
Цитогенетические карты генов кратко
Обновлено: 04.07.2024
Генетически карты- граф-е изображение групп сцепления с указанием распол-я и расстояния м-у генами.В настоящее время карты групп сцепления построены для многих генетических объектов: от бактериофагов до человека. Осн-е этапы картир-я гена:
1. Опред. группы сцепления в кот. нах. картируемый ген.2. Опред. точной локализации гена в найденной группе сцепления.
Опре-е группы сцепления в к-й может нах-ся картир-й ген начинается с опр-я того, наход. он в аутосоме, либо сцеплен с полом. Это опр-е по рез-м рецепрокных скрещ-й. Если они совпадают, то ген расположен в аутосоме, если различны, то сцеплен с полом.
Гены, расположенные в одной хромосоме, в большинстве случаев наследуются совместно и образуют одну группу сцепления, количество которых, т. о., соответствует у каждого организма гаплоидному числу хромосом + кроссинговер.
Чем дальше друг от друга гены, тем выше вероятность кроссинговера и, следовательно, рекомбинации. Генетический анализ позволяет обнаружить перекрест только при различии гомологичных хромосом по составу генов, что при кроссинговере приводит к появлению новых генных комбинаций. Обычно расстояние между генами на Г. к. х. выражают как % кроссинговера (отношение числа мутантных особей, отличающихся от родителей иным сочетанием генов, к общему количеству изученных особей); единица этого расстояния — морганида — соответствует частоте кроссинговера в 1%.
Г. к. х. составляют для каждой пары гомологичных хромосом. Группы сцепления нумеруют последовательно, по мере их обнаружения. Кроме номера группы сцепления, указывают полные или сокращённые названия мутантных генов, их расстояния в морганидах от одного из концов хромосомы, принятого за нулевую точку, а также место центромеры. Составить Г. к. х. можно только для объектов, у которых изучено большое число мутантных генов. Например, у дрозофилы идентифицировано свыше 500 генов, локализованных в её 4 группах сцепления, у кукурузы — около 400 генов, распределенных в 10 группах сцепления. У менее изученных объектов число обнаруженных групп сцепления меньше гаплоидного числа хромосом. Так, у домовой мыши выявлено около 200 генов, образующих 15 групп сцепления (на самом деле их 20). У человека из ожидаемых 23 групп сцепления (23 пары хромосом) идентифицировано только 10, причём в каждой группе известно небольшое число генов; наиболее подробные карты составлены для половых хромосом. У бактерий, которые являются гаплоидными организмами, имеется одна, чаще всего непрерывная, кольцевая хромосома и все гены образуют одну группу сцепления. При переносе генетического материала из клетки-донора в клетку-реципиент, например при конъюгации, кольцевая хромосома разрывается и образующаяся линейная структура переносится из одной бактериальной клетки в другую (у кишечной палочки в течение 110—120 мин). Искусственно прерывая процесс конъюгации, можно по возникшим типам рекомбинантов установить, какие гены успели перейти в клетку-реципиент. В этом состоит один из методов построения Г. к. х. бактерий, детально разработанных у ряда видов. Ещё более детализированы Г. к. х. некоторых бактериофагов.
Цитологические карты хромосом, схематическое изображение хромосом с указанием мест фактического размещения отдельных генов, полученное с помощью цитологических методов. Ц. к. х. составляют для организмов, для которых обычно уже имеются генетические карты хромосом. Каждое место расположения гена (локус) на генетической карте организма, установленное на основе частоты перекреста участков хромосом (кроссинговера), на Ц. к. х. привязано к определённому, реально существующему участку хромосомы, что служит одним из основных доказательств хромосомной теории наследственности. Для построения Ц. к. х. используют данные анализа хромосомных перестроек (вставки, делеции и др.) и, сопоставляя изменения морфологических признаков хромосом при этих перестройках с изменениями генетических свойств организма, устанавливают место того или иного гена в хромосоме. Пользуясь методом хромосомных перестроек, амер. генетик К. Бриджес составил в 1935 подробную Ц. к. х. плодовой мушки дрозофилы, наиболее полно генетически изученного организма. Гигантские хромосомы насекомых отряда двукрылых оказались самыми удобными для построения Ц. к. х., т.к. наряду с большими размерами обладают чёткой морфологической очерченностью: каждый участок этих хромосом имеет свой определённый и чёткий рисунок, обусловленный характерным чередованием по длине ярко окрашиваемых участков (дисков) и слабо окрашиваемых (междисков). Цитологическими методами легко определить отсутствие участка хромосомы или перенос его в др. место. Сопоставление Ц. к. х. с генетическими показало, что физическое расстояние между генами в хромосомах не соответствует генетическому (видимо, частота кроссинговера неодинакова в разных участках хромосом), поэтому плотность распределения генов на цитологических и генетических картах хромосом различна. Так было установлено важное генетическое явление — неравномерность частот перекреста по длине хромосомы. Линейное расположение генов и их последовательность, установленные генетическими методами, подтверждаются Ц. к. х. Современные методы цитологии и генетики позволяют построить Ц. к. х. многих организмов, в том числе человека.
Цитогенетический метод изучения наследственности человека. Хромосомные болезни. Наследование, сцепленное с полом
Цитогенетический метод
Классификация хромосом человека
Генетические карты
Хромосомные болезни (синдромы)
Кариотипы, кариограммы
Тестовые задания
Цитогенетический метод
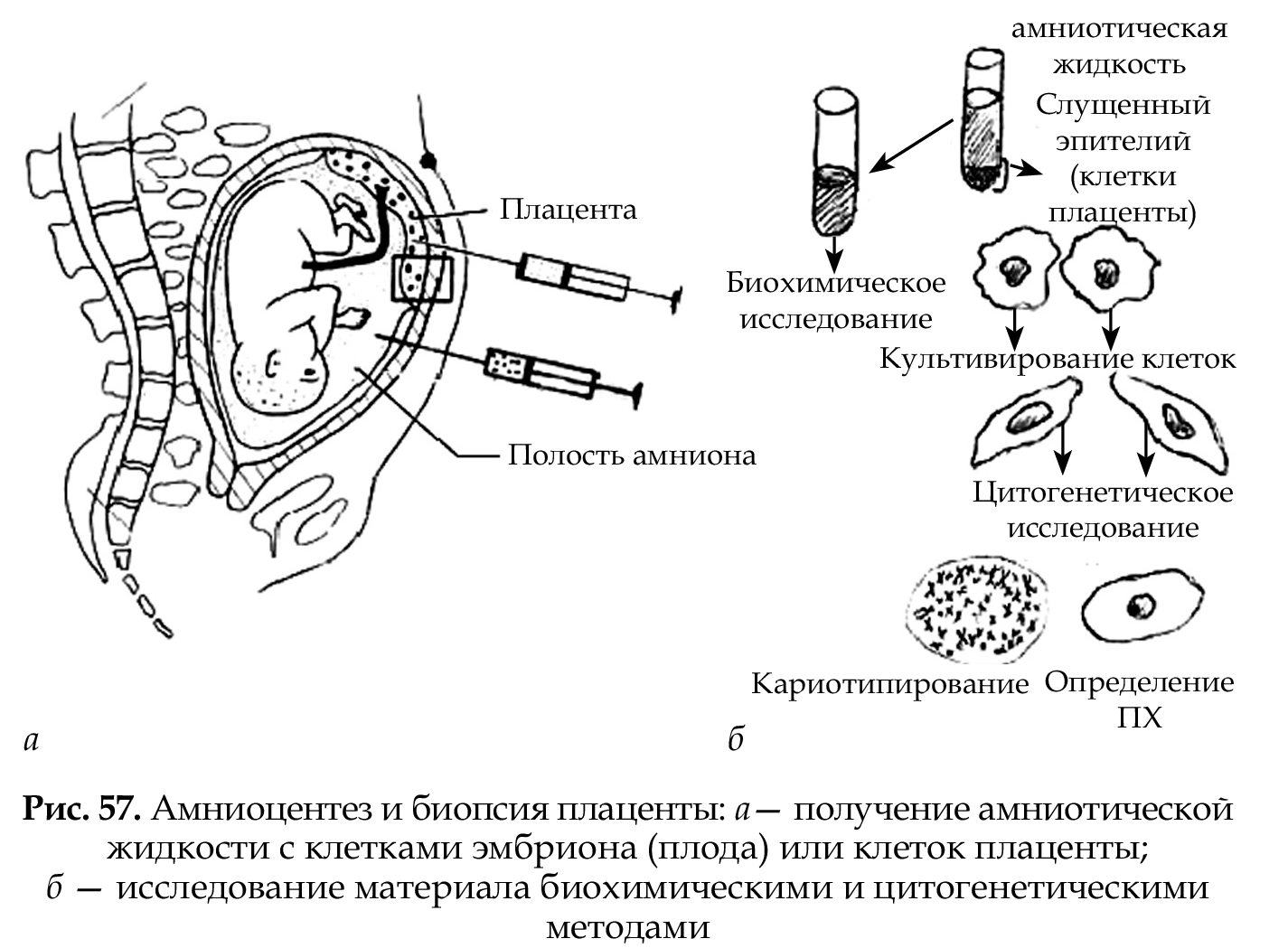
С помощью данного метода можно изучать наследственный материал клетки: совокупность хромосом в целом (кариотипирование) или наличие и количество Х-хромосом (определение полового хроматина — число глыбок полового хроматина или телец Барра). Исследование проводится с помощью светового микроскопа (изготовление и изучение микропрепаратов).
Кариотипирование
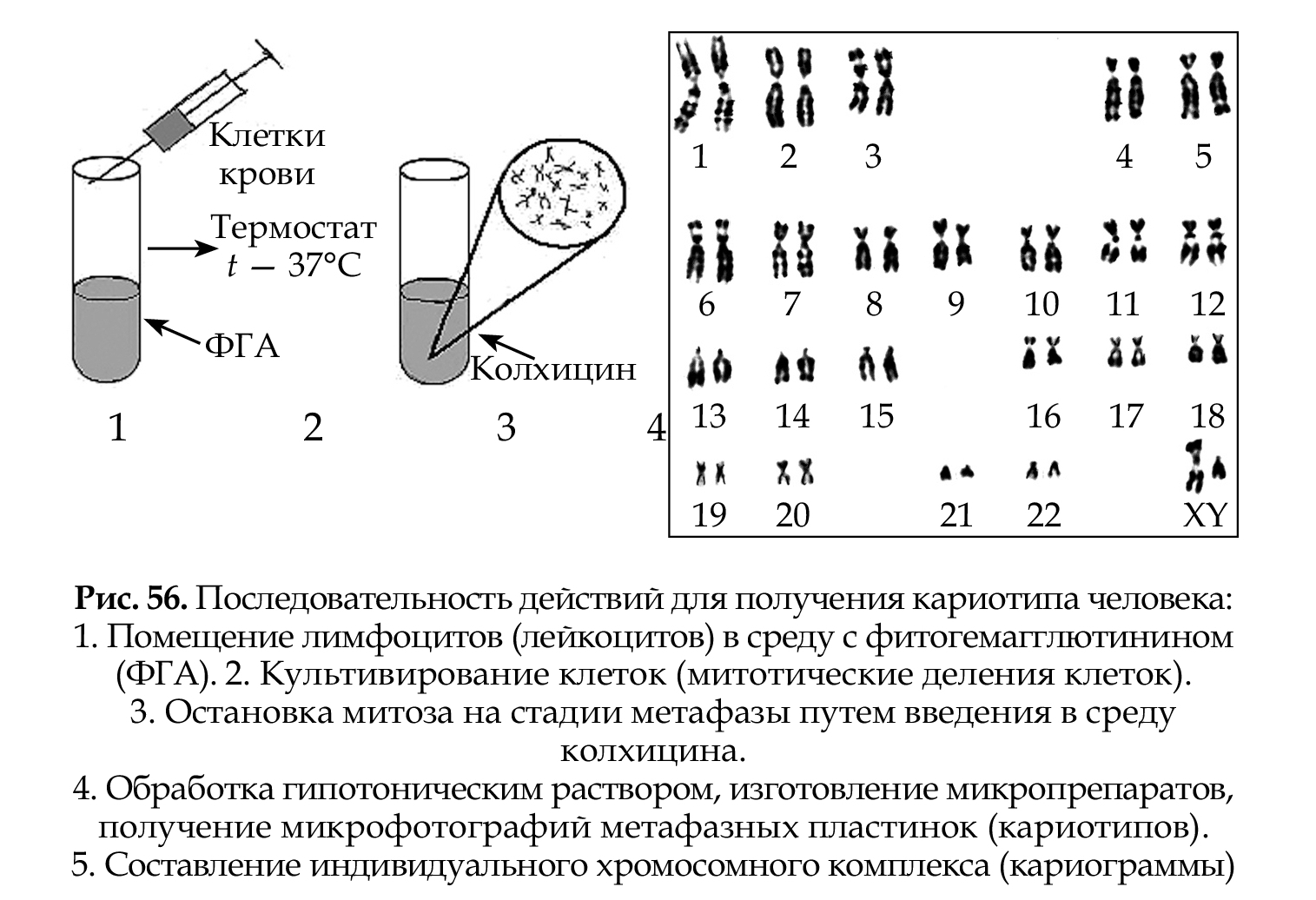
На рис. 56 представлена последовательность действий для получения кариотипа человека.
Половой хроматин
В клетках мужчин (кариотип 46, XY) Х-хромосома всегда выполняет ак-тивную функцию, а у женщин (46, XX) одна Х-хромосома является активной, а другая Х-хромосома находится в неактивном, спирализованном состоянии. Она выявляется в виде компактной темной глыбки в интерфазном ядре соматических клеток нормальных женщин и называется тельцем Барра или половым Х-хроматином. Инактивация одной из двух Х-хромосом происходит в эмбриогенезе, причем закономерности в инактивации отцовской или материнской хромосомы нет (гипотеза Лайон). При любом числе Х-хромосом в активном состоянии будет только одна, следовательно, половой Х-хроматин в норме выявляется только у женщин и отсутствует у мужчин.
Зная число глыбок полового хроматина, можно определить число Х-хромосом по формуле п + 1, где п — число глыбок, а 1 — активная Х-хромосома; и наоборот, зная число Х-хромосом, можно определить число телец Барра по формуле п -1, где п — число Х-хромосом, а 1 — активная Х-хромосома (табл. 12).
У мужчин в норме выявляется Y-половой хроматин (он представляет длинное плечо Y-хромосомы), который обнаруживается с помощью люминисцентной микроскопии и выглядит в виде яркого пятна диаметром 0,3-1 мкм.
Изменение числа глыбок полового хроматина происходит при геномных мутациях (изменение числа X- и Y-хромосом). Определение полового хроматина используется как экспресс-метод при пренатальном и постнатальном определении пола и диагностике хромосомных болезней (см. табл. 12).

Классификация хромосом человека
Хромосомный набор (кариотип) соматической клетки характеризуется формой хромосом, их количеством, размерами, характерными для каждого вида. Препараты хромосом можно приготовить извсех тканей и клеточных суспензий, содержащих делящиеся клетки. Наиболее часто препараты метафазных хромосом готовят из лимфоцитов периферической крови, которые предварительно культивируют в присутствии стимулятора митозов — митогена фитогемаг- глютинина (ФГА), так как клетки периферической крови не делятся.
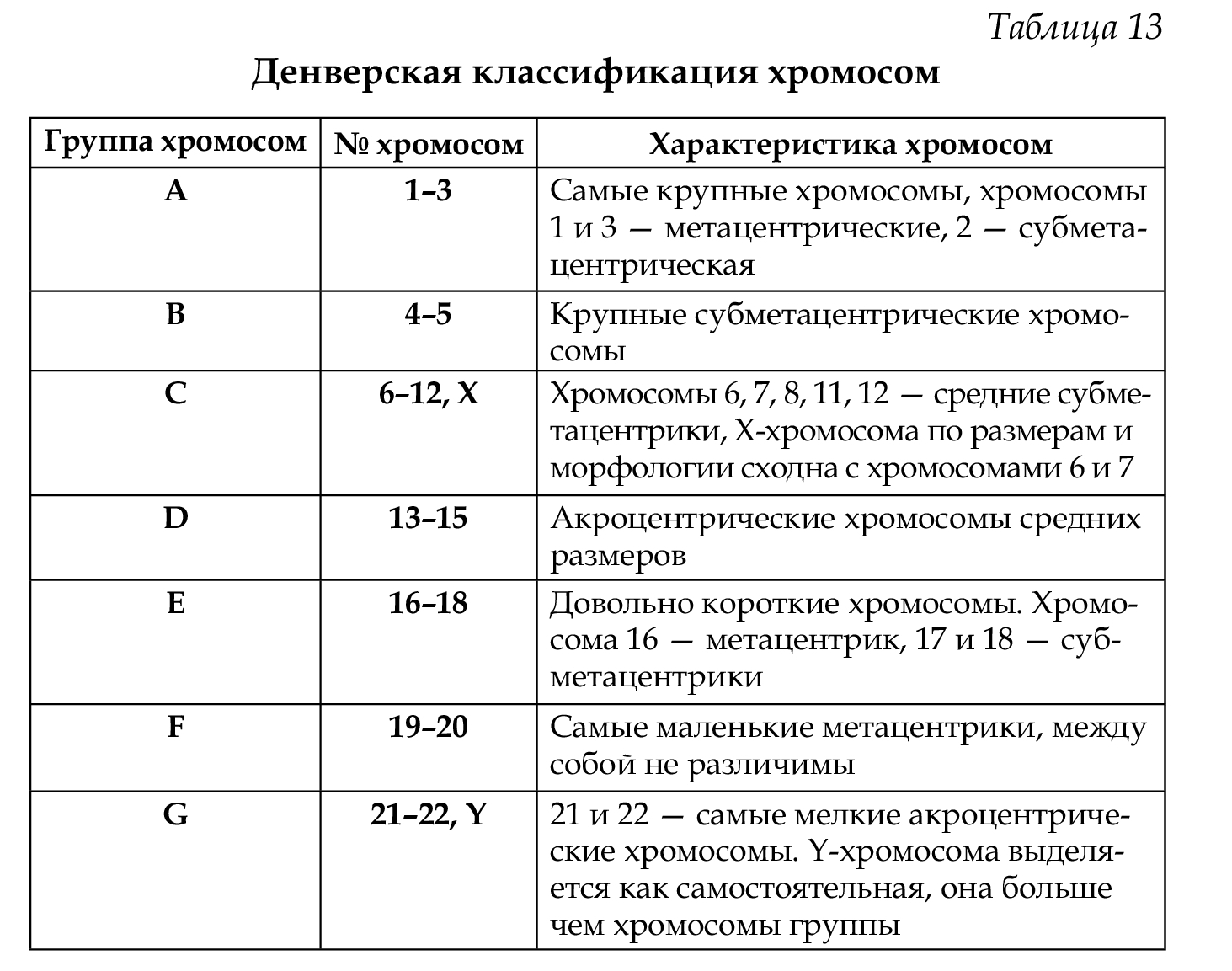
Классификация и номенклатура равномерно окрашенных хромосом (стандартное окрашивание) была разработана на международных совещаниях в Денвере (1960), Лондоне (1963) и Чикаго (1966). Согласно рекомендациям этих конференций, хромосомы располагаются в порядке уменьшения их длины от 1 до 23 хромосомы. Они разделены на 7 групп, которые обозначены буквами английского алфавита от А до G. Все пары хромосом предложено нумеровать арабскими цифрами: группа А 1-3, В 4-5, С 6-12 и Х-хромосома, D 13-15, Е 16-18, F 19- 20, G 21-22, Y-хромосома (табл. 13). При этом хромосомы различных групп хорошо отличаются друг от друга (критерии: размер и форма — метацентрические, субметацентрические, акроцентрические), в то время как внутри группы их сложно различить, за исключением хромосом группы А (рис. 64,65).
Важным параметром является центромерный индекс, который отражает в процентах длину короткого плеча к длине всей хромосомы, например, хромосома № 1 — 48,3%, хромосома № 22 — 22,1%.
Парижская классификация хромосом
В начале 70-х годов XX века был разработан метод дифференциальной окраски хромосом, выявляющий характерную сегментацию, который позволил индивидуализировать каждую хромосому (рис. 58). Различные типы сегментов обозначают по методам, с помощью которых они выявляются наиболее отчетливо (Q-сегменты, G-сегменты, Т-сегменты, S-сегменты). Каждая хромосома человека содержит свойственную только ей последовательность полос, что позволяет идентифицировать каждую хромосому. Хромосомы спирализованы максимально в метафазе, менее спирализованы в профазе и прометафазе, что позволяет выделить большее число сегментов, чем в метафазе.


Классификация хромосом человека
Хромосомная теория наследственности была сформулирована Т. Морганом, основные положения которой сводятся к следующему:
• гены находятся в хромосомах, каждый ген занимает в хромосоме определенное место (локус);
• гены в хромосомах располагаются линейно;
• каждая хромосома представляет собой группу сцепления генов;
• число групп сцепления у каждого вида равно гаплоидному набору хромосом;
• между гомологичными хромосомами в процессе кроссинговера происходит обмен аллельными генами, что приводит к формированию новых сочетаний аллелей в группах сцепления;
• расстояние между генами в хромосоме пропорционально проценту кроссинговера между ними.
Изучение сцепленного наследования явилось основой для составления генетических карт сцепления у разных организмов. Методы классической генетики, цитогенетики и молекулярной генетики позволили подойти к составлению современных генетических карт
Генетическая карта — это система элементов генома, упорядоченная на основе хромосомной принадлежности и взаимного расположения генов в пределах отдельных хромосом, т. е. она определяет принадлежность генов к хромосоме и их расположение относительно друг друга. Возможность ее построения обусловлена линейным характером локализации генов в хромосомах и относительной стабильностью их расположения.
Выделяют следующие генетические карты: карты сцепления, цитологические карты, цитогенетические карты индивидуальных хромосом, рестрикционные и секвенсовые карты. Они различаются единицами измерения (морганида, пара нуклеотидов — п. н., мегабаза — 1 млн оснований) и набором элементов генома.
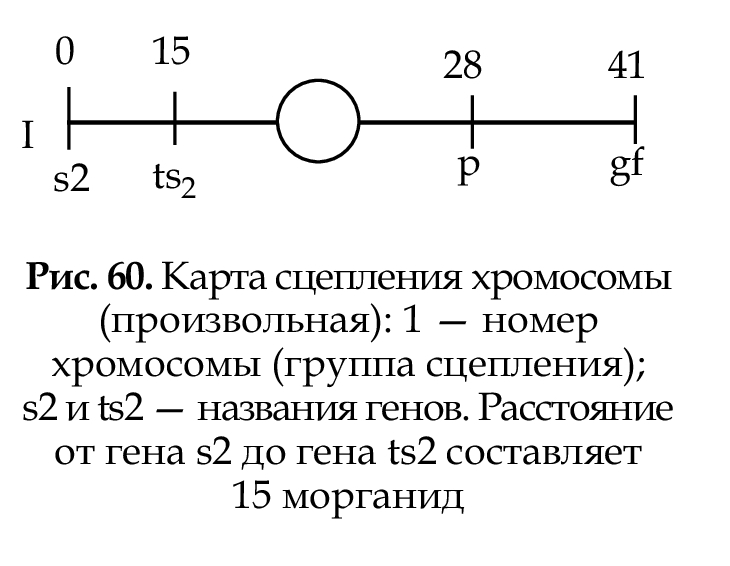
Карта сцепления — схема расположения генов, находящихся в одной группе сцепления, т. е в одной хромосоме (рис. 60). За единицу расстояния между генами принята морганида, которая отражает частоту кроссинговера.Одна морганида — расстояние между генами, при котором кроссинговер происходит в 1% гамет (1 морганида = 1% кроесинговера. При составлении картсцепления указываются: номер хромосомы; полное или сокращенное название генов;
расстояние в морганидах от одного из концов хромосомы, принятого за нулевую точку; место центромеры.
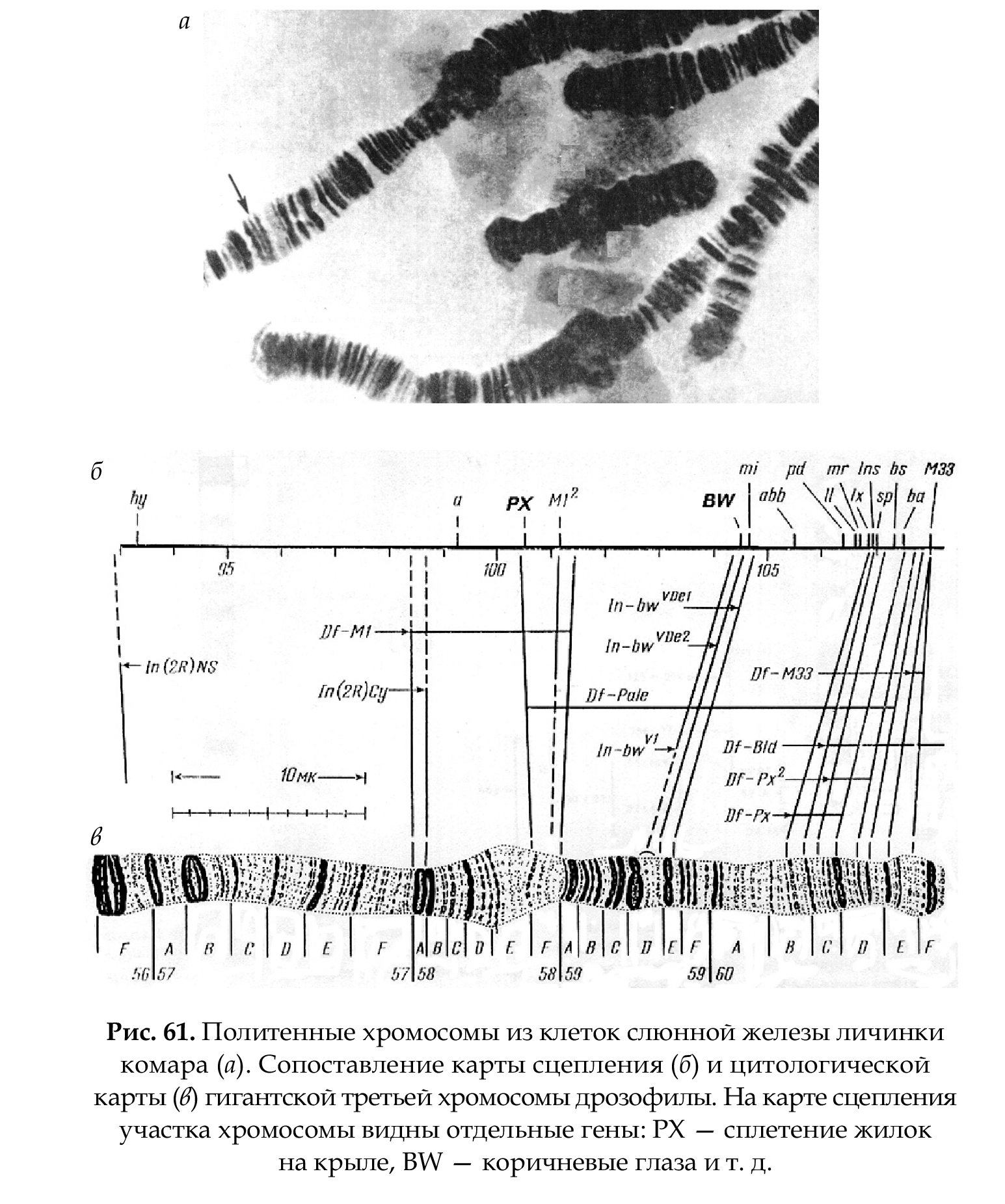
Цитологическая карта составляется на основании из- учения политенных хромосом (рис. 61 а, б), что позволяет сопоставить структуру синтезируемого белка с определенным участком хромосомы (геном), так как транскрибируемый участок определяется под ми- кроскопом в виде пуфа. Это позволяет определить локализацию гена. Изучение политенных хромосом имеет экспери- ментальный характер, так как у человека их нет.
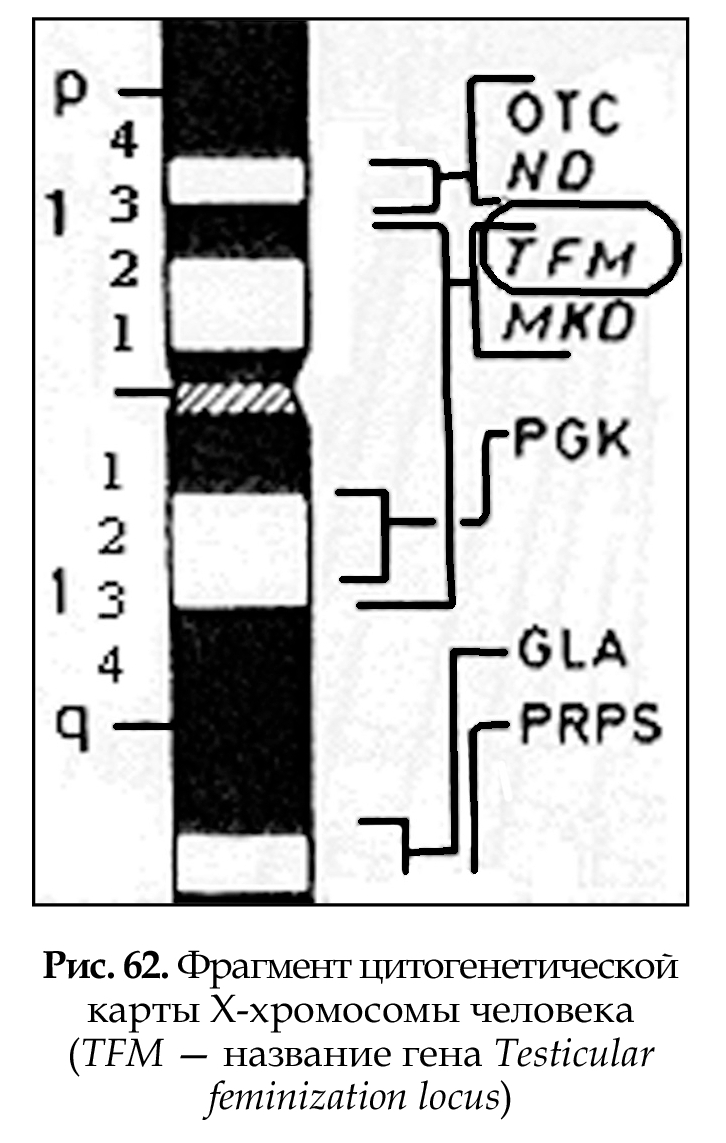
Цитогенетические карты хромосом (рис. 62) составляются на основе дифференциальной окраски (темные и светлые полосы) и картирования генов в отдельных локусах хромосом (основа Парижской классификации). Современные методы окрашивания позволяют выявить до 1000 полос. В среднем на хромосому человека приходится 50 полос. Каждая полоса содержит 3 х 106 пар нуклеотидов, что соответствует нескольким сотням генов (районы и сегменты). Картирование генов в хромосомах является результатом современных молекулярно-генетических методов исследования.
Различные хромосомы и их участки картированы с разной степенью детализации: на Х-хромосоме картировано 400 генов, на хромосоме № 1 — 200 генов. Плотность расположения уже картированных генов в разных хромосомах неравномерна: в 19 хромосоме — 178 генов, в 13 хромосоме — 40 генов, хотя 19 хромосома вдвое меньше, чем 13. На хромосомах № 2 и № 7 примерно одинаковое количество генов, около 175. Каждый картированный ген становится точкой отсчета в геноме, т. е. молекулярным маркером. Генетическое картирование необходимо для определения нуклеотидной последовательности гена и прилегающих к нему участков.
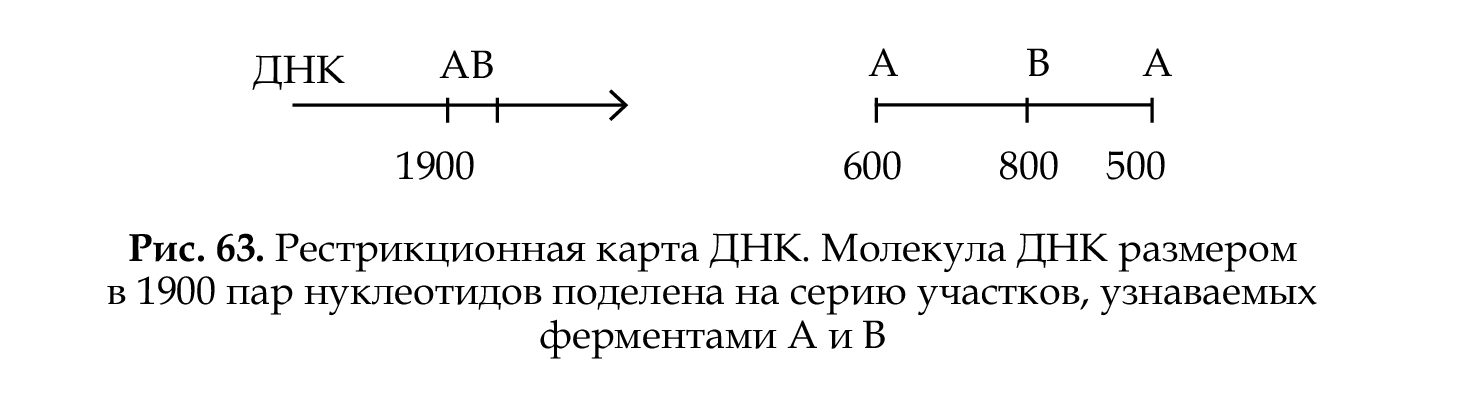
Рестрикционные карты ДНК представляют собой участки ДНК с определенной нуклеотидной последовательностью. Для их получения необходимо выделить ДНК, разрезать в определенных точках рестриктазами, локализовать точки разрыва — сайты расщепления. Карта ДНК, полученная в результате локализации точек разрыва, называется физической рестрикционной картой (рис. 63). Рестрикционная карта ДНК представляет собой линейную последовательность сайтов расщепления, находящихся на определенном расстоянии друг от друга. Расстояние между сайтами рестрикции измеряют в нуклеотидных парах ДНК.
Секвенсовые карты содержат данные о последовательности всех нуклеотидов в целой молекуле ДНК, а не в отдельных ее фрагментах.
Проект геном человека. Генетическое картирование
Таким образом, определение хромосомной локализации искомого гена - генетическое картирование -является первым, ключевым шагом на пути к раскрытию молекулярной основы того или иного наследственного заболевания.

Существует несколько основных методов, позволяющих картировать неизвестный ген в конкретном хромосомном локусе:
а) клинико-генеалогический (простейший и наиболее давний) - основан на анализе наследования признаков в больших родословных; примером может служить установление локализации гена на Х-хромосоме в случае передачи болезни по Х-сцепленному типу;
б) цитогенетический - базируется на ассоциации выявляемых при микроскопии хромосомных перестроек с определенным клиническим фенотипом;
в) метод гибридизации in situ (в том числе его современная модификация - флюоресцентная гибридизация in situ, FISH) - использует специфическую гибридизацию мРНК и кДНК искомого гена с денатурированными хромосомами на метафазных препаратах клетки; г) метод гибридных клеток - основан на анализе совместной сегрегации клеточных признаков и хромосом в клонированных in vitro гибридных соматических клетках [Фогель Ф., Мотульски А., 1990; Gardner E. et al., 1991 ].
Все эти методы нашли свое применение в современной молекулярной генетике, однако они обладают серьезными ограничениями, связанными как с недостаточной разрешающей способностью, так и с существованием жестких предусловий, необходимых для проведения исследования (таких как наличие зондов, доступность селективных систем для отбора гибридных клеток и т.п.). Наиболее мощным, продуктивным и широко используемым в настоящее время методом картирования генов наследственных болезней человека является так называемый linkage-анализ - анализ сцепления искомого гена с набором точно локализованных генетических маркеров [Ott J.,1991; Pulst S., 1999].
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Свидетельство и скидка на обучение каждому участнику
Зарегистрироваться 15–17 марта 2022 г.

Описание презентации по отдельным слайдам:
Картирование генов
Построение генетических и цитологических карт

Картирование генов - определение положения данного гена на какой-либо хромосоме относительно других генов.
Используются три основных группы методов картирования генов
физическое (определение с помощью рестрикционных карт, электронной и световой микроскопии).
Генетическое (определение частот рекомбинаций между генами)
Цитогенетическое (гибридизация in situ, получение монохромосомных клеточных гибридов, делеционной метод и др.)

3
Генетические и физические карты хромосом
Генетическое картирование основано на использовании генетических методов для построения карт, показывающих позиции генов и других последовательностей в геноме.
Генетические методы включают гибридологические эксперименты или, в случае с людьми, генеалогический метод (анализ родословных)

Морган представлял себе гены упорядоченными по длине хромосом, как бусинки в ожерелье
Экспериментальные данные привели его к идее создания генетических карт хромосом
Очевидно, что чем дальше находятся два гена друг от друга, тем больше вероятность разрыва нити, связывающей их, и получения новых сочетаний генов
Стало возможным определить относительное расстояние между генами в хромосоме путем простого расчета процента кроссинговера
4

Частота кроссинговера (расстояние между генами):
число кроссоверных
организмов
= * 100%
общее число потомков

6
Эта частота строго пропорциональна расстоянию между сцепленными генами и измеряется в морганидах
1 морганида соответствует
1% рекомбинантных гамет или генотипов, полученных при анализирующем скрещивании

х 100
Число рекомбинантов
Общее число потомков
Частота рекомбинаций
ЧР =
Серое тело, длинные крылья (BbVv) – 965 (41,5%)
Черное тело, короткие крылья (bbvv) – 944 (41,5%)
Серое тело, короткие крылья (Bbvv) – 206 (8,5%)
Черное тело, длинные крылья (bbVv) – 185 (8,5%)
Всего рекомбинатов - 391 (17%)
Всего потомков - 2300 (100%)
ЧР =
х 100 = 17%
206 + 185
2300
или 17 морганид

Расчёт расстояния между генами

А в С
а В с
А и В – 79 + 14=93 93/521 = 17,9%
В и С – 135+14= 149 149/521=28,6%

10
А. Стертевант в 1913 г. составил первую генетическую карту локализации генов в Х-хромосоме дрозофилы
Генетические карты уже разработаны для дрозофилы, мыши, нейроспоры; для высших растений: кукурузы, риса, ячменя и др.

11
Построение генетической карты на основании частот рекомбинации. Пример показывает реальные эксперименты, выполненные Артуром Стуртевантом на плодовой мушке. Все 4 гена находятся в Х-хромосоме плодовой мушки.Показаны частоты рекомбинации между генами и относительное взаиморасположение генов на карте

Генетические карты (группы сцепления) дрозофилы.


Пример анализа родословной людей.
(A) Родословная показывает наследование генетической болезни в семье двух живых родителей и 6 детей, а также при наличии информации о родителях матери. Аллель болезни является доминантным по отношению к аллелю здоровья. Реальным является определение степени сцепления между геном заболевания и микросателлитом М типированием аллелей для этого микросателлита (M1, M2, и т.д.) у живых членов семьи.
(B) Родословная может быть интерпретирована двумя различными путями: Гипотеза 1 дает низкую частоту рекомбинации и свидетельствует, что ген заболевания сильно сцеплен с микросателлитом М; Гипотеза 2 подтверждает, что ген и микросателлит менее прочно сцеплены
(C) Реконструкция генотипа микросателлита бабушки подтверждает верность Гипотезы 1

15
Физическое картирование использует молекулярно-биологические методы для непосредственного исследования молекул ДНК и построения карт, показывающих позиции определенных последовательностей, в том числе генов

Последовательности, распознаваемые разными рестриктазами
EcoRI
Г ААТТЦ
ЦТТАА Г
SmaI
ЦЦЦ ГГГ
ГГГ ЦЦЦ
ДНК разрезают рестриктазами и подвергают электрофорезу.
Рестрикционная карта - вид физической карты, на которой указаны расстояния между соседними сайтами расщепления ДНК определенной рестриктазой.
Построение рестрикционных карт

17
Опорные точки карт хромосом – гены и ДНК-маркеры
Гены – очень часто используемые маркеры, но они не идеальны. Одна из проблем (особенно для больших геномов позвоночных) состоит в том, что карты, основанные на генах, не очень детальные
Поэтому нужны другие типы маркеров
Опорные точки карт, не являющиеся генами, называются ДНК-маркерами
Основные типы ДНК-маркеров:
полиморфизм длины рестрикционных фрагментов (RFLPs)
полиморфизм длины простой последовательности (SSLPs)
однонуклеотидный полиморфизм (SNPs)

Карта хромосомы 21 и митохондриального генома


20
Методы картирования хромосом человека
метод гибридизации соматических клеток грызунов и человека в культуре ткани
Если изолировать из тела и смешать клетки мыши и человека в культуре, то в результате их слияния можно получить гибридные клетки, содержащие хромосомы одного и другого вида.
Клетки мыши имеют 40 хромосом, а клетки человека - 46. Суммарное число хромосом гибридных клеток должно быть 86, но обычно этого не происходит и чаще всего гибридные клетки содержат обычно от 40 до 50 хромосом.

21
Пример показывает как стабильные человек-мышь гибридные соматические клетки могут получаться применением ПЭГ
По непонятным причинам хромосомы человека избирательно утрачиваются первичным продуктом слияния
Происходящая случайно утрата человеческих хромосом приводит к образованию большого разнообразия гибридных клеток по набору хромосом человека
Эти клетки могут быть клонированными для получения отдельных клеточных линий со специфическим набором хромосом человека
Идентификация хромосом человека может проводиться методами, базирующимися на ПЦР с использованием хромосом-специфических маркеров

22
В гибридных клетках человек-мышь, полученных в результате слияния анеуплоидных клеток мыши и диплоидных эмбриональных фибробластов человека, 75-95% человеческих хромосом утрачиваются в процессе культивирования, причем их утрата носит случайный характер
Среди множества разнообразных гибридов всегда найдется клетка, сохранившая ту или иную хромосому человека
В гибридных клетках хромосомы функционируют, регулируя синтез соответствующих белков

23
После размножения этой клетки можно провести анализ ферментов, активность которых связана с наличием именно данной хромосомы
Использование методов дифференциального окрашивания хромосом позволяет связать гены с определенными локусами хромосом, так как в гибридных клетках довольно часты хромосомные разрывы, перестройки, присутствие не целых хромосом, а отдельных фрагментов


Пример анализа родословной людей.
(A) Родословная показывает наследование генетической болезни в семье двух живых родителей и 6 детей, а также при наличии информации о родителях матери. Аллель болезни является доминантным по отношению к аллелю здоровья. Реальным является определение степени сцепления между геном заболевания и микросателлитом М типированием аллелей для этого микросателлита (M1, M2, и т.д.) у живых членов семьи. (B) Родословная может быть интерпретирована двумя различными путями: Гипотеза 1 дает низкую частоту рекомбинации и свидетельствует, что ген заболевания сильно сцеплен с микросателлитом М; Гипотеза 2 подтверждает, что ген и микросателлит менее прочно сцеплены
(C) Реконструкция генотипа микросателлита бабушки подтверждает верность Гипотезы 1
Читайте также: