
Фотодыхание кратко и понятно
Обновлено: 04.07.2024
Фотодыхание у С3-растений обычно усиливается при низком содержании С02 и высоких концентрациях 02. В этих условиях РДФ-карбоксилаза в хлоропластах может функционировать как оксигеназа, катализируя окислительное расщепление рибулозо-1,5-дифосфата на 3-ФГК и 2-фосфогликолевую кислоту, которая затем дефосфорилируется в гликолевую кислоту (рис. 7.1). Молекулы С02 и 02 конкурируют между собой в каталитическом центре РДФ-карбоксилазы: при относительно высоких концентрациях С02 и низких 02 преобладает карбоксилирование, тогда как высокие концентрации 02 и низкое содержание С02 благоприятствуют окислению, а следовательно, и образованию фосфогликолевой кислоты. В том же направлении действует и повышение температуры.
Фотодыхание осуществляется в результате взаимодействия трех органелл — хлоропластов, пероксисом и митохондрий (рис. 7.1). Гликолат из хлоропласта поступает в пероксисому и там окисляется гликолатоксидазой до глиоксилата. Возникающая перекись водорода устраняется каталазой пероксисомы. Глиоксилат аминируется, превращаясь в глицин, причем в качестве донора аминогруппы функционирует глутамат.
Глицин транспортируется в митохондрию. Здесь из двух молекул глицина образуется серин и освобождается С02. Теперь цикл замыкается: серин может снова поступать в пероксисому и там передать свою аминогруппу на пируват. При этом из пирувата возникает аланин, из серина — гидроксипируват, который сразу после этого восстанавливается в глицерат. Затем глицерат может снова попасть в хлоропласты и благодаря фосфорилированию включиться в цикл Кальвина.
Последовательность реакций фотодыхания не обязательно образует цикл. Гликолатный путь С3-растений может завершиться в митохондриях. Конечными продуктами в этом случае является серин и С02. Освобождение С02 объясняет почему нетто-фотосинтез (чистая продуктивность) при интенсивном фотодыхании снижается (рис. 7.1).
Однако для чего же тогда нужно фотодыхание? Необходимо вспомнить, что гликолатный путь приводит к синтезу глицина и серина, в пероксисомах происходит восстановление NADP+; имеются косвенные данные о том, что в процессе образования серина может генерироваться и АТР. Показано, что С3-растения, помещенные в атмосферу с низким парциальным давлением 02 и высокой концентрацией С02, ведут себя подобно С4-растениям, т. е. имеют низкий уровень фотодыхания.
Из всего сказанного можно сделать вывод, что термин фотодыхание имеет лишь формальный смысл: 02 потребляется, С02 выделяется, однако в функциональном плане к дыханию этот процесс прямого отношения не имеет.

8 Эндогенные механизмы регуляции процесса фотосинтеза
Регуляция фотосинтеза осуществляется на уровне фотохимически активных мембран, хлоропластов, клеток, тканей, органов и целого организма. Все системы регуляции принимают участие в этом процессе.
Появление NADPH, ATP, 02, Mg2+ и изменение рН оказывают прямое и опосредованное влияние на реакции фотосинтетического усвоения С02 в строме. Экспериментально показано, что фиксация С02 изолированными хлоропластами практически не обнаруживается при рН ниже 7,2, но достигает максимальных значений при рН 8,0. Причина этого заключается в изменении активности ряда ферментов, имеющих оптимум рН в слабощелочной среде: рибулозофосфаткиназы (рН 7,9), дегидрогеназы фосфоглицеринового альдегида (рН 7,8), рибулозо-1,5-дифосфаткарбоксилазы (рН от 7,5 до 8,0), фруктозодифосфатазы (рН 7,5 — 8,5). Повышение рН стромы при освещении хлоропласта приводит к значительному связыванию С02 в виде Н2С03 и накоплению НСОз, из которого С02 освобождается с помощью карбоангидразы перед карбоксилированием рибулозо-1,5-дифосфата.
АТР участвует в фосфорилировании рибулозо-5-фосфата и ФГК, а также в реакциях, связанных с синтезом сахарозы и крахмала. NADPH необходим для восстановления ФГК до ФГА и для образования малата из щавелевоуксусной кислоты. Mg2+ нужен для функциональной активности РДФ-карбоксилазы, он также принимает участие в синтезе хлорофилла и белков.
Процесс фиксации С02 регулируется прежде всего светом.
Он активирует ряд ферментов цикла Кальвина: РДФ-карбоксилазу, дегидрогеназу 1,3-дифосфоглицериновой кислоты, фруктозо-1,6-дифосфатфосфатазу, седогептулозо-1,7-дифосфатфосфатазу и рибулозо-5-фосфаткиназу. Наиболее многообразна регуляция функционирования ключевого фермента цикла Кальвина — РДФ-карбоксилазы.

Помимо света, фермент активируется фруктозо-6-фосфатом, а ингибируется 6-фосфоглюконатом и фруктозо-1,6-дифосфатом. 6-Фосфоглюко-нат подавляет также активность завершающего цикл фермента — рибулозо-5-фосфаткиназы. Наконец, продукт цикла фиксации С02 — 3-фосфоглицериновая кислота положительно влияет на синтез крахмала. Повышение концентрации 02 в строме может привести к снижению интенсивности фиксации С02 за счет усиления фотодыхания.
Функциональные изменения в клетках мезофилла. Хлоропласты большинства растений способны перемещаться в клетке в зависимости от интенсивности и направления освещения. Сильный свет вызывает отрицательный фототаксис хлоропластов: они уходят от света, концентрируясь на боковых стенках клеток палисадной паренхимы; слабый свет вызывает положительный фототаксис. Предполагают, что освещенный хлоропласт меняет свое положение в клетке с помощью сократительных белков, связанных с оболочкой хлоропласта и взаимодействующих с сократительными белками цитоплазмы.
Мощное воздействие на клетку, определяющее ее метаболизм, оказывают образующиеся в хлоропластах ATP, NADPH и ассимиляты. При освещении отношение ATP/ADP в хлоропластах резко возрастает и вследствие быстрого обмена энергетическими эквивалентами почти тотчас же может воспроизводиться в цитоплазме, а через нее и в митохондриях. Так как оболочка хлоропласта лишь в слабой степени проницаема для АТР, перенос в цитоплазму высокоэнергетической фосфатной связи АТР осуществляется с помощью челночного механизма в результате реакции восстановления-окисления между ФГК и ФДА, так как скорость выхода триозофосфатов через мембраны оболочки хлоропласта приближается к скорости свободной диффузии. Одновременно это приводит к транспорту восстановительных эквивалентов (NADH).

Переносчиком высокого восстановительного потенциала NADPH может служить система ЩУК-малат, каждый компонент которой легко проходит через мембраны, совершая циклические перемещения между хлоропластом и цитоплазмой. В пользу этой гипотезы говорит то, что в хлоропластах находятся NAD + - и NADP + -зависимые малатдегидрогеназы.
Уже в первые секунды фотосинтеза происходит выход ассимилятов в цитоплазму. После кратковременного фотосинтеза с 14С02 в цитоплазме прежде всего появляются меченые фосфотриозы — ФГК и ФДА, а затем [14С] фруктозо-1,6-дифосфат. На изолированных хлоропластах показано, что оболочка хлоропласта практически непроницаема для сахарозы. По-видимому, основным местом синтеза сахарозы в клетке является не хлоропласт, а цитоплазма. На это указывает также присутствие основного сахарозосинтезирующего фермента сахарозофосфатсинтетазы и UDP-глюкозопирофосфорилазы преимущественно в цитоплазме.
Выход предшественников углеводов из хлоропластов в цитоплазму совершается в виде ФГК, ФГА и ФДА. Дальнейшие превращения, ведущие к синтезу гексозофосфатов и сахарозы, происходят уже в цитоплазме. Аналогичным превращениям могут подвергаться триозофосфаты и в хлоропластах, вследствие чего при достаточно интенсивном фотосинтезе в хлоропластах накапливается крахмал, служащий резервом углеводов, предназначенных для экспорта в цитоплазму во вторую очередь. Отложения крахмала часто бывают очень значительными (до 90% от объема хлоропласта).
Выходящие из хлоропластов ассимиляты могут использоваться в самой фотосинтезирующей клетке двумя путями: 1) в окислительно-восстановительных процессах, что обычно заканчивается освобождением С02, 2) для увеличения массы клетки в процессе ее роста и отложения запасных и вторичных веществ. Большая часть ассимилятов транспортируется из фотосинтезирующих клеток листа в другие органы и ткани.
Взаимодействие тканей листа при фотосинтезе. Под действием света изменения происходят не только в клетках мезофилла. Свет выступает одним из важнейших факторов регуляции работы устьиц. При включении света у большинства растений устьица открываются более широко, а при выключении — закрываются. Исключение из правила — растения семейства толстянковых, у которых ночью устьица открыты, а днем закрыты. На движения устьиц влияет и концентрация С02: ширина устьичных щелей увеличивается при снижении содержания С02 в межклетниках. Открывание устьиц под действием света связано с работой фотосинтетического аппарата в замыкающих клетках. У этиолированных растений свет не влияет на движения устьиц. В замыкающих клетках на свету включается механизм Н +-насоса в плазмалемме, усиливается поглощение К+ и синтез малата, в результате чего возрастает внутриклеточное осмотическое давление и устьица открываются.
Увеличение степени открывания устьиц приводит к усилению транспирации и, следовательно, к более интенсивному поступлению воды и веществ по ксилеме из корней в листья, что необходимо для нормальной функциональной активности клеток мезофилла.
Фотосинтетическая деятельность клеток мезофилла обогащает ткани листа сахарами и другими продуктами фотосинтеза. В результате возрастает функциональная активность проводящих пучков. Теоретически существуют два способа транспорта ассимилятов к проводящим пучкам: по симпласту (через плазмодесмы и цитоплазму последовательного ряда клеток) и по апопласту (по клеточным стенкам). Однако у многих видов растений между клетками мезофилла и флоэмы плазмодесмы развиты крайне слабо или совсем отсутствуют. В апопласте листовой пластинки может находиться около 1/5 сахаров, содержащихся в листе, и значительная доля свободных аминокислот. Клетки листовой паренхимы сравнительно легко выделяют ассимиляты в наружную среду и относительно слабо их поглощают. Клетки флоэмных окончаний, напротив, способны усиленно поглощать из внешних растворов сахара и аминокислоты против концентрационного градиента с помощью энергозависимых переносчиков. Есть основание считать, что, выходя из паренхимных клеток листа в клеточные стенки, сахароза расщепляется находящейся там инвертазой на гексозы (фруктозу и глюкозу), которые в проводящих пучках вновь образуют сахарозу.
Флоэмные окончания выполняют в листе роль коллекторов, аккумулирующих ассимиляты против градиента концентрации. Ведущая роль в загрузке проводящих элементов ассимилятами принадлежит сопровождающим и паренхимным клеткам флоэмы, а у некоторых видов растений и более специализированным передаточным клеткам. Есть данные о том, что клетки флоэмных окончаний поглощают сахара с участием ионов Н +, которые выкачиваются из клеток благодаря работе энергозависимого Н +-насоса.
При перегрузке тканей листа ассимилятами интенсивность фотосинтеза снижается. Переполнение крахмалом пластид вызывает обратимые, а позднее и необратимые изменения структуры хлоропластов, что значительно ослабляет фотосинтез. Отрицательное действие избытка крахмала на фотосинтез в пластидах может быть объяснено: 1) механическим воздействием на тилакоиды, 2) ухудшением светового режима в хлоропласте, 3) сорбцией ферментов на зернах крахмала, 4) сорбцией ионов, особенно Mg2 + , необходимых для поддержания высокой активности ферментов, 5) ухудшением диффузии С02.
Регуляция процессов фотосинтеза в целом растении.
Механизм этих взаимоотношений основан на явлении метаболической репрессии фотосинтеза. Чем мощнее центры, аттрагирующие ассимиляты, тем эффективнее фотосинтезирующий лист освобождается от ассимилятов, что служит необходимым условием интенсивного фотосинтеза.
Во всех этих процессах важнейшую роль играют фитогормоны и эндогенные ингибиторы роста и метаболизма (некоторые полифенолы). Участие фитогормонов в процессах фотосинтеза можно представить в виде следующей схемы, предложенной А. Т. Мокроносовым (1983):

Согласно этой схеме, фитогормоны образуются в разных частях растений, в том числе в хлоропластах, и действуют на процессы фотосинтеза как дистанционно, так и непосредственно на уровне хлоропластов. Дистанционное действие осуществляется благодаря регулирующему влиянию фитогормонов на процессы роста и развития (эпигенез), на отложение веществ в запас, на транспорт ассимилятов, т. е. на формирование и активность аттрагирующих центров. С другой стороны фитогормоны оказывают прямое действие на функциональную активность хлоропластов через изменение состояния мембран, активность ферментов, генерацию трансмембранного потенциала. Доказана также роль фитогормонов, в частности цитокинина, в биогенезе хлоропластов, синтезе хлорофилла и ферментов цикла Кальвина.
Ряд фитогормонов (индолилуксусная кислота, гиббереллины, абсцизовая кислота), а также некоторые физиологически активные фенолы образуются в тканях листа. Цитокинин же, от которого во многом зависит формирование листа и его структур, поступает из других частей растения, прежде всего из корня. Такая система создает взаимозависимость всех органов, обеспечивая регуляцию функциональной активности в целом растении.

Из-за наличия оксигеназной активности у фермента рибулозобисфосфаткарбоксилазы, катализирующего присоединение CO2 к рибулозо-1,5-бифосфату на начальной стадии цикла Кальвина, вместо ассимиляции углекислоты под воздействием того же фермента возможно окисление рибулозофосфата с распадом его на 3-фосфоглицериновую кислоту, которая может поступить в цикл, и на фосфат гликолевой кислоты. Он дефосфорилируется и гликолевая кислота транспортируется в пероксисомы, где окисляется до глиоксилевой кислоты и аминируется с получением глицина. В митохондриях из двух молекул глицина образуется серин и углекислый газ. Серин может использоваться для синтеза белка, либо превращается в 3-фосфоглицериновую кислоту и возвращается в цикл Кальвина.
Фотодыхание снижает эффективность фотосинтеза, приводит к потерям ассимилированного углерода, однако имеет некоторое синтетическое значение. На ранних этапах развития жизни, когда в атмосфере было мало кислорода, рубиско заняло ключевую позицию в фотосинтезе, и ее оксигеназная функция не доставляла проблем. По мере увеличения содержания кислорода потери на фотодыхание нарастали, и у ряда растений возникли механизмы активной доставки к месту работы рубиско углекислого газа (см. CAM-фотосинтез), увеличивающее долю её карбоксилазной активности до 100%.
См. также
Ссылки
Wikimedia Foundation . 2010 .
Полезное
Смотреть что такое "Фотодыхание" в других словарях:
фотодыхание — фотодыхание … Орфографический словарь-справочник
ФОТОДЫХАНИЕ — световое дыхание, совокупность процессов, происходящих в растит, клетках под действием света, в результате к рых поглощается кислород и выделяется СО2. Механизм Ф. и участвующие в нём ферменты изучены недостаточно. Полагают, что при Ф.… … Биологический энциклопедический словарь
Фотодыхание — световое дыхание, совокупность процессов, происходящих в растительных клетках под действием света, в результате которых поглощается кислород и выделяется CO2. Механизм Ф. и участвующие в нём ферменты изучены недостаточно. Полагают, что… … Большая советская энциклопедия
фотодыхание — фотодых ание, я … Русский орфографический словарь
ФОТОДЫХАНИЕ — активируемое светом образование гликолевой кислоты в хлоропластах и ее последующее окисление до CO2 в пероксисомах … Словарь ботанических терминов
Физиология растений — биологическая наука, изучающая общие закономерности жизнедеятельности растительных организмов. Ф. р. изучает процессы поглощения растительными организмами минеральных веществ и воды, процессы роста и развития, цветения и плодоношения,… … Большая советская энциклопедия
Фотосинтез — (от Фото. и Синтез образование высшими растениями, водорослями, фотосинтезирующими бактериями сложных органических веществ, необходимых для жизнедеятельности как самих растений, так и всех др. организмов, из простых соединений (например … Большая советская энциклопедия
МИКРОТЕЛЬЦА — пероксисомы, окружённые одинарной мембраной плазматич. пузырьки (диам. 0,3 1,5 мкм) в клетках позвоночных, высших растений, простейших. М. производные эндоплазматич. сети. Содержат каталазу и нек рые окислит, ферменты. Участвуют в расщеплении… … Биологический энциклопедический словарь
ФОТОСИНТЕЗ — (от фото. и греч. synthesis соединение), образование клетками высших растений, водорослей и нек рыми бактериями органич. веществ при участии энергии света. Происходит с помощью пигментов (хлорофиллов и нек рых других), присутствующих в… … Биологический энциклопедический словарь
фотосинтез — [тэ], а; м. Спец. Процесс образования углеводов из углекислоты и воды под действием света, поглощаемого хлорофиллом, в клетках зелёных растений, водорослей и некоторых микроорганизмов. ◁ Фотосинтетический, ая, ое. Ф ая деятельность. Ф ая… … Энциклопедический словарь

Фотодыхание (гликолатный путь, С2-фотосинтез) — стимулируемое светом выделение углекислого газа и поглощение кислорода у растений преимущественно с С3-типом фотосинтеза. Также под фотодыханием понимают биохимический путь, связанный с регенерацией одной молекулы 3-фосфоглицериновой кислоты (С3) из двух молекул гликолевой кислоты (С2) и лежащий в основе вышеописанного газообмена. Наличие биохимического механизма фотодыхания обусловлено значительной оксигеназной активностью РуБисКО, ключевого фермента цикла Кальвина.
Поглощение кислорода в ходе фотодыхания обусловлено оксигеназной активностью РуБисКО в хлоропластах и работой оксидазы гликолевой кислоты в пероксисомах. Кроме того, окисление образовавшегося в митохондриях НАДН также сопряжено с поглощением кислорода. Выделение углекислого газа (С1) при фотодыхании происходит в митохондриях и связано с конденсацией двух молекул глицина (С2) с образованием одной молекулы серина (С3) (последовательная работа двух ферментов: глициндекарбоксилазы и серингидроксиметилтрансферазы). Также в реакции конденсации глицина в митохондриях выделяется аммиак, который реутилизируется в результате работы глутаминсинтетазы и глутаминоксоглутаратаминотрансферазы (ГС/ГОГАТ-путь). При фотодыхании расходуется АТФ (не происходит запасания энергии) синтезированный в ходе фотофосфорилирования. Также окисление гликолевой кислоты в пероксисомах в ходе фотодыхания служит основным источником токсичного пероксида водорода в фотосинтезирующей растительной клетке.
Эффект Варбурга — ингибирование выделения O2 и ассимиляции CO2 по мере увеличения концентрации O2 в среде — широко распространен у растений с циклом Кальвина.
Он обусловлен конкуренцией CO2 и O2 за субстрат ввиду проявления РБФК свойств, характерных для карбоксилаз и оксигеназ. Фотосинтез даже при нормальных концентрациях CO2 и O2 в среде приводит к образованию не только ФГК, но и гликолата.
Фотодыхание — окисление гликолата в глиоксилат на свету через редуктивный ПФП с усиленным потреблением кислорода и выделением CO2 — накладывает отпечаток на темновое дыхание. При этом излишки электронов фотосинтетической системы выводятся из цепи и используются для синтеза АТФ. Не вызывает сомнения наличие светозависимого потребления O2 у части растений как результат подавления темнового дыхания при фотосинтезе. Однако фотодыхание не всегда оказывается связано с фотосинтезом. Так, фотодыхание наблюдается у бесхлорофиллоносных мутаций хлореллы на сильном свету, которые способны синтезировать только каротиноиды. У таких мутантов фотодыхание составляет 50% от уровня темнового дыхания. В зеленых клетках при отключении фотосинтеза дыхательным ядом также происходит потребление O2 и выделение CO2.
При фотодыхании в 3—5 раз быстрее выделяется CO2, чем при темновом дыхании (М. Зелитч, 1976). Получается так, что, с одной стороны, в процессе фотосинтеза фиксируется CO2, с другой — она тут же теряется на 50% при фотодыхании. Природа здесь оказалась как бы не на уровне защиты интересов своего произведения. И среди растений С3-типа фотосинтеза обнаружены мутанты с блокированным фотодыханием.
В связи с подобными наблюдениями возникает вопрос, насколько С3-растения могут обходиться без фотодыхания. При осуществлении фотодыхания преобладает оксигеназная функция РБФК. При естественных концентрациях CO2 и O2 в атмосфере соотношение между фотосинтезом и фотодыханием у С3-растений составляет 3:1. Поскольку соотношение указанных газов в среде (в том числе и в клетке) часто меняется под влиянием температуры и времени суток (из-за колебания концентрации прежде всего O2), постольку необходимым оказывается переключение функции РБФК на оксигеназную функцию, и наоборот. Все зависит от того, какой газ находится в избытке.
Образование гликолата служит началом фотодыхания. Дальнейшее его превращение происходит в хлоропластах, пероксисомах и митохондриях. У большинства растений гликолат в хлоропластах образуется при падении концентрации CO2 и увеличении концентрации O2. Фотодыханию отводится некоторая положительная роль в обеспечении клеток промежуточными веществами (глутамат, γ-глутаровая кислота, глицин, серин, глицерат и т. п.), важном для дальнейших синтезов. Все это свидетельствует, что фотодыхание нельзя рассматривать как бесполезный или вредный процесс.
Фотодыхание способствует поддержанию постоянства CO2 внутри листа. У растений С4-пути существует другой механизм поддержания концентрации CO2 (фиксация его ЩУК). Фотодыхание тесно связано с общим метаболизмом в зеленой клетке, оно часто усиливается при уменьшении потребности растений в продуктах фотосинтеза и, в принципе, направлено на поддержание активности ферментативных систем и функции хлоропластов и митохондрий (А. Т. Мокроносов, 1983). Его роль иногда связывают с метаболизмом азотистых соединений, переносом их из одной органеллы в другую, а также с возможностью ограничения Накопления фосфогликолата.
Явление фотодыхания открыто в 1955 г. И. Деккером. Оно в большей мере выражено у растений С3-пути фотосинтеза и сопровождается потерей углерода в цикле Кальвина из-за снижения способности РБФК фиксировать CO2 и повышения ее активности окислять субстрат (РБФ) в присутствии O2 (А. Лайск, 1977). В случае окисления субстрата РБФК выступает в роли РБФ-оксигеназы. На примере рода Panicum, в пределах которого встречаются виды с С3-, С4— и промежуточными путями фотосинтеза, выявлены различия в содержании органелл. У P. riculare (С3-путь) клетки обкладки содержат в среднем 3 хлоропласта, 1,1 митохондрий и 0,3 пероксисом; у Р. milioides, Р. decipiens и Р. schenckii (промежуточные луга) органелл соответственно 10,6,17,7 и 3,2; у Р. priontis (С4-путь) в клетках обкладки органелл не только больше, но они и крупнее. У указанных видов фотодыхание снижается в направлении увеличения числа органелл в клетках обкладки (R. Н. Brow et al., 1983).
Начальные стадии фотодыхания (образование гликолевой кислоты) характерны для всех организмов с циклом Кальвина. Виды же различаются механизмами метаболизма гликолевой кислоты. Так, фотосинтезирующие бактерии и цианеи окисляют гликолат и синтезируют глицин. Цианеи содержат фермент гликолатдегидрогеназу и способны поглощать гликолат (в темноте и на свету), но утилизируют его слабо. У эукариотических водорослей в процессе фотосинтеза гликолевая кислота выделяется в окружающую среду, хотя у них несколько лучше выражен механизм утилизации гликолата (при участии фермента гликолатдегидрогеназы). Подобное выделение представляет собой способ защиты цикла Кребса от ингибирования и на начальных этапах развития жизни было необходимо для усиления симбиотических отношений бактерий и цианей с гетеротрофами (А. М. Степанова, А. А. Шумилова, 1980).
У высших же растений с С3-путем фиксации CO2 фотодыхание осуществляется наиболее сложно — путем полного превращения гликолевой кислоты, с образованием ряда важных промежуточных продуктов (аминокислот, порфиринов и др.). Дело в том, что с переходом растений к наземным условиям существования возможности выделения гликолата в среду ограничиваются ввиду невымываемости его из организма. Поэтому защита цикла Кребса от ингибирующего действия глиоксилата могла быть достигнута только путем полного превращения его в гликолат.
Несмотря на эти новшества, у высших растений потери углерода при фотодыхании составляют 50% от его ассимилированного количества. У них фотосинтез падает главным образом из-за конкуренции O2 и CO2 за молекулу РБФК и ее субстрат. Интенсивность фотодыхания у растений С3-типа возрастает при повышении концентрации O2 и температуры среды. Это ведет к ингибированию фотосинтеза в результате снижения сродства РБФК к CO2, особенно в аридных условиях. Поэтому отбор способствовал формированию растений С4-типа, у которых хорошо развита система концентрации CO2 (ФЕП, органические кислоты и др.) и выражена активность РБФК (при одновременном подавлении оксигеназной функции указанного фермента). Благодаря подобным и другим изменениям у растений С4-типа потери углерода при фотосинтезе невелики и гликолат накапливается, медленно.
Представляет интерес оценка роли фотодыхания в онтогенезе. Как известно, в экстремальных условиях водоснабжения устьица закрываются и резко возрастает фотодыхание. Это имеет значение для зашиты хлоропластов от разрушения (путем снабжения CO2) и сохранения нормального соотношения НАДФ/НАДФ ∙ Н2, в результате чего поддерживается процесс фотосинтеза и. растения предохраняются от истощения и теплового шока. В то же время образуемый H2O2 при взаимодействии O2 с ЭТЦ хлоропластов может способствовать отравлению тканей. Во избежание этого часть его также используется в хлоропластах для неэнзиматического окисления глиоксилевой кислоты. Возникающий на промежуточных стадиях фотодыхания АТФ метаболизируется в самих митохондриях, где его концентрация резко падает из-за ограничения оттока триозофосфата из, хлоропластов и поступления CO2 в ткани. Фотодыхание возрастает по мере старения растений и листьев (А. Т. Мокроносов, 1981).
Ведущее место в фотодыхании принадлежит пероксисомам. Ни один другой органоид клетки не выполняет столь разнообразные функции, как микротельца (А. У. Игамбердиев, 1988, 1990), в том числе и при генерации новых путей биохимических превращений. Фотодыхание, хотя и рассматривается как ответвление фотосинтетического пути, играет важную роль в обеспечении нормального функционирования Цикла Кальвина, снабжая его CO2 в обход реакций карбоксилирования. Оно также устраняет ингибирование ЦТК метаболизацией гликолата. По этим и другим причинам необходимо было выработать механизм утилизации гликолата, от которого ранее (водоросли) избавлялись выделением в среду. С появлением пероксисом у части водорослей (хлорелла) окисление гликолата в глиоксилат с образованием H2O2 имело значение для рассеивания свободной энергии, поглощенной клеткой. Выделение гликолата в среду его вымыванием имело значение для развития симбиотических отношений автотрофов и гетеротрофов, но оно не могло получить дальнейшего развития в наземных условиях, где исключалась возможность этого вымывания. Отсюда и необходимость отбора растений в направлении усиления утилизации гликолата в энергетических целях. Косвенно эти механизмы способствовали усилению транспорта воды, минеральных и органических веществ у сосудистых растений благодаря формированию вакуолярной системы. Интересно отметить, что в случае повторного перехода цветковых к обитанию в воде они начинают выделять гликолат в среду.
По-видимому, роль фотодыхания наиболее велика в условиях постоянного обезвоживания клеток растений как механизма для предотвращения падения концентрации CO2 и истощения субстрата дыхания (путем стабилизации фотосинтеза). Быть может, этим и объясняется подавленность гликолатно-глиоксилатного пути у С4-растений и при наличии в клетках обкладки соответствующих ферментов — эти растения имеют механизм для поддержания фотосинтеза даже при Закрытых устьицах. При длительном ограничении доступа CO2 из внешней среды и у С4-растений начинает функционировать гликолатный путь.
Энергетическая эволюция растений связана с увеличением экологического разнообразия видов и форм по эффективному использованию и сочетанию механизмов фотосинтеза и дыхания в различных условиях. Это достигалось преимущественным выживанием в критических ситуациях индивидуумов — носителей благоприятных мутаций по синтезу пигментов и набору ферментов. Поэтому энергетическая система растений развивалась постепенно, дополняя ранее возникшие механизмы новыми реакциями обмена.
Значительны различия видов растений в экологии дыхания, в чем проявляются характерные черты их приспособления. Показателем приспособленности растений в онто- и филогенезе служит и фотодыхание. Механизмы дыхания как энергетическая основа жизнедеятельности оказались базисом для возникновения последующего многообразия растений, в том числе эволюции их химического состава и ритмики.
По типу питания живые организмы делятся на автотрофы, гетеротрофы и миксотрофы. Автотрофы (греч. αὐτός — сам + τροφ - пища) - организмы, которые самостоятельно способны синтезировать органические вещества из неорганических. Гетеротрофы (греч. ἕτερος - иной + τροφή - пища) - организмы, использующие для питания готовые органические вещества.
Наконец, миксотрофы (греч. μῖξις - смешение + τροφή - пища) - организмы, которые могут использовать как гетеротрофный, так и автотрофный способ питания. К примеру, эвглена зеленая на свету начинает фотосинтезировать, а в темноте питается гетеротрофно.

Фотосинтез
Фотосинтез (греч. φῶς - свет и σύνθεσις - синтез) - сложный химический процесс преобразования энергии квантов света в энергию химических связей. В результате фотосинтеза происходит синтез органических веществ из неорганических.

Этот процесс уникален и происходит только в растительных клетках, а также у некоторых бактерий. Фотосинтез осуществляется при участии хлорофилла (греч. χλωρός - зелёный и φύλλον - лист) - зеленого пигмента, окрашивающего органы растений в зеленый цвет. Существуют и другие вспомогательные пигменты, которые вместе с хлорофиллом выполняют светособирающую или светозащитную функции.
Ниже вы увидите сравнение строения хлорофилла и гемоглобина. Обратите внимание, что в центре молекулы хлорофилла находится ион Mg.
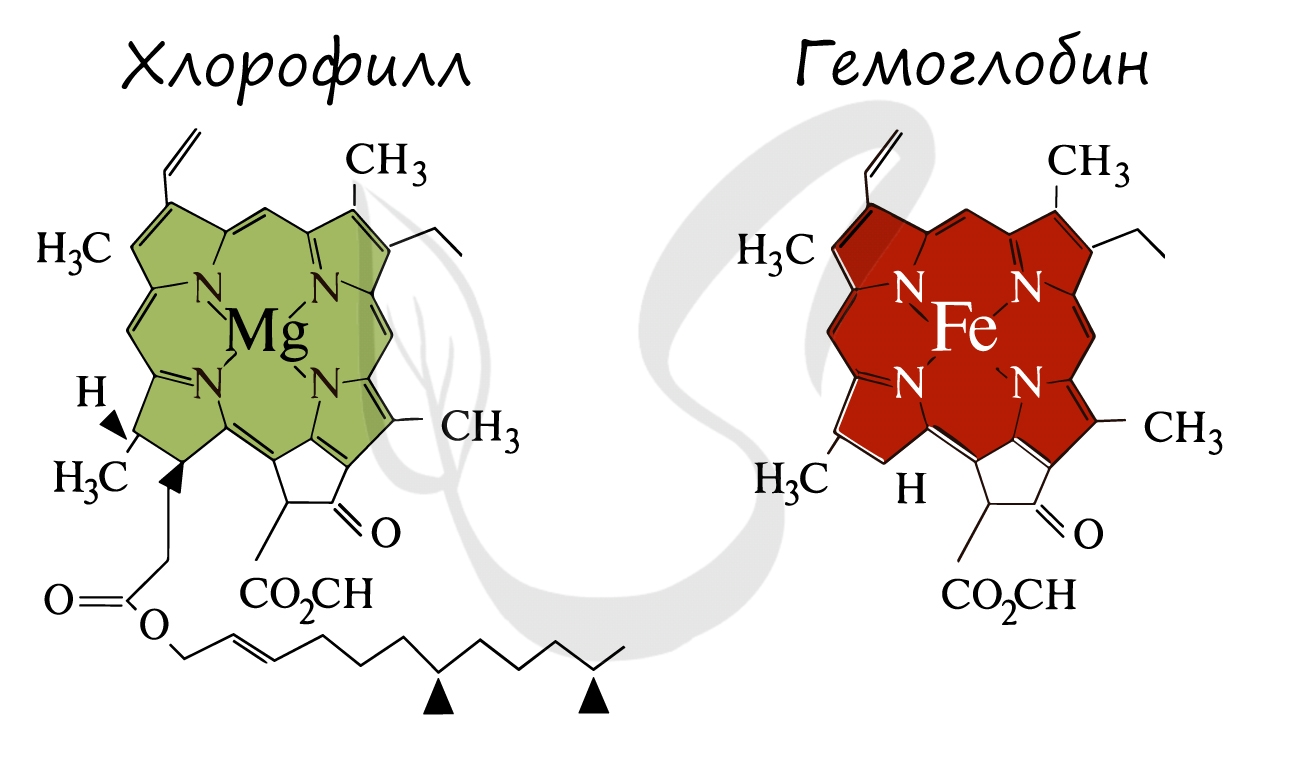
В высшей степени гениально значение процесса фотосинтеза подчеркнул русский ученый К.А. Тимирязев: "Все органические вещества, как бы они ни были разнообразны, где бы они ни встречались, в растении ли, в животном или человеке, прошли через лист, произошли от веществ, выработанных листом. Вне листа или, вернее, вне хлорофиллового зерна в природе не существует лаборатории, где бы выделялось органическое вещество. Во всех других органах и организмах оно превращается, преобразуется, только здесь оно образуется вновь из вещества неорганического"
Более подробно мы обсудим значение фотосинтеза в завершение этой статьи. Фотосинтез состоит из двух фаз: светозависимой (световой) и светонезависимой (темновой). Я рекомендую использовать названия светозависимая и светонезависимая, так как они способствуют более глубокому (и правильному!) пониманию фотосинтеза.
Светозависимая фаза (световая)
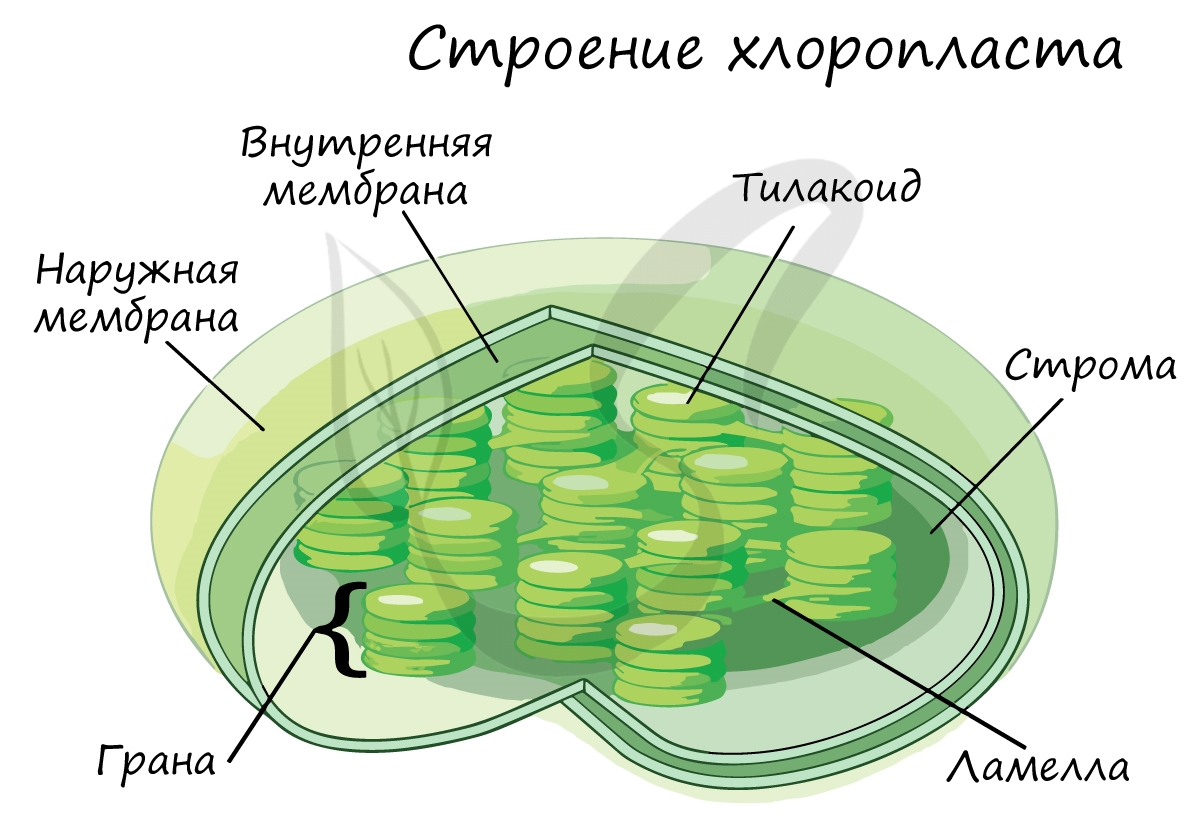
Эта фаза происходит только на свету на мембранах тилакоидов в хлоропластах. В ней принимают участие различные ферменты, белки-переносчики, молекулы АТФ-синтетазы и зеленый пигмент хлорофилл.
Хлорофилл выполняет две функции: поглощения и передачи энергии. При воздействии кванта света хлорофилл теряет электрон, переходя в возбужденное состояние. С помощью переносчиков электроны скапливаются с наружной поверхности мембраны тилакоидов, тем временем внутри тилакоида происходит фотолиз воды (разложение под действием света):
Гидроксид-ионы отдают лишний электрон, превращаясь в реакционно способные радикалы OH, которые собираются вместе и образуют молекулу воды и свободный кислород (это побочный продукт, который в дальнейшем удаляется в ходе газообмена).
Образовавшиеся при фотолизе воды протоны (H + ) скапливаются с внутренней стороны мембраны тилакоидов, а электроны - с внешней. В результате по обе стороны мембраны накапливаются противоположные заряды.
При достижении критической разницы, часть протонов проталкивается на внешнюю сторону мембраны через канал АТФ-синтетазы. В результате этого выделяется энергия, которая может быть использована для фосфорилирования молекул АДФ:
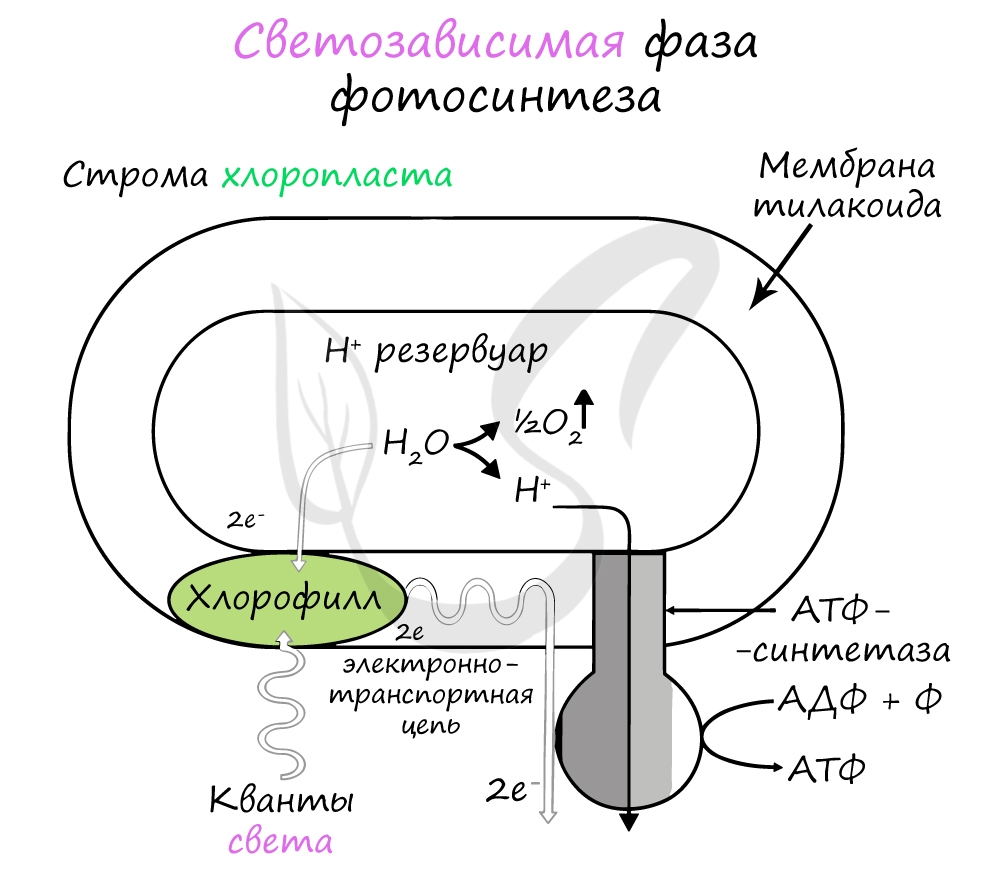
Протоны, попав на поверхность мембраны тилакоидов, соединяются с электронами и образуют атомарный водород, который используется для восстановления молекулы-переносчика НАДФ (никотинамиддинуклеотидфосфат). Благодаря этому окисленная форма - НАФД + превращается в восстановленную - НАДФ∗H2.
- Свободный кислород O2 - в результате фотолиза воды
- АТФ - универсальный источник энергии
- НАДФ∗H2 - форма запасания атомов водорода
Кислород удаляется из клетки как побочный продукт фотосинтеза, он совершенно не нужен растению. АТФ и НАДФ∗H2 в дальнейшем оказываются более полезны: они транспортируются в строму хлоропласта и принимают участие в светонезависимой фазе фотосинтеза.
Светонезависимая (темновая) фаза
Светонезависимая фаза происходит в строме (матриксе) хлоропласта постоянно: и днем, и ночью - вне зависимости от освещения.
При участии АТФ и НАДФ∗H2 происходит восстановление CO2 до глюкозы C6H12O6. В светонезависимой фазе происходит цикл Кальвина, в ходе которого и образуется глюкоза. Для образования одной молекулы глюкозы требуется 6 молекул CO2, 12 НАДФ∗H2 и 18 АТФ.
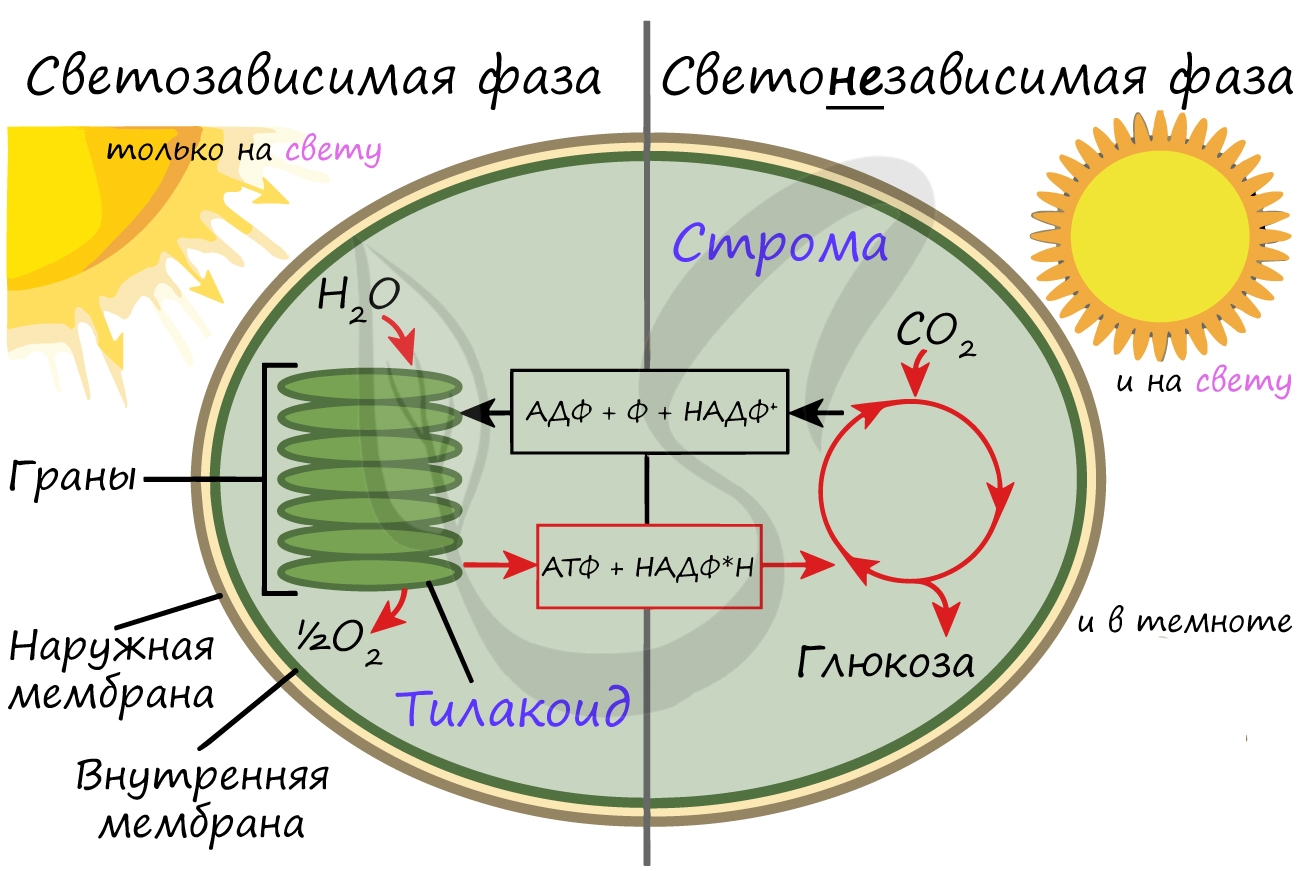
Таким образом, в результате темновой (светонезависимой) фазы фотосинтеза образуется глюкоза, которая в дальнейшем может быть преобразована в крахмал, служащий для запасания питательных веществ у растений.
Значение фотосинтеза
Значение фотосинтеза невозможно переоценить. Уверенно утверждаю: именно благодаря этому процессу жизнь на Земле приобрела такие чудесные и изумительные формы, какие мы видим вокруг себя: удивительные растения, прекрасные цветы и самые разнообразные животные.
В разделе эволюции мы уже обсуждали, что изначально в составе атмосферы Земли не было кислорода: миллиарды лет назад его начали вырабатывать первые фотосинтезирующие бактерии - сине-зеленые водоросли (цианобактерии). Постепенно кислород накапливался, и со временем на Земле стало возможно аэробное (кислородное) дыхание. Возник озоновый слой, защищающий все живое на нашей планете от губительного ультрафиолета.
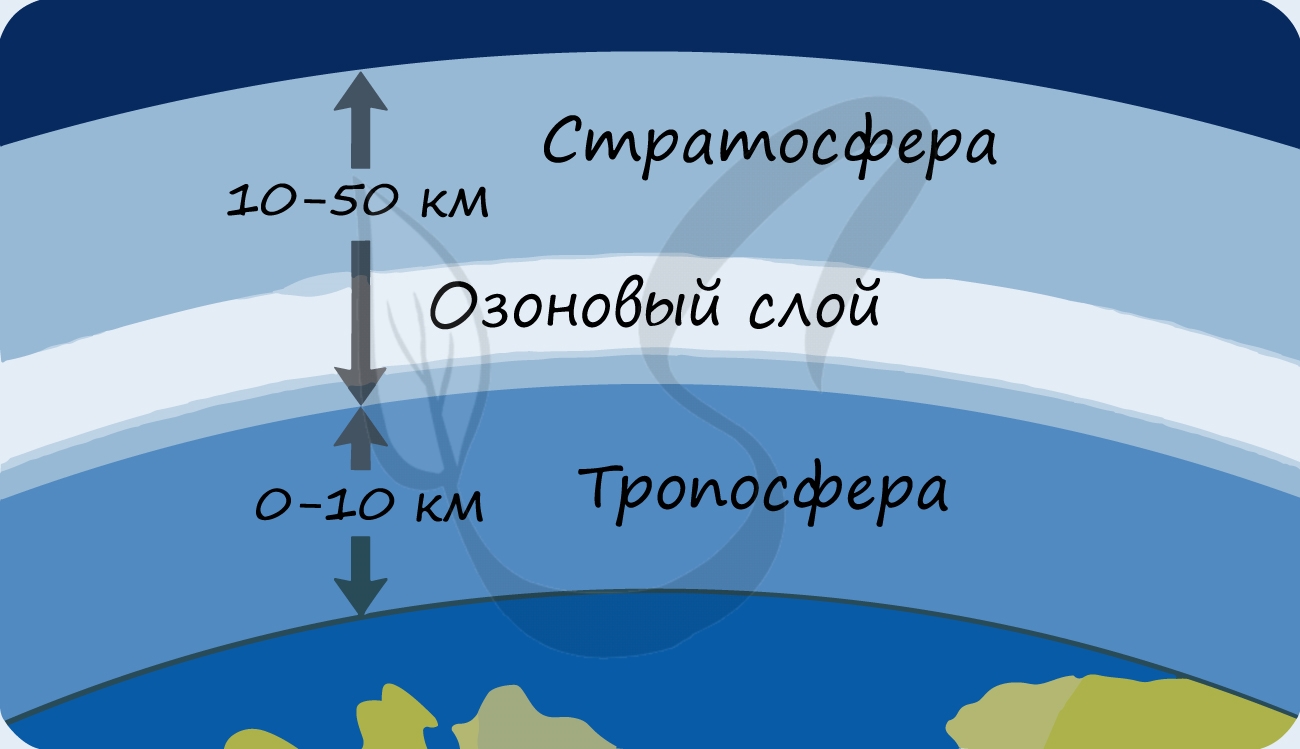
- Синтезируют органические вещества, являющиеся пищей для всего живого на планете
- Преобразуют энергию света в энергию химических связей, создают органическую массу
- Растения поддерживают определенный процент содержания O2 в атмосфере, очищают ее от избытка CO2
- Способствуют образованию защитного озонового экрана, поглощающего губительное для жизни ультрафиолетовое излучение

Хемосинтез (греч. chemeia – химия + synthesis - синтез)
Хемосинтез - автотрофный тип питания, который характерен для некоторых микроорганизмов, способных создавать органические вещества из неорганических. Это осуществляется за счет энергии, получаемой при окислении других неорганических соединений (железо- , азото-, серосодержащих веществ).
Хемосинтез был открыт русским микробиологом С.Н. Виноградским в 1888 году. Большинство хемосинтезирующих бактерий относится к аэробам, для жизни им необходим кислород.
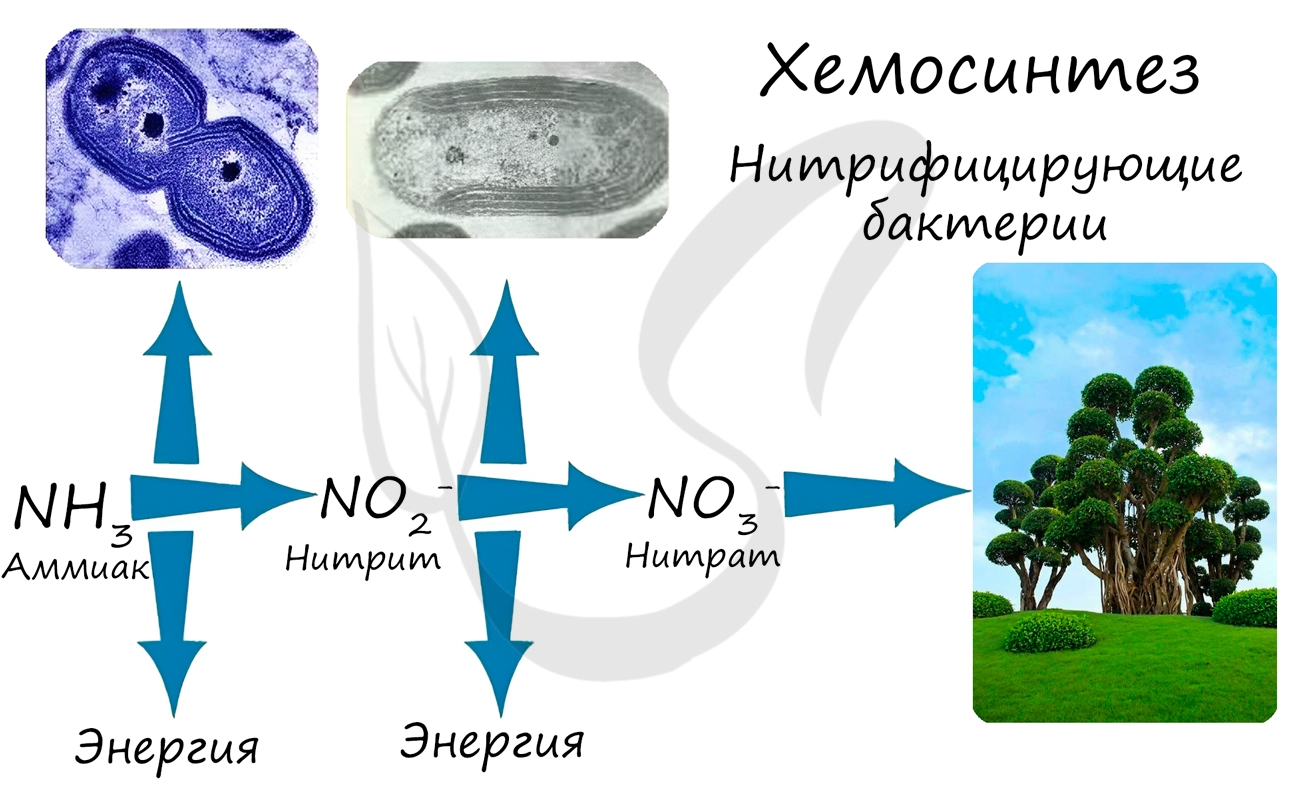
При окислении неорганических веществ выделяется энергия, которую организмы запасают в виде энергии химических связей. Так нитрифицирующие бактерии последовательно окисляют аммиак до нитрита, а затем - нитрата. Нитраты могут быть усвоены растениями и служат удобрением.
- Серобактерии - окисляют H2S --> S 0 --> (S +4 O3) 2- --> (S +6 O4) 2-
- Железобактерии - окисляют Fe +2 -->Fe +3
- Водородные бактерии - окисляют H2 --> H +1 2O
- Карбоксидобактерии - окисляют CO до CO2
Значение хемосинтеза
Хемосинтезирующие бактерии являются неотъемлемым звеном круговорота в природе таких элементов как: азот, сера, железо.
Нитрифицирующие бактерии обеспечивают переработку (нейтрализацию) ядовитого вещества - аммиака. Они также обогащают почву нитратами, которые очень важны для нормального роста и развития растений.
Усвоение нитратов происходит за счет клубеньковых бактерий на корнях бобовых растений, однако важно помнить, что клубеньковые (азотфиксирующие) бактерии, в отличие от нитрифицирующих бактерий, питаются гетеротрофно.
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Читайте также: