
Электромеханическое сопряжение в скелетных мышцах кратко
Обновлено: 05.07.2024
Связь между возбуждением и сокращением мышечного волокна описана А.Хаксли (1959). Осуществляется при помощи системы поперечных трубочек поверхностной мембраны (Т-системы) и внутриволоконного саркоплазматического ретикулума. Деполяризация, вызываемая потенциалом действия, распространяется на Т - систему и стимулирует освобождение ионов кальция из полостей ретикулума. Взаимодействие ионов кальция с регуляторным белком тропонином С приводит к активации системы сократительных белков актина и миозина. Механизм генерации потенциала действия принципиально не отличается от этого процесса в нейроне. Скорость его распространения по мембране мышечного волокна 3 - 5 м/c.
5. Режимы и виды сокращения мышц
Режимы сокращения мышцы: изотонический (когда мышца укорачивается при неизменном внутреннем напряжении, например, при нулевой массе поднимаемого груза) и изометрический (при этом режиме мышца не укорачивается, а лишь развивает внутреннее напряжение, что бывает при нагрузке неподъёмным грузом). Ауксотонический режим - при сокращении мышцы с нагрузкой вначале в мышце возрастает напряжение без укорочения (изометрический режим), затем, когда напряжение преодолевает массу поднимаемого груза, укорочение мышцы происходит без дальнейшего роста напряжения (изотонический режим).
Различают виды сокращений: одиночное и тетаническое. Одиночное сокращение возникает при действии на мышцу одиночного нервного импульса или однократного толчка тока. В миоплазме мышцы происходит кратковременный подъём концентрации кальция, сопровождаемый кратковременной работой - тягой миозиновых мостиков, сменяющейся покоем. В изометрическом режиме одиночное напряжение начинается через 2 мс после развития потенциала действия, причём напряжению предшествует кратковременное и незначительное латентное расслабление.
Тетанус - это сложное сокращение, возникающее при стимуляции с частотой выше, чем длительность одиночного мышечного сокращения. Тетанус бывает зубчатый, если мышца совершает незначительные колебания на высоте амплитуды сокращения, и гладкий - при постоянном во времени сокращении. При относительно малой частоте раздражений возникает зубчатый тетанус, при большой частоте - гладкий тетанус. Чем быстрее сокращаются и расслабляются волокна мышцы, тем чаще должны быть раздражения, чтобы вызвать тетанус.

В естественных условиях мышечные волокна работают в режиме одиночного сокращения только тогда, когда длительность интервала между разрядами мотонейронов равна или превышает длительность одиночного сокращения иннервируемых данным мотонейроном мышечных волокон. В режиме одиночного сокращения мышца способна работать длительное время без утомления, совершая при этом минимальную работу. При увеличении частоты разрядов развивается тетаническое сокращение. При зубчатом тетанусе происходит непрерывное нарастание силы сокращения и выполняемой работы. Во время гладкого тетануса мышечное напряжение не изменяется, а поддерживается на достигнутом уровне. В таком режиме мышца человека работает при развитии максимальных изометрических усилий. Работа мышцы (А) измеряется произведением массы груза (Р) и расстояния (H), на которое этот груз перемещается.
Работа может быть динамической (преобладают изотонические режимы сокращения) или статической. Она может быть преодолевающей и уступающей.
Восстановление потенциала покоя мембраны прекращает поступление из саркоплазматического ретикулума ионов кальция и дальнейший сократительный процесс. Кальций в миоплазме активирует Са-АТФ-азу, кальциевый насос осуществляет активный перенос этого иона в саркоплазматический ретикулум. Возврат мышцы в исходное, растянутое положение определяется массой костей скелета, связанных с данными мышцами и создающими растягивающее усилие после прекращения процесса сокращения. Вторым моментом является упругость мышцы, которая преодолевается в момент сокращения. Структурной основой упругости мышцы являются:
Участки прикрепления концов миофибрилл к сухожильным элементам мышечного волокна.
Для обеспечения молекулярного механизма взаимодействия актиновых и миозиновых нитей необходимы как макроэрги, так и ионы кальция. Именно Са 2+ является важнейшим компонентом электромеханического сопряжения.
Сигналом к сокращению скелетной мышцы являются нервные импульсы, поступающие из спинного или головного мозга к нервно-мышечному соединению. Далее следует синаптическая передача с участием ацетилхолина. Потенциалы действия, образовавшиеся на сарколемме распространяются по всей плазматической мембране мышечного волокна. Деполяризация распространяется и на мембраны саркоплазматического ретикулума СПС и открывает в ней потенциалзависимые кальциевые каналы. Через открывшиеся каналы Са 2+ пассивно (в сторону более низкого электрохимического потенциала) устремляется из цистерн СПС в саркоплазму и достигает миофибрилл. Около них создается достаточная для замыкания миозиновых мостиков концентрация ионов кальция. Дальше возникает сокращение (рисунок 5).
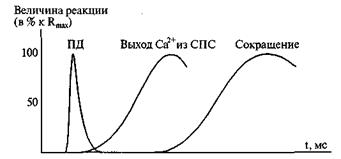
Рисунок 5. Последовательность электрических, осмотических и механических процессов в миоците поперечнополосатых мышц: по оси абсцисс - время (мс), по оси ординат - величины реакций (в % к Rmax)
Как сказано выше, для возникновения сокращения скелетной мышцы ионы кальция должны поступить к миофибриллам из саркоплазматической сети. Так называют систему пузырьков и цистерн, отделенных мембранами от остальной саркоплазмы (рисунок 6). СПС занимает примерно 10% объема мышечного волокна, а суммарная площадь ее мембран в миоците приблизительно в 100 раз больше поверхности сарколеммы (мембраны саркомеров). СПС служит кальциевым депо в мышечном волокне - содержание в ней ионов кальция огромно. Следовательно, на мембране СПС поддерживается колоссальный градиент Са 2+ , но в покое она совершенно непроницаема для этого иона.
При получении сигнала к сокращению скелетной мышцы (нервные импульсы → синаптическая передача → потенциалы действия) начинается деполяризация мембран уже саркоплзматического ретикулума. Ионы кальция из саркоплазматического ретикулума начинают выходить в саркоплазму (по механизмам пассивного транспорта по электрохимическому градиенту через кальциевые каналы, рисунок 6, В). Когда возле миофибрилл концентрация кальция достигает максимума, создаются все условия для мышечного сокращения (ионы кальция действуют на тропонин → тропонин снимает тропомиозионовую блокаду → миозин взаимодействует с актином → гидролиз АТФ → гребковые движения актиновых и миозиновых нитей).
Выход кальция из СПС прекращается сразу вслед за реполяризациией сарколеммы, но миофибриллы пребывают в сокращенном состоянии. Чтобы миофибриллы расслабились, кальции должен обратно вернуться в саркоплазматическую сеть. Но такой транспорт приходится осуществлять вопреки действию огромного концентрационного градиента (в СПС кальция много, в саркоплазме мало). Следовательно, расслабление миофибрилл в миоците скелетных мышц после их сокращения невозможно без участия системы активного транспорта - кальциевой помпы (рисунок 6, В). Ее работа - неотъемлемый элемент сократительного процесса в мышце. Из мембраны СПС выделена Са-активируемая АТФаза, которая служит основным компонентом кальциевого насоса.
Таким образом, электромеханическое сопряжение включает цепь последовательно идущих процессов: распространение ПД по сарколемме -> деполяризация мембран трубочек Т-системы -> открытие кальциевых каналов в мембране СПС -> выход Са 2+ из цистерн СПС в цитозоль мышечного волокна -> замыкание миозиновых мостиков. Расслабление мышцы связано с реполяризацией сарколеммы и последующей активизацией работы кальциевого насоса, локализованного в мембране СПС.

Для обеспечения молекулярного механизма взаимодействия актиновых и миозиновых нитей необходимы как макроэрги, так и ионы кальция. Именно Са 2+ является важнейшим компонентом электромеханического сопряжения.
Сигналом к сокращению скелетной мышцы являются нервные импульсы, поступающие из спинного или головного мозга к нервно-мышечному соединению. Далее следует синаптическая передача с участием ацетилхолина. Потенциалы действия, образовавшиеся на сарколемме распространяются по всей плазматической мембране мышечного волокна. Деполяризация распространяется и на мембраны саркоплазматического ретикулума СПС и открывает в ней потенциалзависимые кальциевые каналы. Через открывшиеся каналы Са 2+ пассивно (в сторону более низкого электрохимического потенциала) устремляется из цистерн СПС в саркоплазму и достигает миофибрилл. Около них создается достаточная для замыкания миозиновых мостиков концентрация ионов кальция. Дальше возникает сокращение (рисунок 5).
Рисунок 5. Последовательность электрических, осмотических и механических процессов в миоците поперечнополосатых мышц: по оси абсцисс - время (мс), по оси ординат - величины реакций (в % к Rmax)
Как сказано выше, для возникновения сокращения скелетной мышцы ионы кальция должны поступить к миофибриллам из саркоплазматической сети. Так называют систему пузырьков и цистерн, отделенных мембранами от остальной саркоплазмы (рисунок 6). СПС занимает примерно 10% объема мышечного волокна, а суммарная площадь ее мембран в миоците приблизительно в 100 раз больше поверхности сарколеммы (мембраны саркомеров). СПС служит кальциевым депо в мышечном волокне - содержание в ней ионов кальция огромно. Следовательно, на мембране СПС поддерживается колоссальный градиент Са 2+ , но в покое она совершенно непроницаема для этого иона.
При получении сигнала к сокращению скелетной мышцы (нервные импульсы → синаптическая передача → потенциалы действия) начинается деполяризация мембран уже саркоплзматического ретикулума. Ионы кальция из саркоплазматического ретикулума начинают выходить в саркоплазму (по механизмам пассивного транспорта по электрохимическому градиенту через кальциевые каналы, рисунок 6, В). Когда возле миофибрилл концентрация кальция достигает максимума, создаются все условия для мышечного сокращения (ионы кальция действуют на тропонин → тропонин снимает тропомиозионовую блокаду → миозин взаимодействует с актином → гидролиз АТФ → гребковые движения актиновых и миозиновых нитей).
Выход кальция из СПС прекращается сразу вслед за реполяризациией сарколеммы, но миофибриллы пребывают в сокращенном состоянии. Чтобы миофибриллы расслабились, кальции должен обратно вернуться в саркоплазматическую сеть. Но такой транспорт приходится осуществлять вопреки действию огромного концентрационного градиента (в СПС кальция много, в саркоплазме мало). Следовательно, расслабление миофибрилл в миоците скелетных мышц после их сокращения невозможно без участия системы активного транспорта - кальциевой помпы (рисунок 6, В). Ее работа - неотъемлемый элемент сократительного процесса в мышце. Из мембраны СПС выделена Са-активируемая АТФаза, которая служит основным компонентом кальциевого насоса.
Таким образом, электромеханическое сопряжение включает цепь последовательно идущих процессов: распространение ПД по сарколемме -> деполяризация мембран трубочек Т-системы -> открытие кальциевых каналов в мембране СПС -> выход Са 2+ из цистерн СПС в цитозоль мышечного волокна -> замыкание миозиновых мостиков. Расслабление мышцы связано с реполяризацией сарколеммы и последующей активизацией работы кальциевого насоса, локализованного в мембране СПС.
Электрические проявления в мышцах при возбуждении приводят к механического сокращению мышцы, т.е. имеет место электромеханическое сопряжение этих процессов.
Сокращение мышцы происходит при поступлении по аксонам двигательных нейронов к нервно-мышечным синапсам возбуждения в виде нервных импульсов (потенциалы действия нервных волокон). Сокращение происходит при взаимодействии толстых и тонких нитей, которое в покое невозможно, так как миозин-связывающие участки молекул актина заблокированы тропомиозином.
Механизм развития сокращения мышцы представлен несколькими этапами.
1. При распространении ПД по мембране миофибриллы на поперечную трубочку, а затем продольную систему клетки – саркоплазматический ретикулум (СПР) происходит высвобождение Са2+ из цистерн СПР.
2. При связывании Са2+ с тропонином ( TnC ) молекула деформируется таким образом, что тропомиозин как бы заталкивается в желобок между двумя цепочками актина, открывая участки для прикрепления миозиновых поперечных мостиков.
4. АДФ и Фн отходят, а на их место присоединяется новая молекула АТФ, что приводит к разрыву связи миозина с актином. После присоединения молекулы АТФ сразу же происходит ее частичный гидролиз и образуется высокоэнергетическое соединение АДФ∙Фн.
Расслабление мышцы вызывается обратным переносом ионов Са2+ посредством кальциевого насоса в каналы саркоплазматического ретикулума. По мере удаления Са2+ из цитоплазмы открытых центров связывания становится все меньше и в конце концов актиновые и миозиновые филламенты полностью рассоединяются; наступает расслабление мышцы.

Передача сигнала на двигательных концевых пластинках обеспечивается благодаря процессам деполяризации мембраны мышечного волокна и проведения возбуждения внутрь волокна через систему Т-трубочек, которые образуют выпячивания, расположенные перпендикулярно к миофибриллам.
Система Т- и L-трубочек в скелетных мышцах отличается очень консервативным строением: по бокам от поперечной трубочки располагаются две продольные трубочки и образуется так называемая триада. В сердечной мышце L-трубочки развиты слабее, и рядом с Т-трубочкой расположена только одна L-трубочка, формируя диаду.
Читайте также: