
Факторы влияющие на поглощение воды растением кратко
Обновлено: 07.07.2024
Как и всякий физиологический процесс, поглощение воды корнями зависит от тех условий, в которых находится корневая система. Очень важное значение для этого процесса имеет обеспечение корней О2. Анаэробизис - недостаток О2, подавляет поглощение воды вследствие снижения дыхания корней, образования ими АТФ и за счет угнетения неосмотического компонента корневого давления. Поэтому при выращивании растений всегда следует заботиться, чтобы среда корнеобитания была снабжена О2. Для этого почву необходимо поддерживать в рыхлом состоянии.
На поглощение воды корнями влияет и t° почвы. В узких физиологических пределах поглощение изменяется пропорционально колебаниям t°. Однако не все растения одинаково чувствительны к понижению t°. Холодоустойчивые (северные злаки, горох) могут достаточно хорошо поглощать воду и при низкой t° почвы, они не испытывают нарушений водного режима при похолоданиях. Теплолюбивые растения (кукуруза, просо) при низкой t° почвы замедляют поглощение воды вследствие снижения метаболизма корня. Поэтому в теплые дни, но при холодной почве у них могут быть нарушения водного обмена, и они будут страдать от недостатка воды, даже при высоком содержании ее в почве.
14 Транспирация, ее регуляция и значение в жизни растений.
Самая пышная и быстрорастущая растительность существует в условиях постоянно флажного тропического леса, где транспирация низкая. Однако в большинстве случаев она идет интенсивно, и растение не в силах ее ограничить и уменьшить. Причиной этого являются сложные отношения водного режима и фотосинтеза. Дело в том, что вода испаряется — выходит из листа, а субстрат фотосинтеза — двуокись углерода — входит в лист через одни и те же устьичные отверстия. Таким образом, защищаясь от излишнего испарения закрыванием устьиц, растение обрекает себя на голод; открывая устьица для нормального питания — фотосинтеза, оно обезвоживается. Значение: Во-первых, транспирация создает в раст. непрерывный водный ток и тем самым облегчает передвижение мин. веществ в органах. Во-вторых, транспирация регулирует t° тела раст., так как она непостоянна и зависит от состояния среды. Испарение воды требует значительной затраты тепловой энергии, поэтому испаряющее тело всегда охлаждается. Охлаждение тканей растения при сильной транспирации может достигать 5 — 7 °С относительно t° среды, что очень существенно для жизни растения в условиях жаркого климата.
15 Завис. Транспирации от вн. И внут. Факторов. Дневной ход трансп.
Как любой физиологический, процесс, транспирация подвержена действию внешних и внутренних факторов, которые могут значительно ее изменять. Солнечная радиация. Один из важных факторов, который активирует процесс транспирации. В определенных пределах транспирация пропорциональна солнечной радиации. Световые и тепловые лучи приводят к раскрытию устьиц и, следовательно, к ускорению испарения воды листом. Влажность воздуха также влияет на этот процесс. Транспирация, как и всякое испарение, обратно пропорциональна влажности среды и прямо пропорциональна дефициту насыщения. Т.о., при повышенной влажности воздуха интенсивность транспирации будет низкой, так как диффузия паров воды через устьица и их удаление от листа будут затруднены.
Движение воздуха — ветер. Он усиливает транспирацию, т.к. относит от листа пары воды, создавая этим не насыщенное водными парами пространство. На транспирацию влияют и некоторые внутренние факторы. Необходимо отметить зависимость транспир. от ярусности листьев: она увеличивается от нижних листьев к верхним, испаряющих самое большое количество воды. Трансп. зависит от соотношения форм воды (свободной и связанной) в растении. При увеличении содержания свободной воды она повышается, в противном случае падает.
Дневной ход транспирации
При правильном ходе метеорологических факторов и в случае, если они не очень напряжены (ясная, умеренно теплая погода с небольшим ветром, без дождя, но и без засухи), транспирация начинается утром с восходом солнца, постепенно усиливается до полудня, затем также постепенно понижается и прекращается вечером, когда солнце заходит. В том случае, когда ход метеорологических факторов также правилен, но они несколько напряжены (жарко), максимум транспирации смещается на более ранние предполуденные часы, в остальном сохраняется тот же характер. При очень сильном напряжении метеорологических факторов (очень жарко, сухо) возможно понижение транспирации в полуденные часы и вторичное повышение в предвечерние. Это указывает на неблагоприятные для существования растений условия. При неправильном ходе метеорологических факторов, когда погода в течение дня непостоянна — ход транспирации будет также неправильным, так как он отражает изменчивость метеорологических факторов.
Поглощение воды корнями растений зависит от влажности грунта. Если он отличается хорошей увлажненностью, то скорость поглощения зависит от двух факторов. Первый – интенсивность транспирации, поскольку именно от нее зависит водный потенциал в проводящих тканях корня. Второй – поглощающая способность поверхности корневой системы.
Когда грунт начинает пересыхать, то доступное количество влаги уменьшается. Причиной является понижение водного потенциала и гидравлической проводимости. Влияют на поглощение воды такие факторы как аэрация грунта, его температура, концентрация почвенного раствора и некоторые другие.
Скорость поглощения корнями воды в первую очередь зависит от такого показателя, как крутизна градиента водного потенциала от почвы к корням. Если в почве содержится меньше влаги, то она становится менее доступной, поскольку уменьшается ее потенциал, зато возрастает сопротивление движения к корням. Эта же характеристика верна и по отношению к прочим субстратам, на которых выращиваются растения. При уменьшении количества влаги на поверхности всасывающих корневых волосков повышается концентрация питательного раствора. Это препятствует также и поглощению питательных веществ, находящихся в растворе.
Понятие легкодоступная вода представляет собой количество влаги, которое находится в промежутке между полевой влагоемкостью почвы и влажностью устойчивого завядания. Полевая влагоемкость представляет собой содержание влаги в почве через несколько дней после ее увлажнения, когда движение воды вниз замедлилось. Влажность устойчивого завядания характеризуется таким содержанием влаги, при котором растения остаются завядшими, пока не будет выполнено увлажнение почвы. Стоит отметить, что в мелком грунте легкодоступной воды находится большее количество, чем в почвах с крупными элементами.
Корневая система часто страдает из-за недостатка кислорода. Она прекращает рост, снижает физиологическую активность. Чаще всего такую реакцию можно наблюдать на затопленных почвах, но нередко невыраженное кислородное голодание можно увидеть и на тяжелых глинистых почвах, проникновение кислорода в которые затруднено. При этом корни не растут вглубь, снижается активность минерального питания. Существует даже мнение, что прерии не заселяются древесными формами именно из-за плохой аэрации почвы.
Если грунт оказывается затоплен водой, то это резко уменьшает поглощение влаги . Объясняется это повышением сопротивления току воды. Но растения приспосабливаются к разным условиям и существуют огромные различия между разными видами. На затопленном грунте могут расти кипарис болотный, нисса водяная, если грунт просто сильно насыщен водой, то на нем хорошо растут ивы, ряд растений выдерживает регулярное затопление. Но существуют и противоположные по характеристикам виды, которые при затоплении быстро отмирают. К таким относятся тюльпанное дерево, а также дерен.
При недостаточной аэрации корневой системы снижается поступление воды, минеральных солей и уменьшается синтетическая активность . Некоторые виды растений в корнях синтезируют регуляторы роста: гиббереллины и цитокинины, а также аминокислоты и амиды Достоверно установлено, что подобные вещества синтезируются в корнях яблони. При переувлажнении или в случае холодной почвы побеги растут слабыми. Они страдают от недостатка воды, минеральных веществ и регуляторов роста.
Ряд авторов уверены, что температура почвы является существенным экологическим фактором, а холодная почва негативно влияет на растения. Так в почве с низкой температурой уменьшается доступность воды. Ограничения распространения лесов на горных склонах связывают, в том числе, и с данной причиной.
Как и в других случаях, данный экологический фактор по-разному влияет на разные виды растений. Козловский (1943) отметил, что при снижении температуры с 15 градусов до пяти сосна ладанная сильнее уменьшала поглощение воды, чем сосна веймутовая. Если сравнивать растения, обитающие в теплых и холодных климатических зонах, то можно отметить, что теплолюбивые растения при понижении температуры сильнее сокращают потребление воды, чем холодостойкие. Так цитрусовые хуже переносят охлаждения почвы, чем деревья из северных широт. Но наверняка и внутри этой группы имеются различия между видами.
Охлаждение почвы уменьшает поглощение воды двумя путями . Во-первых, снижается проницаемость поверхностей корней для воды. Во-вторых, при охлаждении повышается вязкость воды, в результате чего она медленнее движется через почву и корни. Два таких фактора при понижении температуры от 25 до 5 градусов удваивают уровень сопротивления току воды. Данных о влиянии высоких температур на водопоглощение относительно немного.
Протяженность корневых систем и эффективность их работы
На успешность поглощения растением воды и растворенных в ней минеральных солей удобрений влияет проницаемость корней, их протяженность и площадь контакта. Корневые системы многих деревьев имеют значительную протяженность. Они распространяются далеко за пределы пространства, очерченного кроной. Глубина проникновения ограничивается только наличием аэрации и особенностями структуры. Если рассматривать сомкнутые древостои, то можно отметить, что поверхностный слой почвы полностью заполнен корнями. Глубина такого слоя варьируется от тридцати сантиметров и до нескольких метров. Виганс (1936) отмечает, что восемнадцатилетние яблони, произраставшие в штате Небраска, на лессовидных почвах с хорошей аэрацией поглощали влагу с глубины в десять метров. Противоположные характеристики были у груш, которые росли в штате Орегон на наносно-глинистых почвах с плохой аэрацией. Примерно 90% их корневой системы располагалось в поверхностном слое почвы, толщина которого не превышала один метр. Коль (1937) при исследовании дубовых и сосновых лесов в Северной Каролине обнаружил, что примерно 90% корней находится в слое глубиной всего в 12, 5см. Количество корней в глубоких слоях почвы относительно невелико, но несмотря на это почва нередко пересыхает до стадии устойчивого завядания. Интересные данные о связи развития корней с особенностями поглощения воды и минеральных солей приводит Руссель (1973).
Существует два фактора, от которых зависит эффективность работы корней. Это размер контактной поверхности и проницаемость. Вполне очевидно, что наличие у корневой системы множества мелких корней и разветвлений увеличивает площадь контакта с грунтом и эффективность. Корневая система с небольшим количеством маловетвящихся крупных корней менее эффективна.
Микоризы, скорее всего, увеличивают способность к поглощению минеральных веществ , поскольку грибные гифы широко распространяются в почве. В результате появляется дополнительная поглощающая поверхность. Благодаря микоризе старые и уже одревесневшие корни продолжают поглощать минеральные вещества. Опередить роль гифов грибов в поглощении воды деревьями сложнее. Некоторые исследователи указывают, что присутствие микоризы делает сеянцы древесных растений более засухоустойчивыми. Кроме того, в процессе исследований установлено, что присутствие микоризы уменьшает сопротивление при движении воды. Как было отмечено выше, опробковевшие корни с низкой проницаемостью имели высокий уровень поглощения минеральных веществ и воды.
Воду из почвы поглощают корни. Однако в этом процессе участвует далеко не вся корневая система. Наибольшая часть корней покрыта пробкой, не пропускающей воду, и поэтому лишь проводит уже поглощенную воду в ствол, укрепляет дерево в почве. Всасывают воду только корешки толщиной не более полмиллиметра.
К примеру, одна взрослая береза поглощает и испаряет в день 75, а липа 200 литров воды. Один гектар столетнего дубового леса испаряет за весенне-летний период как минимум 1200, а букового леса — 2070 тонн воды.
Вместе с водой дерево поглощает из почвы минеральные соли, содержащие азот, калий, фосфор и другие питательные элементы.
МЕХАНИЗМЫ ПОГЛОЩЕНИЯ ВОДЫ
В соответствии с терминологией Реннера (1915) существуют два механизма поглощения воды: активное поглощение, обычно наблюдаемое у слабо транспирирующих растений, и пассивное, которое преобладает у интенсивно транспирирующих растений и обеспечивает бóльшую часть поглощения воды древесными растениями.
АКТИВНОЕ ПОГЛОЩЕНИЕ. Корни растений, произрастающих на теплой, хорошо аэрируемой, влажной почве, действуют как осмометры, когда растения транспирируют медленно, потому что накопление солей в ксилемном соке понижает его осмотический потенциал, следовательно, и водный потенциал, который становится меньше водного потенциала почвы. Возникающая в результате этого диффузия воды, направленная внутрь, создает корневое давление, благодаря которому происходят гуттация и "плач", наблюдаемый при поранении некоторых растений (например, березы и винограда).
ПАССИВНОЕ ПОГЛОЩЕНИЕ. Когда интенсивность транспирации увеличивается и возникает натяжение в ксилемном соке, начинается массовый приток воды. При этом уносятся соли, накопившиеся в пасоке корней, и уменьшается значение осмотического передвижения воды. Корни становятся пассивной поглощающей поверхностью, через которую вода всасывается благодаря массовому потоку, создаваемому транспирирующимй побегами.
ФАКТОРЫ, ВЛИЯЮЩИЕ НА ПОГЛОЩЕНИЕ ВОДЫ
Во влажной почве скорость поглощения воды контролируется в основном двумя факторами: интенсивностью транспирации, поскольку она в значительной степени определяет величину водного потенциала в ксилеме корня, и эффективностью деятельности корневых систем как поглощающих поверхностей. При высыхании почвы доступность воды начинает ограничиваться вследствие понижения водного потенциала и гидравлической проводимости. Аэрация и температура почвы, концентрация и состав почвенного раствора также иногда ограничивают поглощение воды.
ПОЧВЕННАЯ ВЛАГА. Скорость поглощения воды зависит от крутизны градиента водного потенциала от почвы до корней. При высыхании почвы вода становится все менее доступной, так как ее потенциал уменьшается, а сопротивление ее передвижению по направлению к корням повышается. Легкодоступной водой часто считают ту, которая находится между полевой влагоемкостью и влажностью устойчивого завядания. Полевая влагоемкость - это содержание воды через несколько дней после того, как почва была увлажнена и нисходящее передвижение воды стало очень медленным. Влажность устойчивого завядания - это содержание воды, при котором растения остаются завядшими, пока почва не будет вновь увлажнена.
АЭРАЦИЯ ПОЧВЫ. Рост и физиологическая активность корней часто понижаются из-за недостатка кислорода. Наиболее резко это проявляется в затопленных почвах, однако хронический, но умеренный недостаток кислорода часто существует в тяжелых глинистых почвах, что ограничивает проникновение корней вглубь и, возможно, поглощение минеральных питательных веществ.

Затопление почвы водой обычно резко понижает ее поглощение, так как увеличивает сопротивление току воды в корни. Однако существуют большие различия между видами по их устойчивости к затоплению. Кипарис болотный и нисса водная растут неограниченно долго на затопленной почве, ивы буйно разрастаются на почве, насыщенной водой, многие виды выдерживают ежегодные периоды затопления, но некоторые виды, например дерен и тюльпанное дерево, быстро отмирают на насыщенной водой почве.
Недостаточная аэрация и холодная почва могут вызывать ослабление роста побегов, уменьшая их снабжение регуляторами роста и органическими азотистыми соединениями, а также замедляя поглощение воды и солей.
ТЕМПЕРАТУРА ПОЧВЫ. Пониженная доступность воды в холодной почве на больших высотах над уровнем моря может влиять на растительность и на расположение верхней границы распространения леса. Плохо дренированные почвы медленно нагреваются весной. Имеются данные, что холодные почвы европейских верховых болот ограничивают рост растений.
Существуют значительные различия между видами в отношении влияния низкой температуры на поглощение воды. Козловский (1943) установил, что при понижении температуры почвы от 15 до 5°С поглощение воды у сосны ладанной сокращалось сильнее, чем у сосны веймутовой. Вообще, у видов из теплого климата обнаруживается большее снижение поглощения, чем у видов из холодного климата. Цитрусовые деревья больше страдают от холодной почвы, чем древесные породы из более холодного климата, но между разновидностями и видами существуют, вероятно, различия.
Холодная почва уменьшает поглощение воды двумя путями: прямо - вследствие понижения проницаемости корней для воды, и косвенно, увеличивая вязкость воды, что замедляет ее передвижение как через почву, так и через корни.
ПРОТЯЖЕННОСТЬ И ЭФФЕКТИВНОСТЬ РАБОТЫ КОРНЕВЫХ СИСТЕМ. Успешное поглощение любым растением воды и минеральных веществ зависит от протяженности и проницаемости корней. Глубина залегания корней древесных растений может достигать 20 м и более (песчаная акация, тамарикс и др.). у столетней сосны насчитывается около 5 млн корневых окончаний, а общая длина корневой системы (без корневых волосков) достигает 50 км.
Корневая система является главным аппаратом поглощения воды,но не единственным: небольшое количество воды поглощается надземными частями растения во время дождей, выпадения туманов, росы.
Список литературы
1. Агапова Н. Д. Семейство лютиковые (Ranunculaceae) // Жизнь растений. В 6-ти т. Т. 5. Ч. 1. Цветковые растения / Под ред. А. Л. Тахтаджяна. — М.: Просвещение, 1980. — С. 210—216.
2. Еленевский А.Г., Соловьева М.П., Тихомиров В.Н. Ботаника. Систематика высших или наземных растений. – М.: Академия, 2009. - 432 с.
3. Жуковский П.М. Ботаника. – М.: Колос, 2008. – 623 с.
4. Лотова Л.И. Морфология и анатомия высших растений. - М.: КомКнига, 2007. - 510с.
Содержание
| 1. | Поглощение воды деревом. Влияние внутренних и внешних факторов на поступление воды. |
| 2. | Какие растения называют светолюбивыми, тенелюбивыми и теневыносливыми. Световой и теневой тип листа. |
| 3. | Сущность процесса дыхания и его значение для жизнедеятельности растений. |
| 4. | Движение растений. Положительные и отрицательные тропизмы. Настии. |
| 5. | Бактерии, их строение, размножение, питание. |
| 6. | Лишайники, их морфологическое, анатомическое строение и размножение. |
| 7. | Строение и цикл развития селагинеллы. Изобразите чередование поколений в жизненном цикле равноспоровых и разноспоровых плаунов. |
| 8. | Краткая характеристика семейств лютиковые и розоцветные. Назовите и опишите лесные травянистые растения из данных семейств, укажите условия их местопроизрастания и хозяйственное значение. |
| 9. | Список литературы |
Поглощение воды деревом. Влияние внутренних и внешних факторов на поступление воды.
Воду из почвы поглощают корни. Однако в этом процессе участвует далеко не вся корневая система. Наибольшая часть корней покрыта пробкой, не пропускающей воду, и поэтому лишь проводит уже поглощенную воду в ствол, укрепляет дерево в почве. Всасывают воду только корешки толщиной не более полмиллиметра.
К примеру, одна взрослая береза поглощает и испаряет в день 75, а липа 200 литров воды. Один гектар столетнего дубового леса испаряет за весенне-летний период как минимум 1200, а букового леса — 2070 тонн воды.
Вместе с водой дерево поглощает из почвы минеральные соли, содержащие азот, калий, фосфор и другие питательные элементы.
МЕХАНИЗМЫ ПОГЛОЩЕНИЯ ВОДЫ
В соответствии с терминологией Реннера (1915) существуют два механизма поглощения воды: активное поглощение, обычно наблюдаемое у слабо транспирирующих растений, и пассивное, которое преобладает у интенсивно транспирирующих растений и обеспечивает бóльшую часть поглощения воды древесными растениями.
АКТИВНОЕ ПОГЛОЩЕНИЕ. Корни растений, произрастающих на теплой, хорошо аэрируемой, влажной почве, действуют как осмометры, когда растения транспирируют медленно, потому что накопление солей в ксилемном соке понижает его осмотический потенциал, следовательно, и водный потенциал, который становится меньше водного потенциала почвы. Возникающая в результате этого диффузия воды, направленная внутрь, создает корневое давление, благодаря которому происходят гуттация и "плач", наблюдаемый при поранении некоторых растений (например, березы и винограда).
ПАССИВНОЕ ПОГЛОЩЕНИЕ. Когда интенсивность транспирации увеличивается и возникает натяжение в ксилемном соке, начинается массовый приток воды. При этом уносятся соли, накопившиеся в пасоке корней, и уменьшается значение осмотического передвижения воды. Корни становятся пассивной поглощающей поверхностью, через которую вода всасывается благодаря массовому потоку, создаваемому транспирирующимй побегами.
ФАКТОРЫ, ВЛИЯЮЩИЕ НА ПОГЛОЩЕНИЕ ВОДЫ
Во влажной почве скорость поглощения воды контролируется в основном двумя факторами: интенсивностью транспирации, поскольку она в значительной степени определяет величину водного потенциала в ксилеме корня, и эффективностью деятельности корневых систем как поглощающих поверхностей. При высыхании почвы доступность воды начинает ограничиваться вследствие понижения водного потенциала и гидравлической проводимости. Аэрация и температура почвы, концентрация и состав почвенного раствора также иногда ограничивают поглощение воды.
ПОЧВЕННАЯ ВЛАГА. Скорость поглощения воды зависит от крутизны градиента водного потенциала от почвы до корней. При высыхании почвы вода становится все менее доступной, так как ее потенциал уменьшается, а сопротивление ее передвижению по направлению к корням повышается. Легкодоступной водой часто считают ту, которая находится между полевой влагоемкостью и влажностью устойчивого завядания. Полевая влагоемкость - это содержание воды через несколько дней после того, как почва была увлажнена и нисходящее передвижение воды стало очень медленным. Влажность устойчивого завядания - это содержание воды, при котором растения остаются завядшими, пока почва не будет вновь увлажнена.
АЭРАЦИЯ ПОЧВЫ. Рост и физиологическая активность корней часто понижаются из-за недостатка кислорода. Наиболее резко это проявляется в затопленных почвах, однако хронический, но умеренный недостаток кислорода часто существует в тяжелых глинистых почвах, что ограничивает проникновение корней вглубь и, возможно, поглощение минеральных питательных веществ.
Затопление почвы водой обычно резко понижает ее поглощение, так как увеличивает сопротивление току воды в корни. Однако существуют большие различия между видами по их устойчивости к затоплению. Кипарис болотный и нисса водная растут неограниченно долго на затопленной почве, ивы буйно разрастаются на почве, насыщенной водой, многие виды выдерживают ежегодные периоды затопления, но некоторые виды, например дерен и тюльпанное дерево, быстро отмирают на насыщенной водой почве.
Недостаточная аэрация и холодная почва могут вызывать ослабление роста побегов, уменьшая их снабжение регуляторами роста и органическими азотистыми соединениями, а также замедляя поглощение воды и солей.
ТЕМПЕРАТУРА ПОЧВЫ. Пониженная доступность воды в холодной почве на больших высотах над уровнем моря может влиять на растительность и на расположение верхней границы распространения леса. Плохо дренированные почвы медленно нагреваются весной. Имеются данные, что холодные почвы европейских верховых болот ограничивают рост растений.
Существуют значительные различия между видами в отношении влияния низкой температуры на поглощение воды. Козловский (1943) установил, что при понижении температуры почвы от 15 до 5°С поглощение воды у сосны ладанной сокращалось сильнее, чем у сосны веймутовой. Вообще, у видов из теплого климата обнаруживается большее снижение поглощения, чем у видов из холодного климата. Цитрусовые деревья больше страдают от холодной почвы, чем древесные породы из более холодного климата, но между разновидностями и видами существуют, вероятно, различия.
Холодная почва уменьшает поглощение воды двумя путями: прямо - вследствие понижения проницаемости корней для воды, и косвенно, увеличивая вязкость воды, что замедляет ее передвижение как через почву, так и через корни.
ПРОТЯЖЕННОСТЬ И ЭФФЕКТИВНОСТЬ РАБОТЫ КОРНЕВЫХ СИСТЕМ. Успешное поглощение любым растением воды и минеральных веществ зависит от протяженности и проницаемости корней. Глубина залегания корней древесных растений может достигать 20 м и более (песчаная акация, тамарикс и др.). у столетней сосны насчитывается около 5 млн корневых окончаний, а общая длина корневой системы (без корневых волосков) достигает 50 км.
Корневая система является главным аппаратом поглощения воды,но не единственным: небольшое количество воды поглощается надземными частями растения во время дождей, выпадения туманов, росы.
Корневая система является главным аппаратом поглощения воды, но не единственным: небольшое количество воды поглощается надземными частями растения во время дождей, выпадения туманов, росы. В принципе любая клетка может всасывать воду, если она не насыщена ею, даже кутикула способна разбухать.
Размеры корневых систем растений огромны. Хлебные злаки способны развивать корневую систему, достигающую глубины 1,5 - 2 м и широко распространяющуюся в стороны. Глубина залегания корней древесных растений может достигать 20 м и более (песчаная акация, тамарикс и др.). у столетней сосны насчитывается около 5 млн корневых окончаний, а общая длина корневой системы (без корневых волосков) достигает 50 км.
Все многообразие древесных растений по характеру строения и развития корневых систем делят на три основные группы:
· с глубоко идущей в почву якорной корневой системой (лиственница, дуб, липа, робиния и др.);
· с поверхностной корневой системой (ель, ясень, рябина и др.);
· с промежуточным типом корневых систем, включающим в зависимости от лесорастительных условий как глубоко идущую, так и поверхностную корневую систему (сосна, береза, осина, многие клены, ольха и др.).
Далеко не вся корневая система всасывает воду. Этой способностью обладают лишь молодые корневые окончания. Вода поглощается клетками зоны корневых волосков, а также зоны растяжения. Поглощающие корни относятся к последнему порядку ветвления. Для них характерно первичное строение, отсутствие камбиального слоя. Основная их функция поглощение воды и элементов минерального питания из почвы, тогда как для ростовых - увеличение размеров корневых систем.
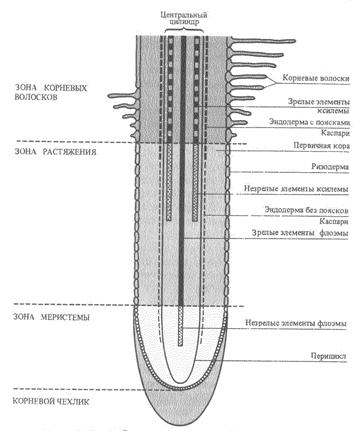
Длина всасывающей части корня травянистых растений достигает нескольких сантиметров, а древесных - всего лишь нескольких миллиметров. При этом у большинства деревьев всасывающий аппарат увеличивается за счет развития микоризы - симбиоза корня с гифами гриба. Грибной чехол микоризных корней и отходящие от него отдельные гифы существенно увеличивают поглощающую поверхность корня. Оказалось также, что микоризные растения более устойчивы к засухе, чем растения без микоризы: грибница продолжает активно всасывать воду при таком осмотическом давлении почвенного раствора, когда безмикоризные корни уже не работают.
По многолетним данным В.П. Рябчука, сезонная сокопродуктивность одного дерева березы в различных районах страны (в зависимости от диаметра и количества подсочных каналов) составляет 37 - 360 л.
В пасоке березы при весеннем сокодвижении содержится до 1,5 % сахаров, а у отдельных деревьев до 3,5 % (сахароза, глюкоза и фруктоза). В небольших количествах в березовом соке обнаруживаются аминокислоты, жирные кислоты, минеральные соли и некоторые другие вещества. Эти качества пасоки широко используют в народном хозяйстве. Она добывается в процесс е так называемой ксилемной подсочки берез, кленов и некоторых других пород.
Под ксилемной подсочкой понимается совокупность производственных процессов, связанных с добычей ксилемного сока лиственных древесных растений. Добываемая жидкость (пасока) используется для приготовления сока, сиропа, кваса, вина, спирта, а также в качестве добавок в парфюмерно-косметическом производстве, медицине, животноводстве и пчеловодстве. Более
всего заготавливалось березового сока среди стран СНГ на Украине (в среднем 35 тыс. т/год), Белоруссии и России. Грамотное ведение ксилемной подсочки березы в 5 раз экологически более эффективно, чем сбыт березовой древесины с единицы лесопокрытой площади. Еще более выгодна пневая подсочка берез. Ксилемная подсочка лиственных древесных растений существенно снижает и затраты картофеля и других ценных с/х продуктов в спиртовом производстве.

Иногда можно наблюдать выделение капелек жидкости из неповрежденных листьев. Это гуттация. Она ярко проявляется у проростков злаков, У каллы, манжетки, земляники. Из древесных растений нередко гуттируют ольха, ивы, вязы и некоторые другие. Гуттация обусловлена тем, что при высокой влажности воздуха и почвы в теплую погоду вода, поступившая в растение, не испаряется, а выдавливается в виде капелек жидкости. У срезанного побега дерева или кустарника даже при 100 % влажности воздуха гуттация наблюдаться не будет, ибо отсутствует корневое давление.
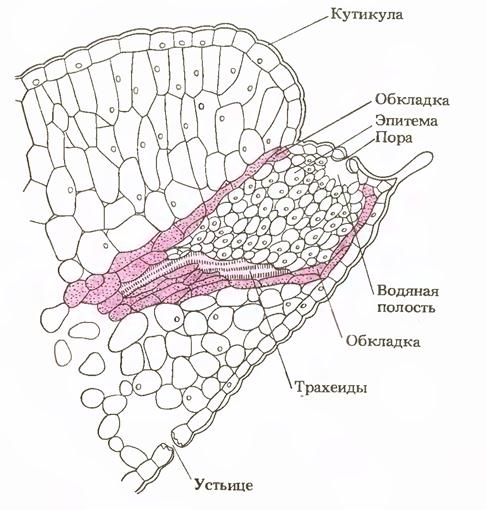
Роль процесса дыхания в проявлении корневого давления может быть не только прямой, но и косвенной. Энергия этого процесса способствует поглощению ионов солей из почвы, передвижению их по живым клеткам корня и выделению в полости сосудов центрального цилиндра корня. При интенсивном дыхании концентрация солей в сосудах корня увеличивается, а вместе с ней повышается осмотическое давление раствора в сосудах.
Существуют две основные гипотезы, объясняющие механизм действия корневого давления:
Осмотuческая теория основана на явлениях осмоса. В сосудах центрального цилиндра поглощающего корня всегда находится раствор определенной концентрации, обладающий сосущей силой, равной целиком осмотическому давлению, так как в мертвых элементах ксилемы отсутствует тургорное давление. Поэтому вода будет отсасываться от ближайшей к сосуду клетки. Теряя воду, эта клетка будет развивать большую сосущую силу, что позволит ей отнимать воду от соседней с ней клетки и т.д. Клетки коровой паренхимы и центрального цилиндра корня будут испытывать недонасыщение водой. Этот дефицит коснется и корневого волоска, способного поглощать воду из почвы. При этом распределение величины осмотического давления в ряду клеток, про водящих воду, не играет никакой роли, ибо основным фактором осмотического поглощения воды выступает сосущая сила клеток.
Метаболuческая теория, выдвинутая Д.А Сабининым, основывается на допущении, что клетки корня, через которые наблюдается односторонний ток воды, обладают полярностью. На противоположных сторонах осуществляются диаметрально разные процессы: на одной (обращенной к периферии) синтезируются осмотически активные сахара, а на противоположной - инертный в осмотическом отношении крахмал. В первом случае осмотическое давление и сосущая сила будут больше, следовательно, к этой стороне клетки будет поступать вода от соседней клетки и выдавливаться с противоположного конца в сторону сосуда. Так создается непрерывный односторонний ток воды в корневой системе, как результат развиваемого в корнях корневого давления.
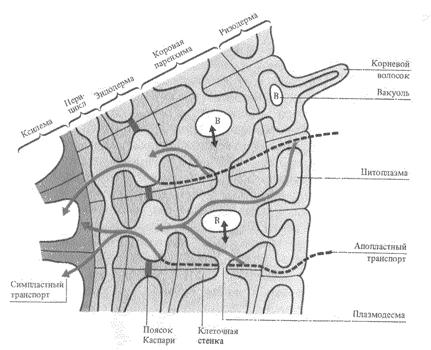
В области эндодермы корня вода проходит через ее пропускные клетки, но их сравнительно мало. Другие клетки эндодермы с суберинизированными поясками Каспари, тесно связанными с плазмалеммой, представляют сильное препятствие току воды и растворов. По существу, эндодерма - основной барьер, препятствующий одностороннему току воды из клеток первичной коры в центральный цилиндр корня. Недавно с помощью современной техники было показано, что растворы проходят только через протопласты клеток эндодермы, тогда как в других клетках - как через симпласт, так и через апопласт (по клеточным стенкам).
Величина корневого давления обычно колеблется около 1 атм, реже 2,5 - 3,0 атм. С помощью корневого давления растения в ночное время суток восполняют водный дефицит, возникающий в жаркий летний день.
Корневое давление, у древесных растений действует в течение всего года, в том числе зимой, пополняя водные запасы ствола. Как установил Л. А. Иванов, после спиливания дерева влажность древесины пня повышается под действием корневого давления. Особенно велико значение корневого давления ранней весной до распускания листьев, когда отсутствует притягивающая воду транспирация. Корневое давление можно определить с помощью манометра, прикрепленного к пеньку срезанного растения.
В обычных условиях роль пассивного поглощения воды в водообмене растений существенно выше. Оно более выгодно и в энергетическом отношении, так как не связано с затратой внутренней энергии, присущей активному поглощению, а осуществляется за счет энергии Солнца.
Корневая система является главным аппаратом поглощения воды, но не единственным: небольшое количество воды поглощается надземными частями растения во время дождей, выпадения туманов, росы. В принципе любая клетка может всасывать воду, если она не насыщена ею, даже кутикула способна разбухать.
Размеры корневых систем растений огромны. Хлебные злаки способны развивать корневую систему, достигающую глубины 1,5 - 2 м и широко распространяющуюся в стороны. Глубина залегания корней древесных растений может достигать 20 м и более (песчаная акация, тамарикс и др.). у столетней сосны насчитывается около 5 млн корневых окончаний, а общая длина корневой системы (без корневых волосков) достигает 50 км.
Все многообразие древесных растений по характеру строения и развития корневых систем делят на три основные группы:
· с глубоко идущей в почву якорной корневой системой (лиственница, дуб, липа, робиния и др.);
· с поверхностной корневой системой (ель, ясень, рябина и др.);
· с промежуточным типом корневых систем, включающим в зависимости от лесорастительных условий как глубоко идущую, так и поверхностную корневую систему (сосна, береза, осина, многие клены, ольха и др.).
Далеко не вся корневая система всасывает воду. Этой способностью обладают лишь молодые корневые окончания. Вода поглощается клетками зоны корневых волосков, а также зоны растяжения. Поглощающие корни относятся к последнему порядку ветвления. Для них характерно первичное строение, отсутствие камбиального слоя. Основная их функция поглощение воды и элементов минерального питания из почвы, тогда как для ростовых - увеличение размеров корневых систем.
Длина всасывающей части корня травянистых растений достигает нескольких сантиметров, а древесных - всего лишь нескольких миллиметров. При этом у большинства деревьев всасывающий аппарат увеличивается за счет развития микоризы - симбиоза корня с гифами гриба. Грибной чехол микоризных корней и отходящие от него отдельные гифы существенно увеличивают поглощающую поверхность корня. Оказалось также, что микоризные растения более устойчивы к засухе, чем растения без микоризы: грибница продолжает активно всасывать воду при таком осмотическом давлении почвенного раствора, когда безмикоризные корни уже не работают.
По многолетним данным В.П. Рябчука, сезонная сокопродуктивность одного дерева березы в различных районах страны (в зависимости от диаметра и количества подсочных каналов) составляет 37 - 360 л.
В пасоке березы при весеннем сокодвижении содержится до 1,5 % сахаров, а у отдельных деревьев до 3,5 % (сахароза, глюкоза и фруктоза). В небольших количествах в березовом соке обнаруживаются аминокислоты, жирные кислоты, минеральные соли и некоторые другие вещества. Эти качества пасоки широко используют в народном хозяйстве. Она добывается в процесс е так называемой ксилемной подсочки берез, кленов и некоторых других пород.
Под ксилемной подсочкой понимается совокупность производственных процессов, связанных с добычей ксилемного сока лиственных древесных растений. Добываемая жидкость (пасока) используется для приготовления сока, сиропа, кваса, вина, спирта, а также в качестве добавок в парфюмерно-косметическом производстве, медицине, животноводстве и пчеловодстве. Более
всего заготавливалось березового сока среди стран СНГ на Украине (в среднем 35 тыс. т/год), Белоруссии и России. Грамотное ведение ксилемной подсочки березы в 5 раз экологически более эффективно, чем сбыт березовой древесины с единицы лесопокрытой площади. Еще более выгодна пневая подсочка берез. Ксилемная подсочка лиственных древесных растений существенно снижает и затраты картофеля и других ценных с/х продуктов в спиртовом производстве.
Иногда можно наблюдать выделение капелек жидкости из неповрежденных листьев. Это гуттация. Она ярко проявляется у проростков злаков, У каллы, манжетки, земляники. Из древесных растений нередко гуттируют ольха, ивы, вязы и некоторые другие. Гуттация обусловлена тем, что при высокой влажности воздуха и почвы в теплую погоду вода, поступившая в растение, не испаряется, а выдавливается в виде капелек жидкости. У срезанного побега дерева или кустарника даже при 100 % влажности воздуха гуттация наблюдаться не будет, ибо отсутствует корневое давление.
Роль процесса дыхания в проявлении корневого давления может быть не только прямой, но и косвенной. Энергия этого процесса способствует поглощению ионов солей из почвы, передвижению их по живым клеткам корня и выделению в полости сосудов центрального цилиндра корня. При интенсивном дыхании концентрация солей в сосудах корня увеличивается, а вместе с ней повышается осмотическое давление раствора в сосудах.
Существуют две основные гипотезы, объясняющие механизм действия корневого давления:
Осмотuческая теория основана на явлениях осмоса. В сосудах центрального цилиндра поглощающего корня всегда находится раствор определенной концентрации, обладающий сосущей силой, равной целиком осмотическому давлению, так как в мертвых элементах ксилемы отсутствует тургорное давление. Поэтому вода будет отсасываться от ближайшей к сосуду клетки. Теряя воду, эта клетка будет развивать большую сосущую силу, что позволит ей отнимать воду от соседней с ней клетки и т.д. Клетки коровой паренхимы и центрального цилиндра корня будут испытывать недонасыщение водой. Этот дефицит коснется и корневого волоска, способного поглощать воду из почвы. При этом распределение величины осмотического давления в ряду клеток, про водящих воду, не играет никакой роли, ибо основным фактором осмотического поглощения воды выступает сосущая сила клеток.
Метаболuческая теория, выдвинутая Д.А Сабининым, основывается на допущении, что клетки корня, через которые наблюдается односторонний ток воды, обладают полярностью. На противоположных сторонах осуществляются диаметрально разные процессы: на одной (обращенной к периферии) синтезируются осмотически активные сахара, а на противоположной - инертный в осмотическом отношении крахмал. В первом случае осмотическое давление и сосущая сила будут больше, следовательно, к этой стороне клетки будет поступать вода от соседней клетки и выдавливаться с противоположного конца в сторону сосуда. Так создается непрерывный односторонний ток воды в корневой системе, как результат развиваемого в корнях корневого давления.
В области эндодермы корня вода проходит через ее пропускные клетки, но их сравнительно мало. Другие клетки эндодермы с суберинизированными поясками Каспари, тесно связанными с плазмалеммой, представляют сильное препятствие току воды и растворов. По существу, эндодерма - основной барьер, препятствующий одностороннему току воды из клеток первичной коры в центральный цилиндр корня. Недавно с помощью современной техники было показано, что растворы проходят только через протопласты клеток эндодермы, тогда как в других клетках - как через симпласт, так и через апопласт (по клеточным стенкам).
Величина корневого давления обычно колеблется около 1 атм, реже 2,5 - 3,0 атм. С помощью корневого давления растения в ночное время суток восполняют водный дефицит, возникающий в жаркий летний день.
Корневое давление, у древесных растений действует в течение всего года, в том числе зимой, пополняя водные запасы ствола. Как установил Л. А. Иванов, после спиливания дерева влажность древесины пня повышается под действием корневого давления. Особенно велико значение корневого давления ранней весной до распускания листьев, когда отсутствует притягивающая воду транспирация. Корневое давление можно определить с помощью манометра, прикрепленного к пеньку срезанного растения.
В обычных условиях роль пассивного поглощения воды в водообмене растений существенно выше. Оно более выгодно и в энергетическом отношении, так как не связано с затратой внутренней энергии, присущей активному поглощению, а осуществляется за счет энергии Солнца.
Читайте также: