
Цитозольный механизм действия липофильных гормонов кратко
Обновлено: 02.07.2024
Механизмы действия гормонов. Рецепторы
Во внеклеточных жидкостях гормоны яичника и гипофиза циркулируют в чрезвычайно малых концентрациях, порядка 10-15-10-9 моль/л. Чтобы оказать биологическое воздействие на клетки-мишени, они нуждаются в специальных механизмах распознавания. Клетки-мишени способны различать гормоны в низких концентрациях за счет связанных с клеткой специальных молекул-детекторов, называемых рецепторами.
Гормоны оказывают свое биологическое действие, взаимодействуя с этими высокоаффинными рецепторами, которые в свою очередь запускают одну или несколько эффекторных внутриклеточных систем. Высокая аффинность, специфичность и уровень экспрессии рецептора вместе определяют характер и интенсивность биологического действия гормона.
Каждый рецептор содержит в своем составе по крайней мере два домена — распознающий домен и сигнал-генерирующий домен. Распознающий домен связывается с гормоном, а второй домен генерирует сигнал, который переводит процесс распознавания гормона в процесс выполнения какой-либо внутриклеточной функции.
Такая последовательность связывание-передача сигнала, или взаимодействие рецептор-эффектор, обеспечивает первый этап усиления действия гормона и отличает клеточный рецептор от транспортных белков плазмы, которые связывают гормон, но не генерируют никаких сигналов.
На основании только одной локализации гормональных рецепторов (например, внутриклеточная/ядерная локализация или локализация на поверхности клетки) можно выделить два механизма действия гормонов. Эти механизмы отличаются еще и путями передачи сигнала, или вторичным мессенджером, ответственным за осуществление действия гормона).
Стероидные гормоны, к примеру, взаимодействуют с рецепторами клеточного ядра: будучи липофильными, они легко проникают через клеточную мембрану и связываются с рецепторами, расположенными либо в цитоплазме, либо в ядре клетки. Это в свою очередь изменяет процесс транскрипции гена в ядерном компартменте. Полипептидные гормоны (ЛГ, ФСГ, чХГ, ГнРГ, ингибины и активины) и факторы роста являются гидрофильными и взаимодействуют с поверхностными рецепторами клеточной мембраны.
Они провоцируют вспышку сигнальной активности в клеточной мембране и цитоплазматических компартментах, одновременно оказывая воздействие и на механизмы транскрипции в ядерном компартменте. Рецепторы клеточной поверхности можно разделить на 4 большие группы в зависимости от вторичного мессенджера).
Различают несколько механизмов реализации гормонального сигнала для водорастворимых и водонерастворимых гормонов.
Все гормоны оказывают три конечных эффекта:
изменение количества белков и ферментов за счёт изменения скорости их синтеза;
изменение активности имеющихся в клетки ферментов;
изменение проницаемости клеточных мембран.

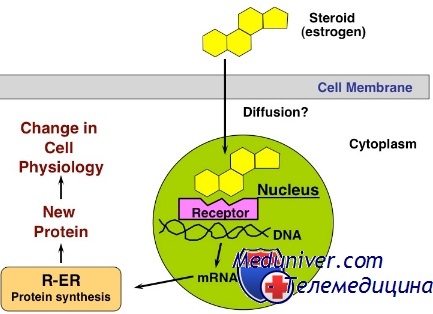
10.3.1. Цитозольный механизм действия гидрофобных (липофильных) гормоновЛипофильные гормоны способны проникать в клетку через клеточную мембрану, поэтому рецепторы для них располагаются внутриклеточно в цитозоле, на митохондриях, на поверхности ядра. Рецепторы гормонов чаще всего включают 2 домена: для связывания с гормоном и для связывания с ДНК. Рецептор при взаимодействии с гормоном изменяет свою структуру, освобождается от шаперонов, в результате чего гормон - рецепторный комплекс приобретает способность проникать внутрь ядра и взаимодействовать с определёнными участками ДНК. Это, в свою очередь, ведёт к изменению скорости транскрипции (синтез РНК), а вследствие этого меняется и скорость трансляции (синтез белка).
10.3.2. Мембранный механизм действия водорастворимых гормонов
Водорастворимые гормоны не способны проникать через цитоплазматическую мембрану. Рецепторы для данной группы гормонов располагаются на поверхности клеточной мембраны. Поскольку гормоны не проходят внутрь клеток, между ними и внутриклеточными процессами необходим вторичный посредник, который передаёт гормональный сигнал внутрь клетки. В качестве вторичных посредников могут служить инозитолсодержащие фосфолипиды, ионы кальция, циклические нуклеотиды.
10.3.2.1. Циклические нуклеотиды - цАмф, цГмф - вторичные посредники
Гормон взаимодействует с рецептором и образует гормон - рецепторный комплекс, в котором меняется конформация рецептора. Это, в свою очередь, изменяет конформацию мембранного ГТФ - зависимого белка (G-белка) и ведёт к активации мембранного фермента аденилатциклазы, который переводит АТФ в цАМФ.


.
10.3.2.2. Ионы кальция - вторичные посредники
Взаимодействие гормона с рецептором повышает проницаемость кальциевых каналов клеточной мембраны, и внеклеточный кальций поступает в цитозоль. В клетках ионы Са 2+ взаимодействуют с регуляторным белком кальмодулином. Комплекс кальций-кальмодулин активирует кальцийзависимые протеинкиназы, которые активируют фосфолирирование различных белков и приводят к конечным эффектам.

10.3.2.3. Инозитолсодержащие фосфолипиды -вторичные посредники.
Аденилатциклазная мессенджерная система. Наиболее изученным является аденилатциклазный путь передачи гормонального сигнала. В нем задействовано минимум пять хорошо изученных белков:
1) рецептор гормона;
2) фермент аденилатциклаза, выполняющая функцию синтеза цАМФ;
3) G-белок, осуществляющий связь между аденилатциклазой и рецептором;
4) цАМФ-зависимая протеинкиназа, катализирующая фосфорилир-ование внутриклеточных ферментов или белков-мишеней, соответственно изменяя их активность;
5) фосфодиэстераза, которая вызывает распад цАМФ и тем самым прекращает (обрывает) действие сигнала.
Аденилатциклаза представляет собой интегральный белок плазматических мембран, его активный центр ориентирован в сторону цитоплазмы и катализирует реакцию синтеза цАМФ из АТФ.
Протеинкиназа – это внутриклеточный фермент, через который цАМФ реализует свой эффект.
Следует отметить, что в клетках открыт большой класс цАМФ-зависи-мых протеинкиназ, названных протеинкиназами А; они катализируют перенос фосфатной группы на ОН-группы серина и треонина (так называемые серин-треонинкиназы). Другой класс протеинкиназ действует только на ОН-группу тирозина. Однако во всех случаях добавление фосфатной группы вызывает изменение активности белков или кинетические свойства.
Гормон соматостатин, соединяясь со своим специфическим рецептором – ингибиторным G-белком, ингибирует аденилатциклазу и синтез цАМФ, т.е. вызывает эффект, прямо противоположный вызываемому адреналином и глюкагоном.

Механизм действия гормонов цитозольной (цитоплазматической) рецепции.Липофильные гормоны диффундируют через плазматическую мембрану и взаимодействуют со специфическими рецепторами внутри клетки, имеющими сродство к данному гормону (Н). Эти рецепторы (R) находятся или в цитоплазме или в ядре клетки, где соответственно и образуется гормон-рецепторный комплекс (НR). Если он сформирован в цитоплазме, то поступает в ядро и связывается с ДНК, регулируя процесс транскрипции, т.е. синтеза матричной РНК (м-РНК). Аналогично действует гормон-рецепторный комплекс, сформированный непосредственно в ядре. Вновь образованная м-РНК поступает из ядра в цитоплазму, где запускает процесс трансляции, т.е. синтез новых белков-ферментов (рис. 10). Помимо влияния на синтез белков-ферментов гормоны цитозольной рецепции повышают проницаемость клеточных мембран для субстратов. Таким образом, в результате действия гормонов цитозольной рецепции в клетке увеличивается количество ферментов и субстратов, что приводит к определенным биологическим эффектам.
Некоторые гормоны, такие как стероидные гормоны и гормоны щитовидной железы , по свойствам являются гидрофобными. В плазме крови для их транспортировки используются специальные белки-транспортеры. В комплексе с этими белками они не способны взаимодействовать с мембранными рецепторами, но способны отрываться от них и диффундировать через клеточную мембрану внутрь клетки. После перехода в цитозоль гормоны немедленно подхватываются другими белками, которые уже являются рецепторами.
Комплекс гормон-рецептор в некоторых случаях дополнительно модифицируется и активируется. Далее он проникает в ядро, где может связываться с ядерным рецептором . В результате гормон приобретает сродство к ДНК. Связываясь с гормон-чувствительным элементом в ДНК, гормон влияет на транскрипцию определенных генов, изменяет концентрацию РНК и, соответственно, количество белков (транспортных, ферментативных. структурных) в клетке.
Читайте также: