидкость, которая окружает тилакоиды, называется стромой.Кроме того, тилакоиды окружены мембраной, называемой просветом, которая ограничивает внутритилакоидное пространство.
Преобразование световой энергии в химическую энергию во время фотосинтеза происходит внутри мембран тилакоидов. С другой стороны, производство и хранение углеводов в результате фотосинтеза происходит в стромах.
Фотосинтетические пигменты
Это белки, способные поглощать световую энергию для использования ее в процессе фотосинтеза, они полностью или частично связаны с тилакоидной мембраной. Пигмент, непосредственно участвующий в световых реакциях фотосинтеза, - хлорофилл.
В растениях есть два основных типа хлорофилла, называемые хлорофиллами. к Y б.Однако у некоторых водорослей другие типы хлорофилла, такие как c и d, последний присутствует только в некоторых красных водорослях.
Есть и другие фотосинтетические пигменты, такие как каротины и ксантофиллы, которые вместе составляют каротиноиды. Эти пигменты представляют собой изопреноиды, обычно состоящие из сорока атомов углерода. Каротины - это не кислородсодержащие каротиноиды, а ксантофиллы - это кислородсодержащие пигменты.
В растениях только хлорофилл к он принимает непосредственное участие в световых реакциях. Остальные пигменты не поглощают непосредственно световую энергию, а действуют как вспомогательные пигменты, передавая энергию, захваченную от света, к хлорофиллу. к. Таким образом улавливается больше энергии, чем может уловить хлорофилл. к Сам по себе.
Фотосинтез
Фотосинтез - это биологический процесс, который позволяет растениям, водорослям и некоторым бактериям использовать энергию солнечного света. Благодаря этому процессу растения используют световую энергию для преобразования атмосферного углекислого газа и воды, получаемой из земли, в глюкозу и кислород.
Свет вызывает сложную серию реакций окисления и восстановления, которые позволяют преобразовывать световую энергию в химическую энергию, необходимую для завершения процесса фотосинтеза. Фотосистемы являются функциональными единицами этого процесса.
Компоненты фотосистем
Антенный комплекс
Он состоит из большого количества пигментов, включая сотни молекул хлорофилла. к и даже большее количество дополнительных пигментов, а также фикобилинов. Сложная антенна позволяет поглощать большое количество энергии.
Он работает как воронка или как антенна (отсюда и ее название), которая улавливает энергию солнца и преобразует ее в химическую энергию, которая передается в реакционный центр.
Благодаря передаче энергии молекула хлорофилла к он получает от реакционного центра гораздо больше световой энергии, чем мог бы получить сам по себе. Кроме того, если молекула хлорофилла получает слишком много света, она может фотоокисляться, и растение погибнет.
Центр реакции
Это комплекс, образованный молекулами хлорофилла. к, молекула, известная как первичный электронный рецептор, и окружающие ее многочисленные белковые субъединицы.
Функционирование
Обычно молекула хлорофилла к присутствующий в реакционном центре, который инициирует световые реакции фотосинтеза, не принимает фотоны напрямую. Вспомогательные пигменты, а также некоторые молекулы хлорофилла к присутствующие в сложной антенне получают световую энергию, но не используют ее напрямую.
Эта энергия, поглощенная антенным комплексом, передается хлорофиллу. к из реакционного центра. Каждый раз, когда активируется молекула хлорофилла к, он высвобождает заряженный электрон, который затем поглощается первичным электронным рецептором.
Как следствие, первичный акцептор восстанавливается, а хлорофилл к восстанавливает свой электрон благодаря воде, которая действует как последний освободитель электронов, а кислород получается как побочный продукт.
Типы
Фотосистема I
Он находится на внешней поверхности тилакоидной мембраны и имеет небольшое количество хлорофилла. б, помимо хлорофилла к и каротиноиды.
Хлорофилл к реакционного центра лучше поглощает волны длиной 700 нанометров (нм), поэтому его называют P700 (пигмент 700).
В фотосистеме I группа белков ферродоксиновой группы - сульфид железа - действует как конечные акцепторы электронов.
Фотосистема II
Сначала он действует в процессе преобразования света в фотосинтез, но был открыт после первой фотосистемы. Он находится на внутренней поверхности тилакоидной мембраны и имеет большее количество хлорофилла. б чем фотосистема I. Также содержит хлорофилл к, фикобилины и ксантофиллы.
В этом случае хлорофилл к реакционного центра лучше поглощает длину волны 680 нм (P680), а не 700 нм, как в предыдущем случае. Конечным акцептором электронов в этой фотосистеме является хинон.
Связь между фотосистемами I и II
Фотосинтетический процесс требует обеих фотосистем. Первая фотосистема, которая действует, - это II, которая поглощает свет, поэтому электроны в хлорофилле реакционного центра возбуждаются, а первичные акцепторы электронов захватывают их.
Электроны, возбужденные светом, перемещаются в фотосистему I через цепь переноса электронов, расположенную в тилакоидной мембране. Это смещение вызывает падение энергии, которое позволяет переносить ионы водорода (H +) через мембрану к просвету тилакоидов.
Транспорт ионов водорода обеспечивает разность энергий между просветом тилакоидов и стромой хлоропласта, которая служит для генерации АТФ.
Хлорофилл в реакционном центре фотосистемы I получает электрон, исходящий от фотосистемы II. Электрон может продолжать циклический перенос электронов вокруг фотосистемы I или использоваться для образования НАДФН, который затем переносится в цикл Кальвина.

Два основных мультисубъединичных мембранных белковых комплекса отличаются по своей поглощающей длине волны, где фотосистема I или PS 1 поглощает большую длину волны света, которая составляет 700 нм, в то время как фотосистема II или PS 2 поглощает более короткую длину волны света 680 нм .
Во-вторых, каждая фотосистема пополняется электронами после потери электрона, но источники отличаются, когда PS II получает электроны из воды, в то время как PS I получает электроны от PS II через цепь переноса электронов.
Фотосистемы участвуют в фотосинтезе и обнаруживаются в тилакоидных мембранах водорослей, цианобактерий и главным образом в растениях. Все мы знаем, что растения и другие фотосинтезирующие организмы накапливают солнечную энергию, которая поддерживается молекулами пигмента, поглощающими свет, присутствующими в листьях.
Поглощенная солнечная энергия или энергия света в листьях преобразуется в химическую энергию на первом этапе фотосинтеза. Этот процесс проходит ряд химических реакций, известных как светозависимые реакции.
Фотосинтетические пигменты, такие как хлорофилл а, хлорофилл b и каротиноиды, присутствуют в тилакоидных мембранах хлоропласта. Фотосистема представляет собой светосборные комплексы, состоящие из 300-400 хлорофиллов, белков и других пигментов. Эти пигменты возбуждаются после поглощения фотона, и затем один из электронов переключается на орбиту с более высокой энергией.
Возбужденный пигмент передает свою энергию соседнему пигменту посредством передачи резонансной энергии, и это прямые электромагнитные взаимодействия. Далее, в свою очередь, соседний пигмент передает энергию пигменту, и процесс повторяется многократно. Вместе эти молекулы пигмента собирают свою энергию и проходят к центральной части фотосистемы, известной как реакционный центр.
Хотя две фотосистемы в реакциях, зависящих от света, получили свое название в серии, они были обнаружены, но фотосистема II (PS II) сначала идет по пути в потоке электронов, а затем фотосистема I (PSI). В этом материале мы рассмотрим разницу между двумя типами фотосистемы и кратко их описание.
Сравнительная таблица
| Основа для сравнения | Фотосистема I (PS I) | Фотосистема II (PS II) |
|---|---|---|
| Смысл | Фотосистема I или PS I использует энергию света для преобразования NADP + в NADPH2. Он включает P700, хлорофилл и другие пигменты. | Фотосистема II или PS II представляет собой белковый комплекс, который поглощает световую энергию, включая P680, хлорофилл и вспомогательные пигменты, и переносит электроны из воды в пластохинон и, таким образом, работает в диссоциации молекул воды и производит протоны (H +) и O2. |
| Место расположения | Он расположен на внешней поверхности тилакоидной мембраны. | Он расположен на внутренней поверхности тилакоидной мембраны. |
| Фотоцентр или реакционный центр | P700 это фотоцентр. | P680 - фотоцентр. |
| Поглощающая длина волны | Пигменты в фотосистеме 1 поглощают свет с большей длиной волны, который составляет 700 нм (P700). | Пигменты в фотосистеме2 поглощают свет с более короткими длинами волн, который составляет 680 нм (P680). |
| фотофосфорилирования | Эта система участвует как в циклическом, так и в нециклическом фотофосфорилировании. | Эта система участвует как в циклическом фотофосфорилировании. |
| фотолиз | Фотолиза не происходит. | Фотолиз происходит в этой системе. |
| Пигменты | Фотосистема I или PS 1 содержит хлорофилл A-670, хлорофилл A-680, хлорофилл A-695, хлорофилл A-700, хлорофилл B и каротиноиды. | Фотосистема II или PS 2 содержит хлорофилл A-660, хлорофилл A-670, хлорофилл A-680, хлорофилл A-695, хлорофилл A-700, хлорофилл B, ксантофиллы и фикобилины. |
| Соотношение хлорофилловых каротиноидных пигментов | 20-30: 1. | 3-7: 1. |
| функция | Основная функция фотосистемы I заключается в синтезе NADPH, где она получает электроны от PS II. | Основная функция фотосистемы II заключается в гидролизе воды и синтезе АТФ. |
| Основной состав | PSI состоит из двух субъединиц, которые являются psaA и psaB. | PS II состоит из двух субъединиц, состоящих из D1 и D2. |
Определение Фотосистемы I
Фотосистема I или PSI расположена в тилакоидной мембране и представляет собой многосубъединичный белковый комплекс, найденный в зеленых растениях и водорослях. Первый начальный этап улавливания солнечной энергии и последующего преобразования посредством электронного транспорта, управляемого светом. PS I - это система, где хлорофилл и другие пигменты собираются и поглощают длину волны света при 700 нм. Это последовательность реакции, а реакционный центр состоит из хлорофилла А-700 с двумя субъединицами, а именно psaA и psaB.
Субъединицы PSI больше, чем субъединицы PS II. Эта система также состоит из хлорофилла А-670, хлорофилла А-680, хлорофилла А-695, хлорофилла В и каротиноидов. Поглощенные фотоны переносятся в реакционный центр с помощью вспомогательных пигментов. Фотоны далее высвобождаются реакционным центром в виде электронов высокой энергии, которые подвергаются ряду электронных носителей и, наконец, используются НАДФ + редуктазой. NADPH производится с помощью фермента NADP + редуктазы из электронов с такой высокой энергией. NADPH используется в цикле Кальвина.
Таким образом, основная цель комплексного мембранного белкового комплекса, который использует энергию света для производства АТФ и НАДФН. Фотосистема I также известна как пластоцианин-ферредоксин-оксидоредуктаза.
Определение Фотосистемы II
Фотосистема II или PS II представляет собой мембранно-внедренный белковый комплекс, состоящий из более чем 20 субъединиц и около 100 кофакторов. Свет поглощается пигментами, такими как каротиноиды, хлорофилл и фикобилин, в области, известной как антенны, и в дальнейшем эта возбужденная энергия передается в реакционный центр. Основным компонентом являются периферические антенны, которые поглощают свет вместе с хлорофиллом и другими пигментами. Эта реакция проводится в ядро комплекса, который является местом для начальных цепных реакций переноса электронов.
Как обсуждалось ранее, PS II поглощает свет при 680 нм и входит в высокоэнергетическое состояние. P680 жертвует электрон и передает его феофитину, который является основным акцептором электронов. Как только P680 теряет электрон и получает положительный заряд, он нуждается в электроне для пополнения, которое выполняется путем расщепления молекул воды.
Окисление воды происходит в центре марганца или кластере Mn4OxCa . Марганцевый центр окисляет две молекулы одновременно, выделяя четыре электрона и, таким образом, образуя молекулу O2 и высвобождая четыре иона H +.
Существует различный противоречивый механизм вышеуказанного процесса в PS II, хотя протоны и электроны, извлеченные из воды, используются для уменьшения NADP + и в производстве АТФ. Фотосистема II также известна как водно-пластохиноноксидоредуктаза и считается первым белковым комплексом в световой реакции.
Ключевые различия между Фотосистемой I и Фотосистемой II
Данные точки будут демонстрировать различия между фотосистемой I и фотосистемой II:
- Фотосистема I или PS I и Фотосистема II или PS II являются белково-опосредованным комплексом, и основная цель заключается в выработке энергии (АТФ и NADPH2), которая используется в цикле Кальвина, PSI использует световую энергию для преобразования NADP + в NADPH2. Он включает P700, хлорофилл и другие пигменты, в то время как PS II представляет собой комплекс, который поглощает световую энергию, включая P680, хлорофилл и вспомогательные пигменты, и переносит электроны из воды в пластохинон и, таким образом, работает в диссоциации молекул воды и производит протоны (H +) и O2.
- Фотосистема I расположена на наружной поверхности тилакоидной мембраны и связана со специальным реакционным центром, известным как P700, тогда как PS II расположен на внутренней поверхности тилакоидной мембраны, а реакционный центр известен как P680.
- Пигменты в фотосистеме 1 поглощают свет с большей длиной волны, который составляет 700 нм (P700), с другой стороны, пигменты в фотосистеме 2 поглощают свет с более короткими длинами волн, который составляет 680 нм (P680).
- Фотофосфорилирование в PS I участвует как в циклическом, так и в нециклическом фотофосфорилировании, а PS II участвует в циклическом фотофосфорилировании.
- В PS I фотолиз не происходит, хотя происходит фотосистема II.
- Фотосистема I или PS I содержит хлорофилл A-670, хлорофилл A-680, хлорофилл A-695, хлорофилл A-700, хлорофилл B и каротиноиды в соотношении 20-30: 1, тогда как в Photosystem II или PS 2 содержится хлорофилл А-660, хлорофилл А-670, хлорофилл А-680, хлорофилл А-695, хлорофилл А-700, хлорофилл В, ксантофиллы и фикобилины в соотношении 3-7: 1.
- Основная функция фотосистемы I в синтезе NADPH, где она получает электроны от PS II, а фотосистема II - в гидролизе воды и синтезе АТФ.
- Композиция ядра в PSI состоит из двух субъединиц, которые представляют собой psaA и psaB, а PS II состоит из двух субъединиц, состоящих из D1 и D2.
Вывод
Таким образом, мы можем сказать, что в растениях фотосинтез охватывает два процесса; светозависимые реакции и реакция ассимиляции углерода, которая также вводит в заблуждение также как темные реакции. В световых реакциях фотосинтетические пигменты и хлорофилл поглощают свет и преобразуются в АТФ и НАДФН (энергию).
Основой жизнедеятельности растений является фотосинтез. В процессе фотосинтеза в тканях растения из неорганических веществ образуются органические. Как и любая химическая реакция синтеза, фотосинтез проходит с поглощением энергии.
Источником энергии световой фазы фотосинтеза является свет. В процессе эволюции растения научились для своей жизнедеятельности использовать данную энергию. О световой фазе фотосинтеза мы постарались изложить кратко и понятно.
Что такое световая фаза фотосинтеза
Фотосинтез – сложный процесс, состоящий из 2 фаз: световой и темновой. Реакции протекающие в световой фазе фотосинтеза могут проходить только при освещении. Для темновой фазы свет не важен, она может проходить в любое время. Ниже представлена схема световой и темновой фазы фотосинтеза.
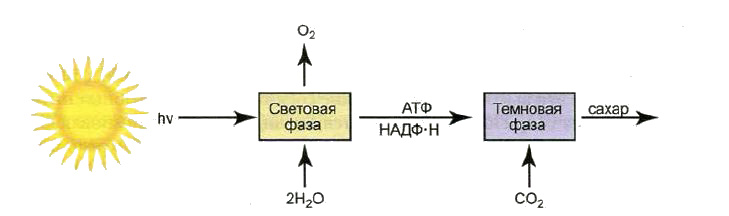
В течение световой стадии, растение захватывает фотоны света с помощью специальных светособирающих комплексов. Энергия фотонов необходима для прохождения процесса распада воды на кислород и водород.
Этот процесс, который происходит в световую фазу фотосинтеза называется фотолизом воды. Далее происходит образование конечных продуктов световой фазы, которые необходимы для прохождения реакций темновой фазы. Световая энергия накапливается в виде АТФ – аденозинтрифосфата.
Это вещество является носителем энергии и может ее высвобождать, превращаясь в АДФ – аденозиндифосфат.
Водород после фотолиза воды соединяется с ферментом НАДФ (никотинамидадениндинуклеотидфосфат), образуя НАДФН, который является источником водорода для дальнейших химических реакций.
Кислород при фотолизе воды выделяется в атмосферу. Таким образом, результатом световой фазы фотосинтеза является: распад воды под воздействием световой энергии с образованием конечных продуктов АТФ и НАДФН, использующихся для синтеза органики и свободного кислорода.
Где происходит световая фаза
Все процессы происходящие в световой стадии и темновой фазе фотосинтеза протекают в специальных клеточных структурах, называемых хлоропластами. Хлоропласт – зеленая пластида, внутри которой содержится хлорофилл. В растительной клетке содержится большое количество хлоропластов, необходимых для прохождения химических реакций фотосинтеза.
Различные фазы процесса проходят в разных частях хлоропласта. Эта пластида имеет сложную структуру, в ее состав входит большое количество тилакоидов. Тилакоиды – особые структуры внутри хлоропласта, отвечающие за преобразование световой энергии. Тилакоиды, расположенные рядом, образуют стопки – граны.
Световая фаза фотосинтеза происходит в гранах тилакоидов, на их мембранах и во внутритилакоидном пространстве. В этом особенность световой фазы фотосинтеза и этим она отличается от темновой, во время которой химические реакции протекают в строме хлоропласта – плотном веществе между тилакоидами.

Фотохимическая суть процесса
Основным процессом световой фазы фотосинтеза является фотолиз воды, представленный следующим уравнением:
Расщепление воды под действием света происходит с помощью молекул хлорофилла, которые сосредоточены вблизи мембран тилакоидов.
Молекулы хлорофилла имеют свойство возбуждаться, и терять электроны при попадании на них кванта света. Эти электроны оседают на внешней стороне мембран, заряжая их отрицательно.
Сами молекулы хлорофилла, потерявшие электрон, восстанавливаются, отбирая электроны у воды, находящейся внутри тилакоида. При фотолизе вода расщепляется на следующие компоненты:
- Протоны водорода;
- Электроны водорода;
- Кислород.
Кислород является побочным продуктом фотосинтеза и не участвует в дальнейших процессах. Он выводится наружу из ткани растения и поступает в атмосферу. Протоны накапливаются в специальном протонном резервуаре, находящемся внутри тилакоида.
Протоны заряжают внутреннюю часть тилакоидной мембраны положительно. Таким образом, мембраны тилакоидов имеют отрицательный заряд с внешней стороны, а положительный – с внутренней. Эти заряды постепенно увеличиваются по мере накопления протонов и электронов.
Разность потенциалов между внешней и внутренней поверхностью мембраны должна достигать не менее 200mВ, чтобы начался процесс образования конечных продуктов световой стадии. Только в этом случае протоны начнут проходить сквозь каналы АТФ-синтазы, находящиеся в тилакоидных мембранах.
АТФ-синтаз – комплекс белковых молекул, обеспечивающий восстановление содержащегося в растительной клетке АДФ (аденозиндифосфат) до АТФ (аденозинтрифосфат).
Для этого используется энергия протона, проходящего тилакоидную мембрану. Прошедший мембрану протон и электрон, находящийся на внешней ее стороне вступают в реакцию с находящейся в строме хлоропласта молекулой НАДФ с образованием НАДФН.
2Н + + 2е — + НАДФ → НАДФН
Светособирающие комплексы
Только небольшая часть молекул хлорофилла поглощает энергию, отдавая электроны. Они находятся в реакционных центрах и называются молекулами-ловушками. Остальная же часть этого пигмента собрана в светособирающие комплексы, задачей которых является не поглощение, а передача энергии.
Для чего нужны светособирающие комплексы? Если бы каждая молекула хлорофилла улавливала свет, то такая работа была бы крайне неэффективной. Процесс возбуждения и потери электрона проходил бы очень редко, а структура передачи электронов была бы слишком сложной из-за очень большого количества молекул.
На самом деле существует очень мало молекул, поглощающих энергию и отдающих электроны. На каждую из них приходится до 300 молекул, собранных в светособирающие комплексы по антенному типу. Они расположены на нескольких уровнях.
На первом уровне сосредоточено наибольшее количество молекул, улавливающих свет. Они передают энергию на более низкий уровень, где молекул уже гораздо меньше.
Конечно, происходит передача не квантов света, а только энергии, полученной при поглощении света. Таким образом, хлорофилл может не только поглощать световую энергию, но и передавать ее.

На самом нижнем уровне светособирающего комплекса находится 1 молекула-ловушка. Энергия поступает к ней со всего антенного комплекса. Передача энергии происходит с определенными потерями ее количества.
Но молекула-ловушка получает энергию в десятки или даже в сотни раз чаще, чем молекулы, расположенные на самом высоком уровне. Молекулы-ловушки входят в состав фотосистем, которые участвуют в транспорте электронов во время световой стадии фотосинтеза.
Основные компоненты цепи переноса электронов
В течение световой стадии происходит перенос электронов от тилакоидных фотосистем с помощью промежуточных веществ-переносчиков до образования конечных продуктов фазы. Электрон-транспортная цепь имеет сложную структуру и множество компонентов.
Основными компонентами цепи переноса электронов являются:
- Фотосистема 1;
- Фотосистема 2;
- Комплекс цитохромов b6-f;
- Вещества-переносчики;
- НАДФ-редуктаза.
Фотосистема 1 способна поглощать свет с длиной волны 700 нм, фотосистема 2 – 680 нм. Фотосистемы работают параллельно. При поглощении света фотосистемы отдают электроны на вещества-переносчики или акцепторы. В электрон-транспортной цепи задействовано множество акцепторов, которые захватывают электроны и отдают их другому компоненту цепи.
Когда фотосистема 2 теряет электрон под воздействием света, он сначала попадает на акцептор феофитин. Далее в его транспорте принимает участие целый ряд акцепторов, последним из которых является пластоцианин. Далее электрон попадает в фотосистему 1, восполняя электрон, отданный этой фотосистемой под воздействием кванта света.
Фотосистема 1 отдает электрон на акцептор ферредоксин. Отсюда он поступает в последний компонент цепи НАДФ-редуктазу. Здесь образуется в световую фазу фотосинтеза вещество НАДФН. Недостачу электронов фотосистема 1 восполняет за счет электронов, приходящих от фотосистемы 2.

Особое значение в электрон-транспортной цепи имеет комплекс цитохромов b6-f. Электроны, проходя через этот комплекс, многократно взаимодействуют с акцептором пластохиноном. При этом комплекс цитохромов увеличивает количество, не только электронов, но также и протонов, что повышает эффективность световой стадии.
Продукты световой стадии
При прохождении этапа световой фазы фотосинтеза образуются следующие продукты, необходимые для синтеза органики в дальнейших темновых реакциях: АТФ и НАДФН. АТФ – источник биохимической энергии. Эта молекула синтезируется из АДФ при поглощении энергии движущегося протона.
Формулу синтеза АТФ во время световой фазы фотосинтеза можно представить в следующем формуле:
АДФ + ортофосфорная кислота + энергия → АТФ + Н2О
Синтезированный АТФ может участвовать во всех химических реакциях, для прохождения которых необходима энергия. При взаимодействии с водой происходит обратная реакция с выделением энергии.
АТФ вновь расщепляется на АДФ и ортофосфорную кислоту:
АТФ + H2O → АДФ + ортофосфорная кислота + энергия
Для образования органических веществ при фотосинтезе такая энергетическая составляющая крайне необходима, так как синтез органики требует поглощения большого количества энергии. НАДФН – восстановленный фермент, который является источником водорода. Он используется в химических процессах темновой фазы, где отдает водород и превращается в фермент НАДФ.
Читайте также: