
Борозда деления это кратко
Обновлено: 18.05.2024
Как Вы уже знаете, в результате оплодотворения яйцеклетки образуется зигота, способная к дальнейшему развитию. Деление зиготы называют дроблением. Дробление – это многократное деление зиготы после оплодотворения, в результате которого образуется многоклеточный зародыш.
Зигота делится очень быстро, клетки уменьшаются в размерах и не успевают расти. Поэтому зародыш не увеличивается в объеме. Клетки, образующиеся в результате дробления зиготы, называются бластомерами, а перетяжки, отделяющие их друг от друга, называются бороздами дробления.
По направлению различают следующие борозды дробления: меридиональные – это борозды, которые делят зиготу от анимального к вегетативному полюсу; экваториальная борозда разделяет зиготу по экватору; широтные борозды проходят параллельно экваториальной борозде; тангенциальные борозды проходят параллельно поверхности зиготы.
Экваториальная борозда всегда одна, а меридиональных, широтных и тангенциальных может быть много. Направление борозд дробления всегда определяется положением веретена деления.
Дробление всегда проходит по определенным правилам:
Первое правило отражает местоположение веретена дробления в бластомере, а именно:
– веретено дробления располагается в сторону наибольшей протяженности цитоплазмы, свободной от включений.
Второе правило отражает направление борозд дробления:
– борозды дробления проходят всегда перпендикулярно веретену деления.
Третье правило отражает скорость прохождения борозд дробления:
– скорость прохождения борозд дробления обратно пропорционально количеству желтка в яйцеклетке, т.е. в той части клетки, где желтка мало, борозды будут проходить с большей скоростью, а в той части, где желтка больше, скорость прохождения борозд дробления замедляется.
Дробление зависит от количества и местоположения желтка в яйцеклетке. При небольшом содержании желтка дробится вся зигота, при значительном количестве дробится только часть зиготы, свободная от желтка. В связи с этим яйцеклетки разделяют на голобластические (дробящиеся полностью) и меробластические (с частичным дроблением). Следовательно, дробление зависит от количества желтка и с учетом ряда признаков подразделяется: по полноте охвата процессом материала зиготы на полное и неполное; по отношению размеров образующихся бластомеров на равномерное и неравномерное и по согласованности делений бластомеров – синхронное и асинхронное.
Полное дробление может быть равномерным и неравномерным. Полное равномерное дробление характерно для яйцеклеток с небольшим количеством желтка и его более или менее равномерным расположением в оплодотворённой яйцеклетке. Таким типом дробится яйцеклетка ланцетника. В этом случае первая борозда проходит от анимального к вегетативному полюсу, образуется два бластомера; вторая борозда тоже меридиональная, но проходит перпендикулярно первой, образуются четыре бластомера. Третья – экваториальная, образуются восемь бластомеров. После этого идет чередование меридиональных и широтных борозд дробления. Количество бластомеров после каждого деления увеличивается кратно двум (2; 4; 16; 32 и т.д.). В результате такого дробления образуется шарообразный зародыш, который называется бластулой. Клетки, которые образуют стенку бластулы, называют бластодермой, а полость внутри бластоцелью. Анимальная часть бластулы называется – крышей, а вегетативная часть – дном бластулы.

Полное неравномерное дробление характерно для яйцеклеток со средним содержанием желтка, расположенным в вегетативной части. Такие яйцеклетки характерны для круглоротых и земноводных. При этом типе дробления образуются бластомеры неодинаковых размеров. В анимальном полюсе образуются мелкие бластомеры, которые называются микромерами, а в вегетативном – крупные – макромеры. Первые две борозды, как и у ланцетника, проходят меридионально; третья борозда соответствует экваториальной борозде, но сдвинута от экватора к анимальному полюсу. Поскольку в анимальном полюсе находится свободная от желтка цитоплазма, то здесь дробление происходит быстрее и образуются мелкие бластомеры. В вегетативном полюсе содержится основная масса желтка, поэтому борозды дробления проходят медленнее и образуются крупные бластомеры.
Неполное дробление характерно для телолецитальных и центролецитальных яйцеклеток. В дроблении принимает участие только часть яйца, свободная от желтка. Неполное дробление делится на дискоидальное (костистые рыбы, пресмыкающиеся, птицы) и поверхностное (членистоногие).
Неполным дискоидальным дроблением делятся телолецитальные яйцеклетки, у которых большое количество желтка сконцентрировано в вегетативной части. У этих яйцеклеток безжелтковая часть цитоплазмы в виде зародышевого диска распластана на желтке в анимальном полюсе. Дробление происходит только в области зародышевого диска. Вегетативная часть яйцеклетки, заполненная желтком, участия в дроблении не принимает. Толщина зародышевого диска незначительна, поэтому веретена дробления при первых четырех делениях располагаются горизонтально, а борозды дробления проходят вертикально. Образуется один ряд клеток. После нескольких делений клетки становятся высокими и веретена дробления располагаются в них в вертикальном направлении, а борозды дробления проходят параллельно поверхности яйца. В результате зародышевый диск превращается в пластинку, состоящую из нескольких рядов клеток. Между зародышевым диском и желтком возникает небольшая полость в виде щели, которая аналогична бластоцели.
Неполное поверхностное дробление наблюдается в центролецитальных яйцеклетках с большим количеством желтка в его середине. Цитоплазма в таких яйцеклетках располагается по периферии и незначительная ее часть в центре около ядра. Вся остальная часть клетки заполнена желтком. Через массу желтка проходят тонкие цитоплазматические тяжи, соединяющие периферическую цитоплазму с околоядерной. Дробление начинается с деления ядер, в результате количество ядер увеличивается. Они окружаются тонким ободком цитоплазмы, передвигаются к периферии и располагаются в свободной от желтка цитоплазме. Как только ядра попадают в поверхностный слой, он делится соответственно их количеству на бластомеры. В результате такого дробления вся центральная часть цитоплазмы перемещается к поверхности и сливается с периферической. Снаружи образуется сплошная бластодерма, из которой развивается зародыш, а внутри находится желток. Поверхностное дробление свойственно яйцеклеткам членистоногих.
На характер дробления оказывают влияние и свойства цитоплазмы, которые определяют взаимное расположение бластомеров. По этому признаку выделяют радиальное, спиральное и билатеральное дробление. При радиальном дроблении каждый верхний бластомер располагается точно под нижним (кишечнополостные, иглокожие, ланцетник и др.). При спиральном дроблении каждый верхний бластомер смещен относительно нижнего наполовину, т.е. каждый верхний бластомер располагается между двумя нижними. В этом случае бластомеры располагаются как бы по спирали (черви, моллюски). При билатеральном дроблении через зиготу можно провести только одну плоскость, по обеим сторонам которой будут наблюдаться одинаковые бластомеры (круглые черви, асцидии).
Сейчас же после оплодотворения зигота разделяется на 2 клетки, к-е вновь делятся; образовавшиеся 4 клетки делятся на 8 и т.д. Деления следуют одно за другим настолько быстро, что клетки не успевают расти и становятся все мельче. Т.к. увеличение числа клеток при дроблении сопровождается уменьшением их размеров, общий объем зародыша почти не меняется.
Клетки, возникающие в результате дробления, называются бластомерами, а те перетяжки по к-м они отделяются одна от другой, - бороздами дробления. Последние могут иметь различное направление, и соответственно этому различают борозды:
- меридиональные, к-е проходят от анимального полюса к вегетативному;
- экваториальные, пересекающие зиготу по экватору;
- широтные, параллельные экватору;
- тангенциальные, проходящие параллельно поверхности зиготы.
Правила дробления. Первое правило отражает местоположение веретена в бластомере, а второе – его направление. К этим двум правилам следует добавить еще третье: скорость прохождения борозд дробления всегда обратно пропорциональна кол-ву желтка в клетке.
Дробление у разных многоклеточных протекает различно. Это зависит от особенностей строения яйцеклеток, прежде всего от кол-ва и распределения в них желтка. При отсутствии желтка, при небольшом или даже среднем его кол-ве дробится вся оплодотворенная яйцеклетка. Чем больше в ней желтка, тем сильнее затрудняется прохождение борозд дробления. При значительном кол-ве желтка дробится только часть зиготы, к-я свободна от него. Поэтому различают дробление полное и неполное. Соответственно этому яйца разделяются на голобластические , дробящиеся полностью, и меробластические – с частичным дроблением.
Полное дробление м.б. равномерным и неравномерным. Это также обусловливается кол-вом и расположением желтка в яйцеклетке. Если его мало и он распределяется по всему яйцу, борозды дробления проходят с одинаковой скоростью на всем протяжении клетки и разделяют ее на равные бластомеры. Если желток в яйцеклетке размещается неравномерно, то участки, где его много, дробятся медленнее, чем участки, к-е бедны им. В результате получаются неравные бластомеры: мелкие –в анимальном полушарии (микромеры) и крупные – в вегетативном (макромеры). В таком случае дробление хотя и остается полным, но будет уже неравномерным.
Полное равномерное дробление наблюдается у ланцетника. Его яйцо относится к гомолецитальному типу: оно содержит небольшое кол-во желтка, равномерно распределенного по всей клетке. Как у б-ва животных, яйцеклетка ланцетника слегка сплюснута в направлении полюсов. В связи с такой формой наибольшая протяженность ее находится в горизонтальной плоскости, поэтому согласно правилам дробления 1-е веретено ложится горизонтально, а борозда дробления проходит вертикально. В каждом из образовавшихся бластомеров наибольшая протяженность ЦП сохраняется в горизонтальной плоскости, поэтому 2-я борозда дробления проходит опять меридионально. После 2-го дробления направление наибольшей протяженности ЦП в бластомерах меняется. Соответственно этому веретено дробления становится вертикально, и 3-я борозда проходит в экваториальной плоскости. Теперь уже отделяются 4 верхних (анимальных) бластомера от 4 нижних (вегетативных). 4-е дробление опять меридиональное. После него происходит правильное чередование поперечных (экваториальных и широтных) дроблений с продольными (меридиональными). Кол-во бластомеров увеличивается в отношениях, кратных 2: первые 2→ 4→ 8→ 16→ 32→ 64 и т.д. Бластомеры разных полушарий сохраняют почти одинаковые размеры.
В результате дробления получается многоклеточный зародыш в форме полого шара, к-й называется бластулой. В ней различают стенку, состоящую из большого кол-ва мелких клеток – бластодерму и полость – бластоцель (первичная полость тела). Анимальную часть бластулы называют крышей бластулы, а вегетативную – дном бластулы. Полное и равномерное дробление у ланцетника завершается всегда образованием бластулы, стенка к-й состоит из одного слоя клеток, а полость лежит в центре. Такая бластула называется целобластулой.

Полное неравномерное дробление наблюдается у земноводных, яйца к-х относятся к телолецитальным, со средним кол-вом желтка. Последний распределяется по всему яйцу, но большая часть его сосредоточена всегда в вегетативной половине, где и зерна его крупнее. В оплодотворенном яйце лягушки отчетливо выражена полярная дифференцировка, к-я проявляется в резкой пигментации анимальной половины. Первые две борозды дробления проходят меридионально. Они быстро разделяют анимальную часть, значительно задерживаясь в вегетативной. 3-я борозда, проходя параллельно экваториальной плоскости, разделяет бластомеры поперек. Она располагается ближе к анимальному полюсу. Это происходит потому, что веретено дробления смещается от от вегетативного полушария, где сосредоточена основная часть желтка, к анимальному. Верхние бластомеры получаются меньших размеров, чем нижние. При дальнейшем дроблении анимальные бластомеры дробятся заметно быстрее вегетативных, и поэтому разница между их размерами становится все более резкой.
Неполное дробление сопровождается делением только той части яйца, к-я свободна от желтка; часть, заполненная им, не дробится. По этому типу развиваются телолецитальные яйца (у костистых рыб, птиц, пресмыкающихся) и центролецитальные (у насекомых). Соответственно особенностям строения этих яиц различают дискоидальное и поверхностное дробление.
Дискоидальное дробление наблюдается у костистых рыб, птиц, пресмыкающихся. Яйца этих животных богаты желтком, большое кол-во к-го обусловливает их значительные размеры. Свободная от него ЦП, в виде небольшого участка находится в верхней части яйца и называется зародышевым диском. Т.к. дробится только последний, дробление получило название дискоидального.
Толщина зародышевого диска очень незначительна, он распластан на желтке, и его наибольшая протяженность совпадает с плоскостью, параллельной поверхности яйца. Поэтому веретена при первых 3-х и даже 4-х дроблениях располагаются горизонтально, а борозды дробления проходят вертикально. В результате получается 1 ряд клеток. После нескольких делений они становятся высокими и веретена располагаются в них вертикально, а борозды дробления – параллельно поверхности яйца. В связи с этим в зародышевом диске появляются поверхностные клетки и клетки, расположенные на желтке. Последующие деления проходят в самых различных направлениях, и зародышевый диск превращается в пластинку, состоящую из нескольких рядов клеток. Между диском и желтком возникает небольшая щелевидная полость, к-ю приравнивают к бластоцелю.
Поверхностное дробление наблюдается в центролецитальных яйцах с большим кол-вом желтка в их середине. ЦП в таких яйцах располагается по периферии клетки в центре, около ядра. Через массу желтка проходят тонкие ЦП-кие тяжи, соединяющие периферическую ЦП с околоядерной. Дробление начинается с деления ядра и обособления вокруг возникших ядер ЦП. Число ядер увеличивается. Окруженные ЦП, они постепенно передвигаются к периферии яйцеклетки. Как только ядра попадают в ее поверхностный слой, последний разделяется соответственно их кол-ву на бластомеры. Т.о., в результате дробления вся центр. часть ЦП перемещается к поверхности и сливается с периферической. Образуется сплошная бластодерма, из к-й развивается зародыш. Поверхностное дробление свойственно почти исключительно яйцам членистоногих.
После оплодотворения яйцеклетка начинает делиться. Многократное деление зиготы называется дроблением. Оно приводит к образованию многоклеточного зародыша.
Сейчас же после оплодотворения зигота разделяется на 2 клетки, к-е вновь делятся; образовавшиеся 4 клетки делятся на 8 и т.д. Деления следуют одно за другим настолько быстро, что клетки не успевают расти и становятся все мельче. Т.к. увеличение числа клеток при дроблении сопровождается уменьшением их размеров, общий объем зародыша почти не меняется.
Клетки, возникающие в результате дробления, называются бластомерами, а те перетяжки по к-м они отделяются одна от другой, - бороздами дробления. Последние могут иметь различное направление, и соответственно этому различают борозды:
- меридиональные, к-е проходят от анимального полюса к вегетативному;
- экваториальные, пересекающие зиготу по экватору;
- широтные, параллельные экватору;
- тангенциальные, проходящие параллельно поверхности зиготы.
Правила дробления. Первое правило отражает местоположение веретена в бластомере, а второе – его направление. К этим двум правилам следует добавить еще третье: скорость прохождения борозд дробления всегда обратно пропорциональна кол-ву желтка в клетке.
Дробление у разных многоклеточных протекает различно. Это зависит от особенностей строения яйцеклеток, прежде всего от кол-ва и распределения в них желтка. При отсутствии желтка, при небольшом или даже среднем его кол-ве дробится вся оплодотворенная яйцеклетка. Чем больше в ней желтка, тем сильнее затрудняется прохождение борозд дробления. При значительном кол-ве желтка дробится только часть зиготы, к-я свободна от него. Поэтому различают дробление полное и неполное. Соответственно этому яйца разделяются на голобластические , дробящиеся полностью, и меробластические – с частичным дроблением.
Полное дробление м.б. равномерным и неравномерным. Это также обусловливается кол-вом и расположением желтка в яйцеклетке. Если его мало и он распределяется по всему яйцу, борозды дробления проходят с одинаковой скоростью на всем протяжении клетки и разделяют ее на равные бластомеры. Если желток в яйцеклетке размещается неравномерно, то участки, где его много, дробятся медленнее, чем участки, к-е бедны им. В результате получаются неравные бластомеры: мелкие –в анимальном полушарии (микромеры) и крупные – в вегетативном (макромеры). В таком случае дробление хотя и остается полным, но будет уже неравномерным.
Полное равномерное дробление наблюдается у ланцетника. Его яйцо относится к гомолецитальному типу: оно содержит небольшое кол-во желтка, равномерно распределенного по всей клетке. Как у б-ва животных, яйцеклетка ланцетника слегка сплюснута в направлении полюсов. В связи с такой формой наибольшая протяженность ее находится в горизонтальной плоскости, поэтому согласно правилам дробления 1-е веретено ложится горизонтально, а борозда дробления проходит вертикально. В каждом из образовавшихся бластомеров наибольшая протяженность ЦП сохраняется в горизонтальной плоскости, поэтому 2-я борозда дробления проходит опять меридионально. После 2-го дробления направление наибольшей протяженности ЦП в бластомерах меняется. Соответственно этому веретено дробления становится вертикально, и 3-я борозда проходит в экваториальной плоскости. Теперь уже отделяются 4 верхних (анимальных) бластомера от 4 нижних (вегетативных). 4-е дробление опять меридиональное. После него происходит правильное чередование поперечных (экваториальных и широтных) дроблений с продольными (меридиональными). Кол-во бластомеров увеличивается в отношениях, кратных 2: первые 2→ 4→ 8→ 16→ 32→ 64 и т.д. Бластомеры разных полушарий сохраняют почти одинаковые размеры.
В результате дробления получается многоклеточный зародыш в форме полого шара, к-й называется бластулой. В ней различают стенку, состоящую из большого кол-ва мелких клеток – бластодерму и полость – бластоцель (первичная полость тела). Анимальную часть бластулы называют крышей бластулы, а вегетативную – дном бластулы. Полное и равномерное дробление у ланцетника завершается всегда образованием бластулы, стенка к-й состоит из одного слоя клеток, а полость лежит в центре. Такая бластула называется целобластулой.
Полное неравномерное дробление наблюдается у земноводных, яйца к-х относятся к телолецитальным, со средним кол-вом желтка. Последний распределяется по всему яйцу, но большая часть его сосредоточена всегда в вегетативной половине, где и зерна его крупнее. В оплодотворенном яйце лягушки отчетливо выражена полярная дифференцировка, к-я проявляется в резкой пигментации анимальной половины. Первые две борозды дробления проходят меридионально. Они быстро разделяют анимальную часть, значительно задерживаясь в вегетативной. 3-я борозда, проходя параллельно экваториальной плоскости, разделяет бластомеры поперек. Она располагается ближе к анимальному полюсу. Это происходит потому, что веретено дробления смещается от от вегетативного полушария, где сосредоточена основная часть желтка, к анимальному. Верхние бластомеры получаются меньших размеров, чем нижние. При дальнейшем дроблении анимальные бластомеры дробятся заметно быстрее вегетативных, и поэтому разница между их размерами становится все более резкой.
Неполное дробление сопровождается делением только той части яйца, к-я свободна от желтка; часть, заполненная им, не дробится. По этому типу развиваются телолецитальные яйца (у костистых рыб, птиц, пресмыкающихся) и центролецитальные (у насекомых). Соответственно особенностям строения этих яиц различают дискоидальное и поверхностное дробление.
Дискоидальное дробление наблюдается у костистых рыб, птиц, пресмыкающихся. Яйца этих животных богаты желтком, большое кол-во к-го обусловливает их значительные размеры. Свободная от него ЦП, в виде небольшого участка находится в верхней части яйца и называется зародышевым диском. Т.к. дробится только последний, дробление получило название дискоидального.
Толщина зародышевого диска очень незначительна, он распластан на желтке, и его наибольшая протяженность совпадает с плоскостью, параллельной поверхности яйца. Поэтому веретена при первых 3-х и даже 4-х дроблениях располагаются горизонтально, а борозды дробления проходят вертикально. В результате получается 1 ряд клеток. После нескольких делений они становятся высокими и веретена располагаются в них вертикально, а борозды дробления – параллельно поверхности яйца. В связи с этим в зародышевом диске появляются поверхностные клетки и клетки, расположенные на желтке. Последующие деления проходят в самых различных направлениях, и зародышевый диск превращается в пластинку, состоящую из нескольких рядов клеток. Между диском и желтком возникает небольшая щелевидная полость, к-ю приравнивают к бластоцелю.
Поверхностное дробление наблюдается в центролецитальных яйцах с большим кол-вом желтка в их середине. ЦП в таких яйцах располагается по периферии клетки в центре, около ядра. Через массу желтка проходят тонкие ЦП-кие тяжи, соединяющие периферическую ЦП с околоядерной. Дробление начинается с деления ядра и обособления вокруг возникших ядер ЦП. Число ядер увеличивается. Окруженные ЦП, они постепенно передвигаются к периферии яйцеклетки. Как только ядра попадают в ее поверхностный слой, последний разделяется соответственно их кол-ву на бластомеры. Т.о., в результате дробления вся центр. часть ЦП перемещается к поверхности и сливается с периферической. Образуется сплошная бластодерма, из к-й развивается зародыш. Поверхностное дробление свойственно почти исключительно яйцам членистоногих.
ОСНОВЫ ЭМБРИОЛОГИИ ЧЕЛОВЕКА
ДРОБЛЕНИЕ И ОБРАЗОВАНИЕ БЛАСТУЛЫ
Дробление — это последовательное деление зиготы без роста образующихся клеток — бластомеров.
Дробление у человека полное, неравномерное, асинхронное.
После первого деления дробления образуются 2 бластомера. Один из них более темный и крупный, другой более мелкий и светлый (см. рис. - А).

Из крупного бластомера развивается зародыш и почти все провизорные органы (соединительная ткань хориона и плодной части плаценты, амнион, желточный мешок, аллантоис). Из мелкого бластомера развивается трофобласт.
В процессе дробления мелкие клетки делятся быстрее крупных. В результате этого мелкие клетки обрастают снаружи крупные (см. рис. - B). Поэтому образующаяся клеточная масса — морула состоит из двух групп клеток (см. рис. - C). Внутри находятся крупные клетки. Их совокупность называется эмбриобластом. Снаружи располагаются мелкие клетки, называемые трофобластом.
В процессе деления клеток трофобласта и эмбриобласта объем морулы увеличивается, а клетки зародыша начинают секретировать жидкость, которая накапливается внутри под трофобластом.
Со временем количество жидкости увеличивается, и внутри зародыша образуется полость, заполненная этой жидкостью, а клетки эмбриобласта оттесняются к периферии и прилипают к трофобласту. Это и есть бластула (см. рис. - D). Такая бластула называется бластоцистой. Она состоит из (см. рис. - D):
1) трофобласта, образующего как бы стенку бластулы;
2) клеток эмбриобласта, располагающихся внутри;
3) полости бластулы, заполненной жидкостью.
Поверхность бластоцисты неровная, так как трофобласт образует выросты. Эти выросты называются первичными ворсинками трофобласта. Они состоят только из клеток самого трофобласта. Трофобласт является первым провизорным органом , образующимся у зародыша человека. Трофобласт в последующем войдет в состав плаценты. Возникновение трофобласта и его первичных ворсинок — это первый этап в развитии плаценты.
С помощью трофобласта происходит имплантация, то есть внедрение зародыша в толщу слизистой оболочки матки (см.рисунок ниже).
Лекция № 13. Способы деления эукариотических клеток: митоз, мейоз, амитоз
Митотический цикл. Митоз
Митоз — основной способ деления эукариотических клеток, при котором сначала происходит удвоение, а затем равномерное распределение между дочерними клетками наследственного материала.
Митоз представляет собой непрерывный процесс, в котором выделяют четыре фазы: профазу, метафазу, анафазу и телофазу. Перед митозом происходит подготовка клетки к делению, или интерфаза. Период подготовки клетки к митозу и собственно митоз вместе составляют митотический цикл. Ниже приводится краткая характеристика фаз цикла.
Интерфаза состоит из трех периодов: пресинтетического, или постмитотического, — G1, синтетического — S, постсинтетического, или премитотического, — G2.
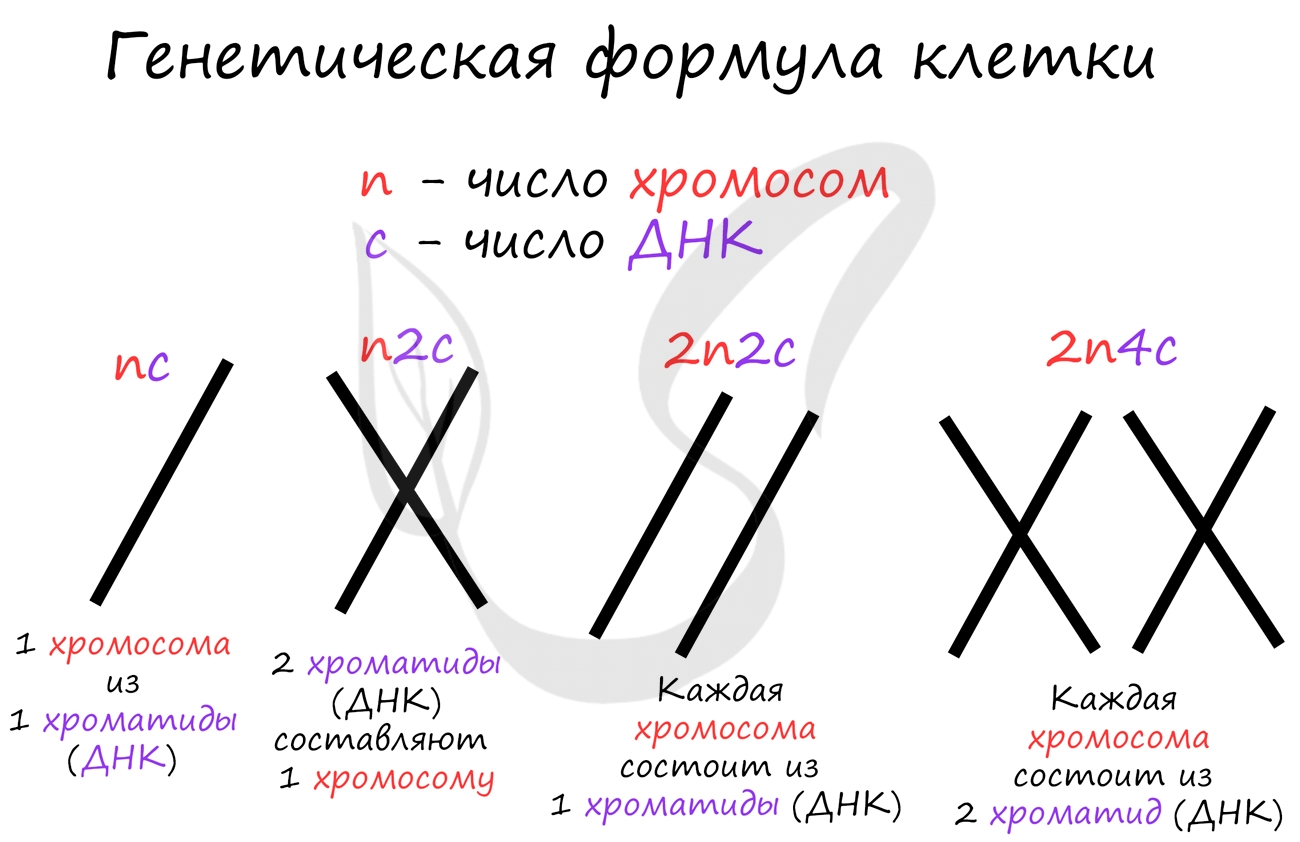
Пресинтетический период (2n 2c, где n — число хромосом, с — число молекул ДНК) — рост клетки, активизация процессов биологического синтеза, подготовка к следующему периоду.
Синтетический период (2n 4c) — репликация ДНК.
Постсинтетический период (2n 4c) — подготовка клетки к митозу, синтез и накопление белков и энергии для предстоящего деления, увеличение количества органоидов, удвоение центриолей.
Метафаза (2n 4c) — выстраивание максимально конденсированных двухроматидных хромосом в экваториальной плоскости клетки (метафазная пластинка), прикрепление нитей веретена деления одним концом к центриолям, другим — к центромерам хромосом.
Анафаза (4n 4c) — деление двухроматидных хромосом на хроматиды и расхождение этих сестринских хроматид к противоположным полюсам клетки (при этом хроматиды становятся самостоятельными однохроматидными хромосомами).
Телофаза (2n 2c в каждой дочерней клетке) — деконденсация хромосом, образование вокруг каждой группы хромосом ядерных мембран, распад нитей веретена деления, появление ядрышка, деление цитоплазмы (цитотомия). Цитотомия в животных клетках происходит за счет борозды деления, в растительных клетках — за счет клеточной пластинки.

Митотический цикл, митоз: 1 — профаза; 2 — метафаза; 3 — анафаза; 4 — телофаза.
Биологическое значение митоза. Образовавшиеся в результате этого способа деления дочерние клетки являются генетически идентичными материнской. Митоз обеспечивает постоянство хромосомного набора в ряду поколений клеток. Лежит в основе таких процессов, как рост, регенерация, бесполое размножение и др.
Мейоз
Мейоз — это особый способ деления эукариотических клеток, в результате которого происходит переход клеток из диплоидного состояния в гаплоидное. Мейоз состоит из двух последовательных делений, которым предшествует однократная репликация ДНК.
Первое мейотическое деление (мейоз 1) называется редукционным, поскольку именно во время этого деления происходит уменьшение числа хромосом вдвое: из одной диплоидной клетки (2n 4c) образуются две гаплоидные (1n 2c).
Интерфаза 1 (в начале — 2n 2c, в конце — 2n 4c) — синтез и накопление веществ и энергии, необходимых для осуществления обоих делений, увеличение размеров клетки и числа органоидов, удвоение центриолей, репликация ДНК, которая завершается в профазе 1.
Профаза 1 подразделяется на стадии: лептотена (завершение репликации ДНК), зиготена (конъюгация гомологичных хромосом, образование бивалентов), пахитена (кроссинговер, перекомбинация генов), диплотена (выявление хиазм, 1 блок овогенеза у человека), диакинез (терминализация хиазм).

Мейоз: 1 — лептотена; 2 — зиготена; 3 — пахитена; 4 — диплотена; 5 — диакинез; 6 — метафаза 1; 7 — анафаза 1; 8 — телофаза 1;
9 — профаза 2; 10 — метафаза 2; 11 — анафаза 2; 12 — телофаза 2.
Купить проверочные работы
и тесты по биологии




Метафаза 1 (2n 4c) — выстраивание бивалентов в экваториальной плоскости клетки, прикрепление нитей веретена деления одним концом к центриолям, другим — к центромерам хромосом.
Анафаза 1 (2n 4c) — случайное независимое расхождение двухроматидных хромосом к противоположным полюсам клетки (из каждой пары гомологичных хромосом одна хромосома отходит к одному полюсу, другая — к другому), перекомбинация хромосом.
Телофаза 1 (1n 2c в каждой клетке) — образование ядерных мембран вокруг групп двухроматидных хромосом, деление цитоплазмы. У многих растений клетка из анафазы 1 сразу же переходит в профазу 2.
Второе мейотическое деление (мейоз 2) называется эквационным.
Интерфаза 2, или интеркинез (1n 2c), представляет собой короткий перерыв между первым и вторым мейотическими делениями, во время которого не происходит репликация ДНК. Характерна для животных клеток.
Профаза 2 (1n 2c) — демонтаж ядерных мембран, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления.
Метафаза 2 (1n 2c) — выстраивание двухроматидных хромосом в экваториальной плоскости клетки (метафазная пластинка), прикрепление нитей веретена деления одним концом к центриолям, другим — к центромерам хромосом; 2 блок овогенеза у человека.
Анафаза 2 (2n 2с) — деление двухроматидных хромосом на хроматиды и расхождение этих сестринских хроматид к противоположным полюсам клетки (при этом хроматиды становятся самостоятельными однохроматидными хромосомами), перекомбинация хромосом.
Телофаза 2 (1n 1c в каждой клетке) — деконденсация хромосом, образование вокруг каждой группы хромосом ядерных мембран, распад нитей веретена деления, появление ядрышка, деление цитоплазмы (цитотомия) с образованием в итоге четырех гаплоидных клеток.
Биологическое значение мейоза. Мейоз является центральным событием гаметогенеза у животных и спорогенеза у растений. Являясь основой комбинативной изменчивости, мейоз обеспечивает генетическое разнообразие гамет.

Амитоз
Амитоз — прямое деление интерфазного ядра путем перетяжки без образования хромосом, вне митотического цикла. Описан для стареющих, патологически измененных и обреченных на гибель клеток. После амитоза клетка не способна вернуться в нормальный митотический цикл.
Клеточный цикл
Клеточный цикл — жизнь клетки от момента ее появления до деления или смерти. Обязательным компонентом клеточного цикла является митотический цикл, который включает в себя период подготовки к делению и собственно митоз. Кроме этого, в жизненном цикле имеются периоды покоя, во время которых клетка выполняет свойственные ей функции и избирает дальнейшую судьбу: гибель или возврат в митотический цикл.
Смотреть оглавление (лекции №1-25)
Митоз и мейоз
Жизненный цикл клетки (клеточный цикл)
С момента появления клетки и до ее смерти в результате апоптоза (программируемой клеточной гибели) непрерывно продолжается жизненный цикл клетки.
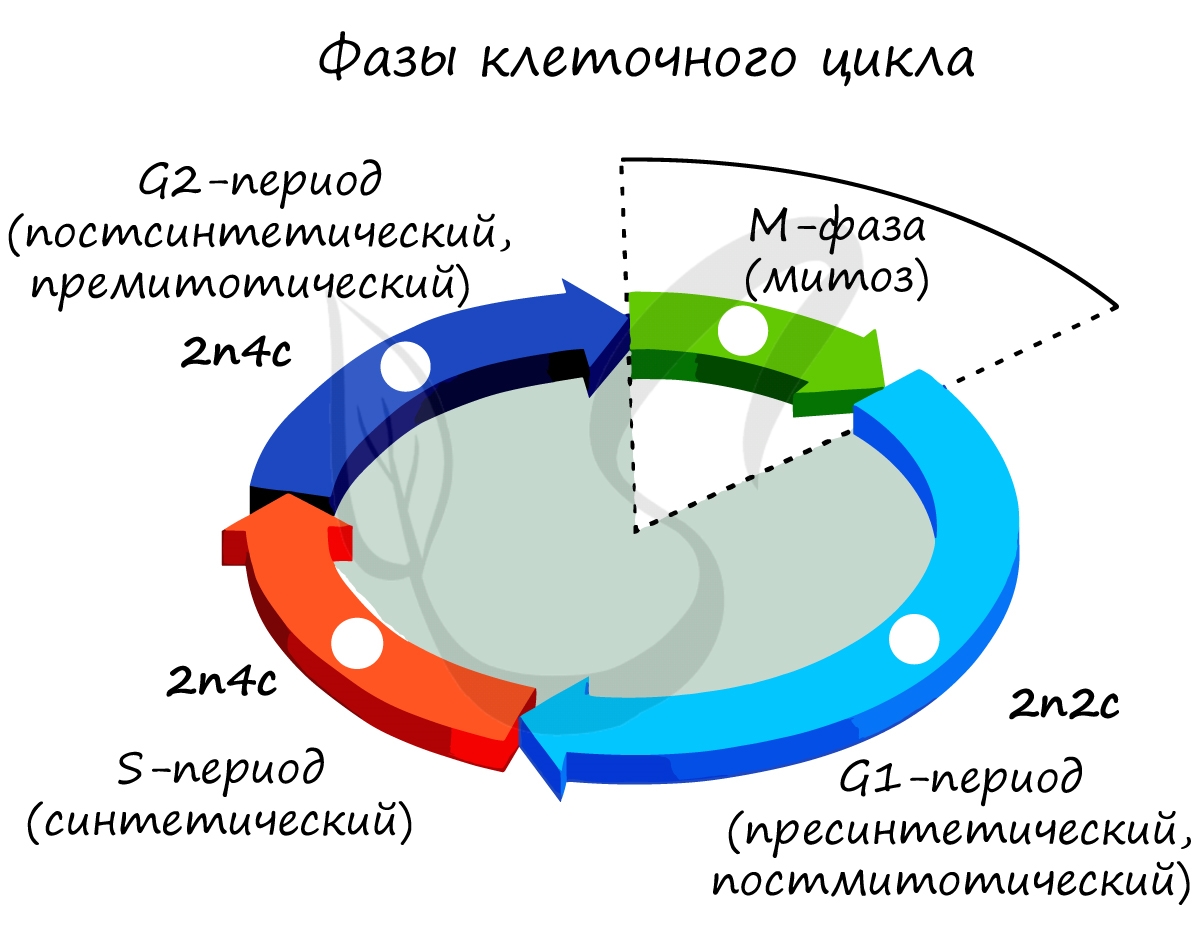
Клеточный цикл включает в себя несколько этапов: деление (митоз), постмитотический (пресинтетический), синтетический, постсинтетический (премитотический) период. Три последних периода составляют интерфазу — подготовку к делению клетки.
Разберем периоды интерфазы более подробно:
-
Пресинтетический (постмитотический) период G1 — 2n2c
Интенсивно образуются рибосомы, синтезируется АТФ и все виды РНК, ферменты, клетка растет.
Синтетический период S — 2n4c
Длится 6-10 часов. Важнейшее событие этого периода — удвоение ДНК, вследствие которого к концу синтетического периода каждая хромосома состоит из двух хроматид. Активно синтезируются структурные белки ДНК — гистоны.
Постсинтетический (премитотический) период G2 — 2n4c
Короткий, длится 2-6 часов. Это время клетка тратит на подготовку к последующему процессу — делению клетки, синтезируются белки и АТФ, удваиваются центриоли, делятся митохондрии и хлоропласты.
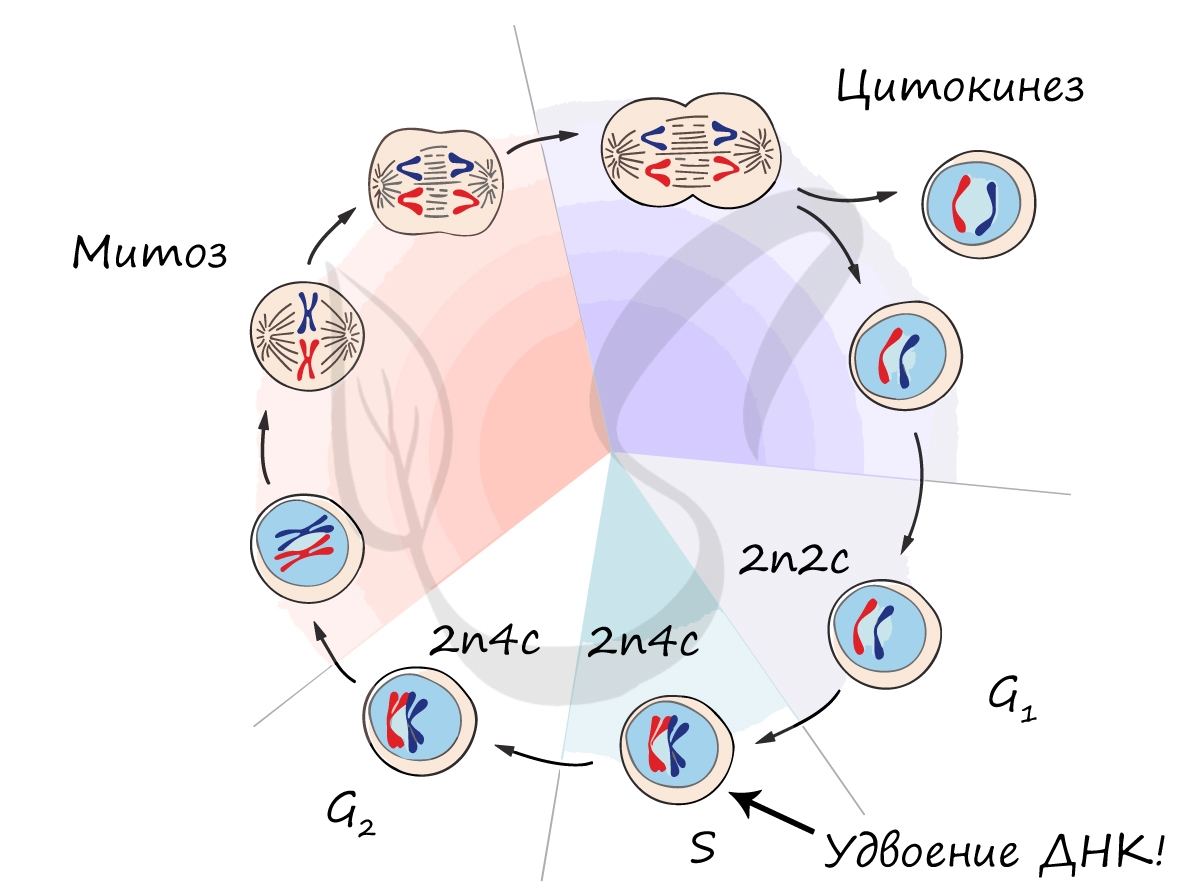
Митоз (греч. μίτος — нить)
Митоз является непрямым способом деления клетки, наиболее распространенным среди эукариотических организмов. По продолжительности занимает около 1 часа. К митозу клетка готовится в период интерфазы путем синтеза белков, АТФ и удвоения молекулы ДНК в синтетическом периоде.
Митоз состоит из 4 фаз, которые мы далее детально рассмотрим: профаза, метафаза, анафаза, телофаза. Напомню, что клетка вступает в митоз с уже удвоенным (в синтетическом периоде) количеством ДНК. Мы рассмотрим митоз на примере клетки с набором хромосом и ДНК 2n4c.
- Бесформенный хроматин в ядре начинает собираться в четкие оформленные структуры — хромосомы — происходит это за счет спирализации ДНК (вспомните мой пример ассоциации хромосомы с мотком ниток)
- Оболочка ядра распадается, хромосомы оказываются в цитоплазме клетки
- Центриоли перемещаются к полюсам клетки, образуются центры веретена деления
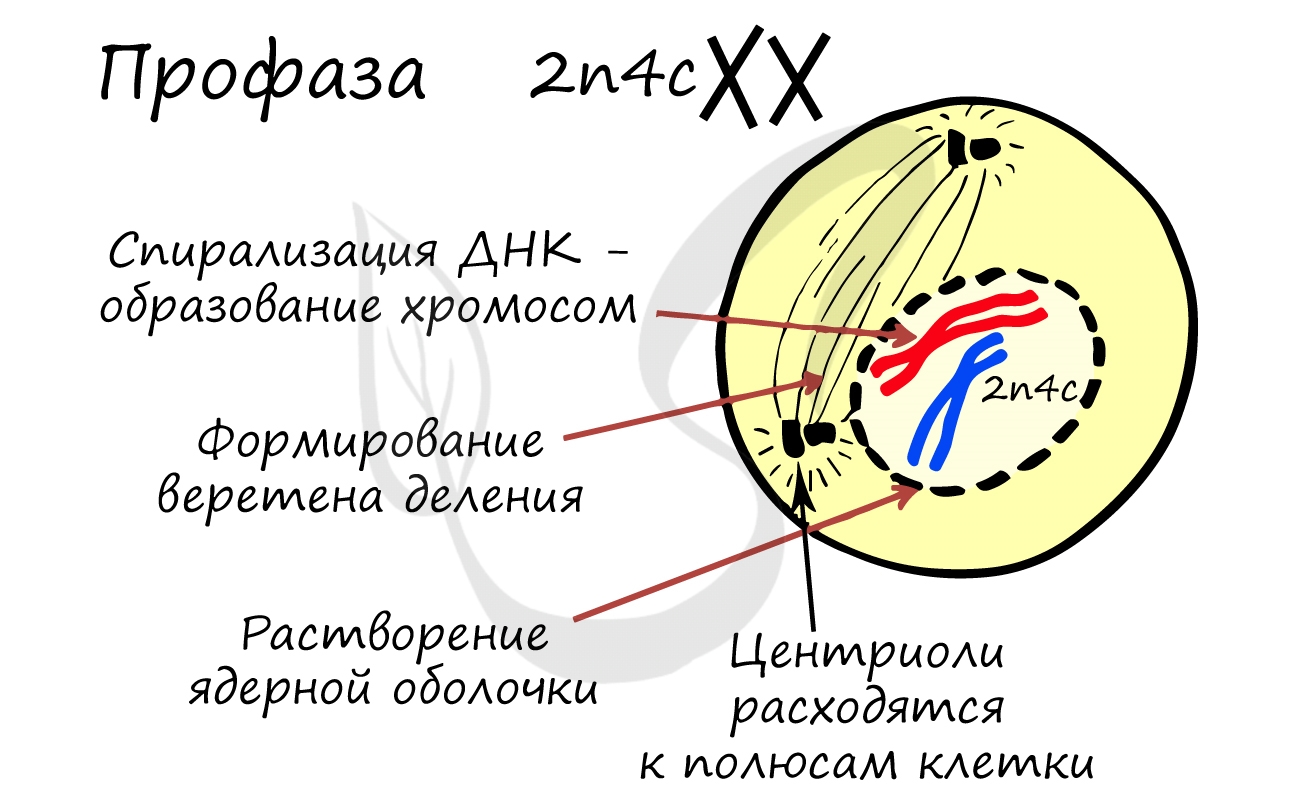
ДНК максимально спирализована в хромосомы, которые располагаются на экваторе клетки. Каждая хромосома состоит из двух хроматид, соединенных центромерой (кинетохором). Нити веретена деления прикрепляются к центромерам хромосом (если точнее, прикрепляются к кинетохору центромеры).
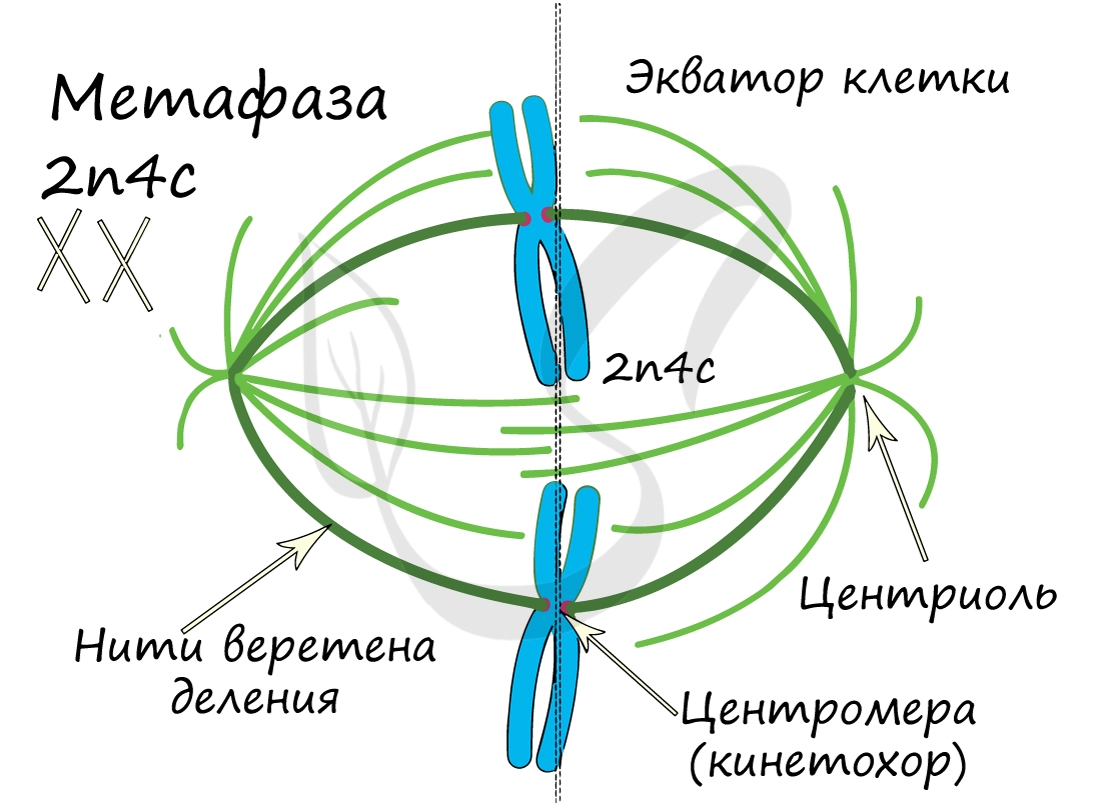
Самая короткая фаза митоза. Хромосомы, состоящие из двух хроматид, распадаются на отдельные хроматиды. Нити веретена деления тянут хроматиды (синоним — дочерние хромосомы) к полюсам клетки.
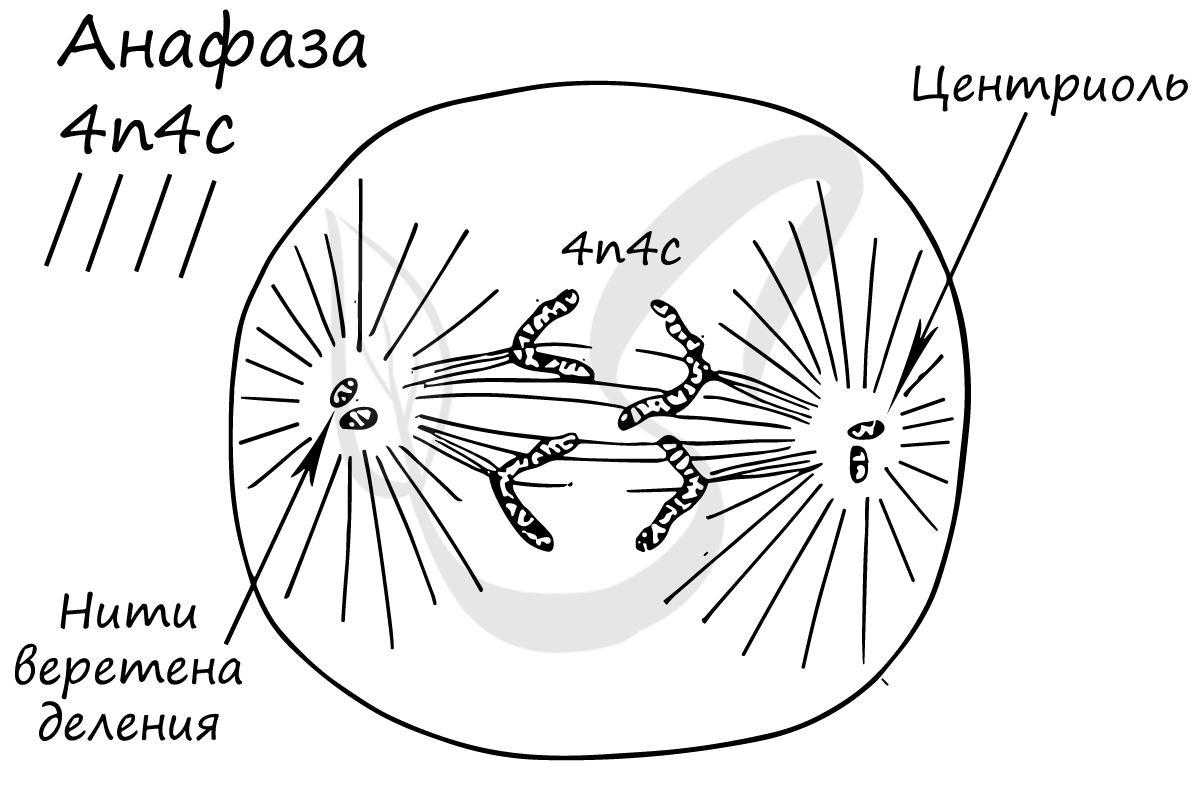
В этой фазе хроматиды (дочерние хромосомы) достигают полюсов клетки.
- Начинается процесс деспирализации ДНК, хромосомы исчезают и становятся хроматином (вспомните ассоциацию про раскрученный моток ниток)
- Появляется ядерная оболочка, формируется ядро
- Разрушаются нити веретена деления
В телофазе происходит деление цитоплазмы — цитокинез (цитотомия), в результате которого образуются две дочерние клетки с набором 2n2c. В клетках животных цитокинез осуществляется стягиванием цитоплазмы, в клетках растений — формированием плотной клеточной стенки (которая растет изнутри кнаружи).
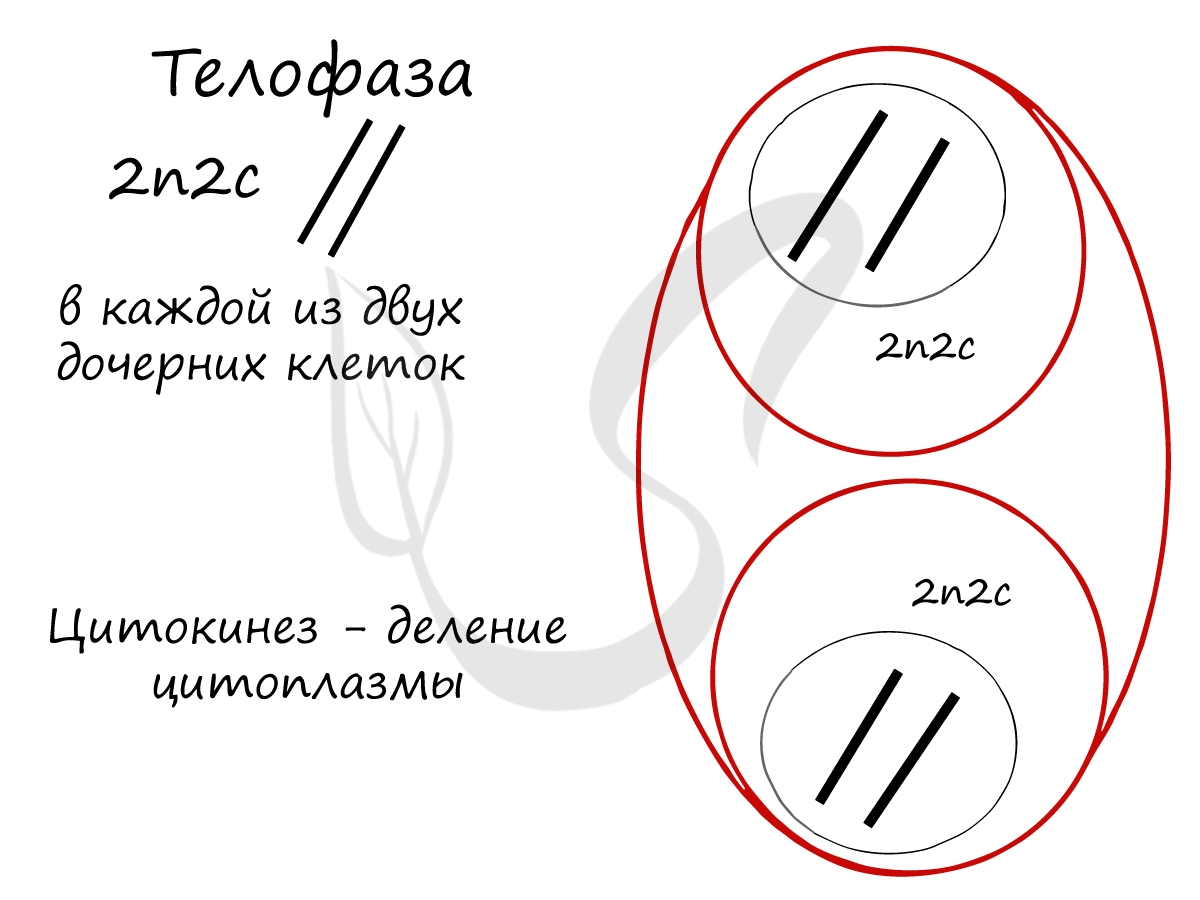
Образовавшиеся в телофазе дочерние клетки 2n2c вступают в постмитотический период. Затем в синтетический период, где происходит удвоение ДНК, после чего каждая хромосома состоит из двух хроматид — 2n4c. Клетка с набором 2n4c и попадает в профазу митоза. Так замыкается клеточный цикл.
Биологическое значение митоза очень существенно:
- В результате митоза образуются дочерние клетки — генетические копии (клоны) материнской.
- Митоз является универсальным способом бесполого размножения, регенерации и протекает одинаково у всех эукариот (ядерных организмов).
- Универсальность митоза служит очередным доказательством единства всего органического мира.
Попробуйте самостоятельно вспомнить фазы митоза и описать события, которые в них происходят. Особенное внимание уделите состоянию хромосом, подчеркните сколько в них содержится молекул ДНК (хроматид).
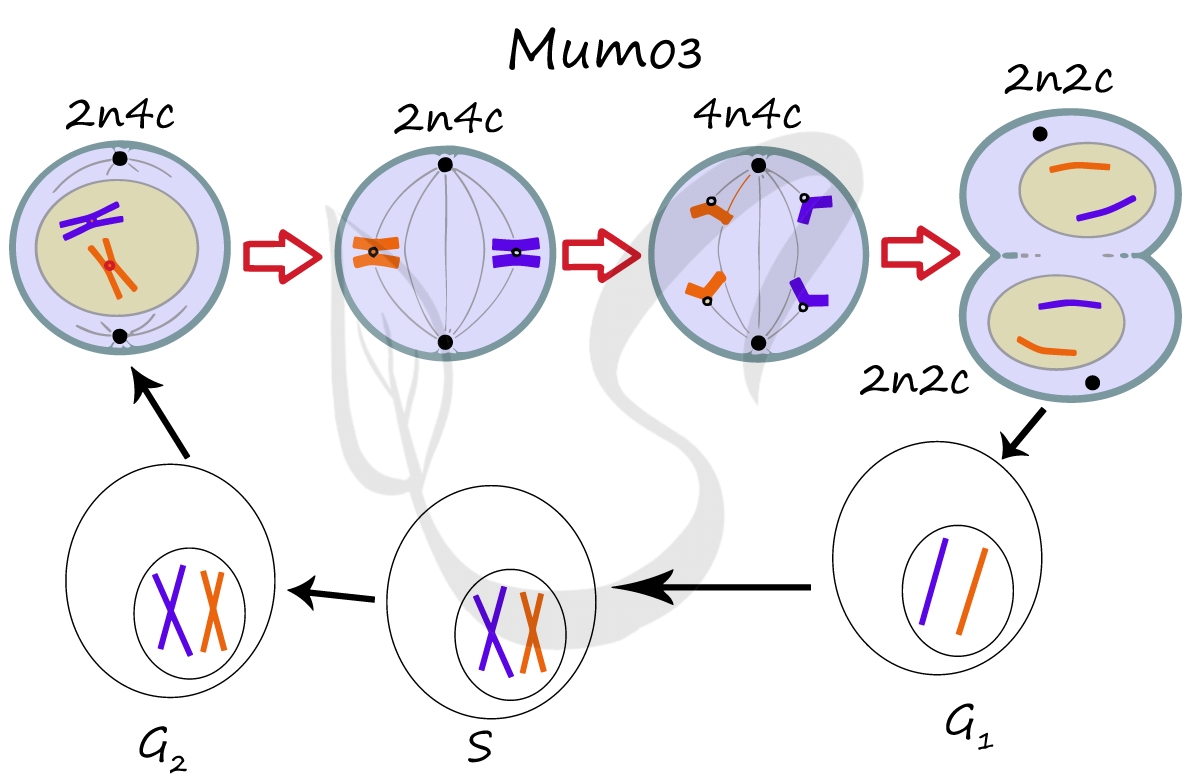
Мейоз
Мейоз (от греч. μείωσις — уменьшение), или редукционное деление клетки — способ деления клетки, при котором наследственный материал в них (число хромосом) уменьшается вдвое. Мейоз происходит в ходе образования половых клеток (гамет) у животных и спор у растений.
В результате мейоза из диплоидных клеток (2n) получаются гаплоидные (n). Мейоз состоит из двух последовательных делений, между которыми практически отсутствует пауза. Удвоение ДНК перед мейозом происходит в синтетическом периоде интерфазы (как и при митозе).
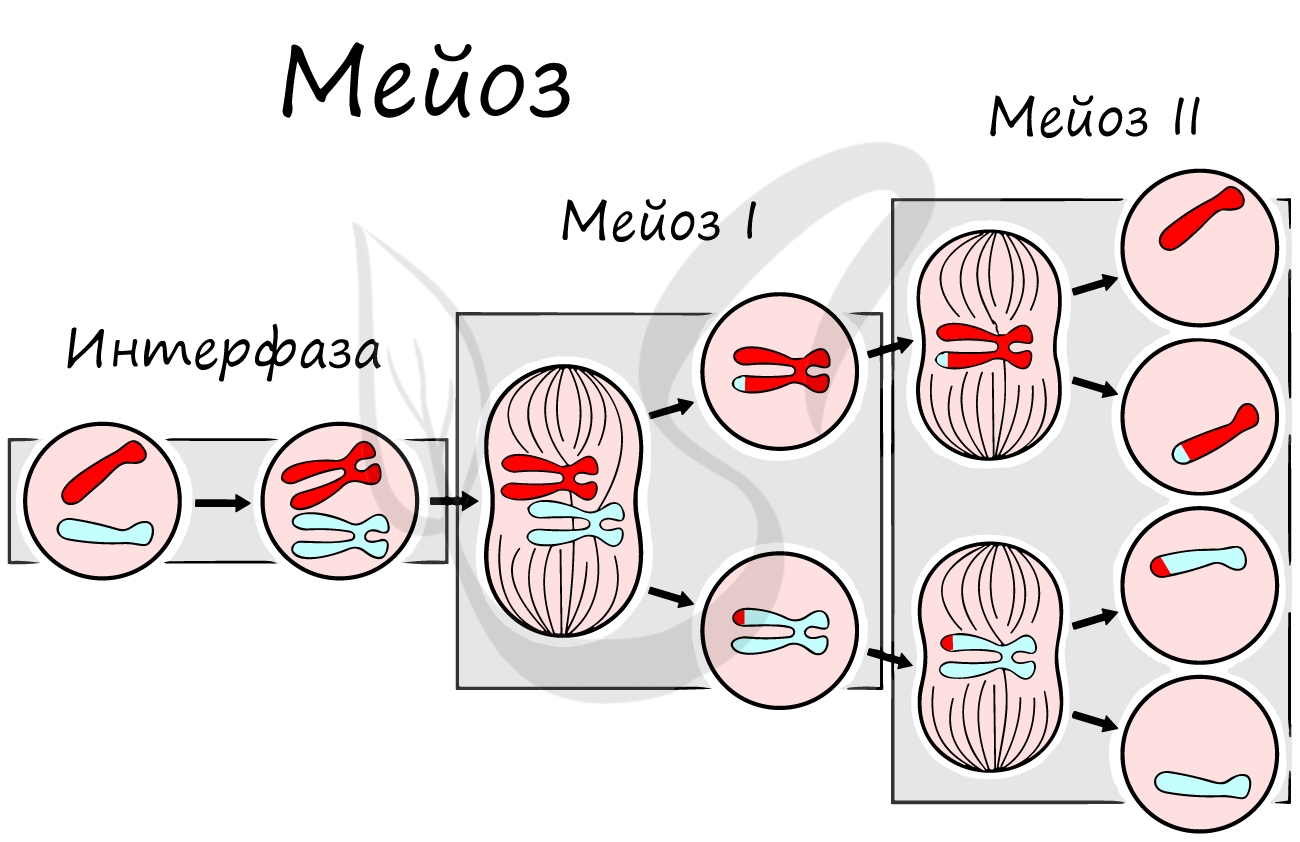
Как уже было сказано, мейоз состоит из двух делений: мейоза I (редукционного) и мейоза II (эквационного). Первое деление называют редукционным (лат. reductio — уменьшение), так как к его окончанию число хромосом уменьшается вдвое. Второе деление — эквационное (лат. aequatio — уравнивание) очень похоже на митоз.
Приступим к изучению первого деления мейоза. За основу возьмем клетку с двумя хромосомами и удвоенным (в синтетическом периоде интерфазы) количеством ДНК — 2n4c.
-
Профаза мейоза I
Помимо типичных для профазы процессов (спирализация ДНК в хромосомы, разрушение ядерной оболочки, движение центриолей к полюсам клетки) в профазе мейоза I происходят два важнейших процесса: конъюгация и кроссинговер.
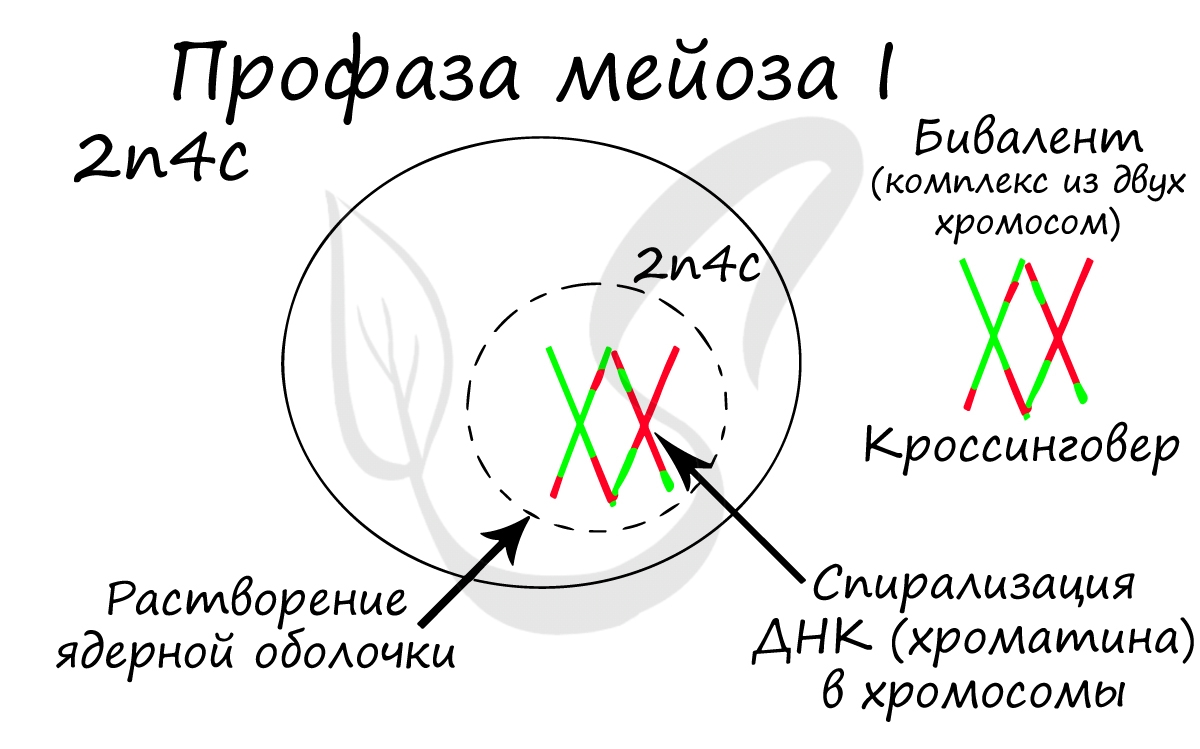
Конъюгация (лат. conjugatio — соединение) — сближение гомологичных хромосом друг с другом. Гомологичными хромосомами называются такие, которые соответствуют друг другу по размерам, форме и строению. В результате конъюгации образуются комплексы, состоящие из двух хромосом — биваленты (лат. bi — двойной и valens — сильный).
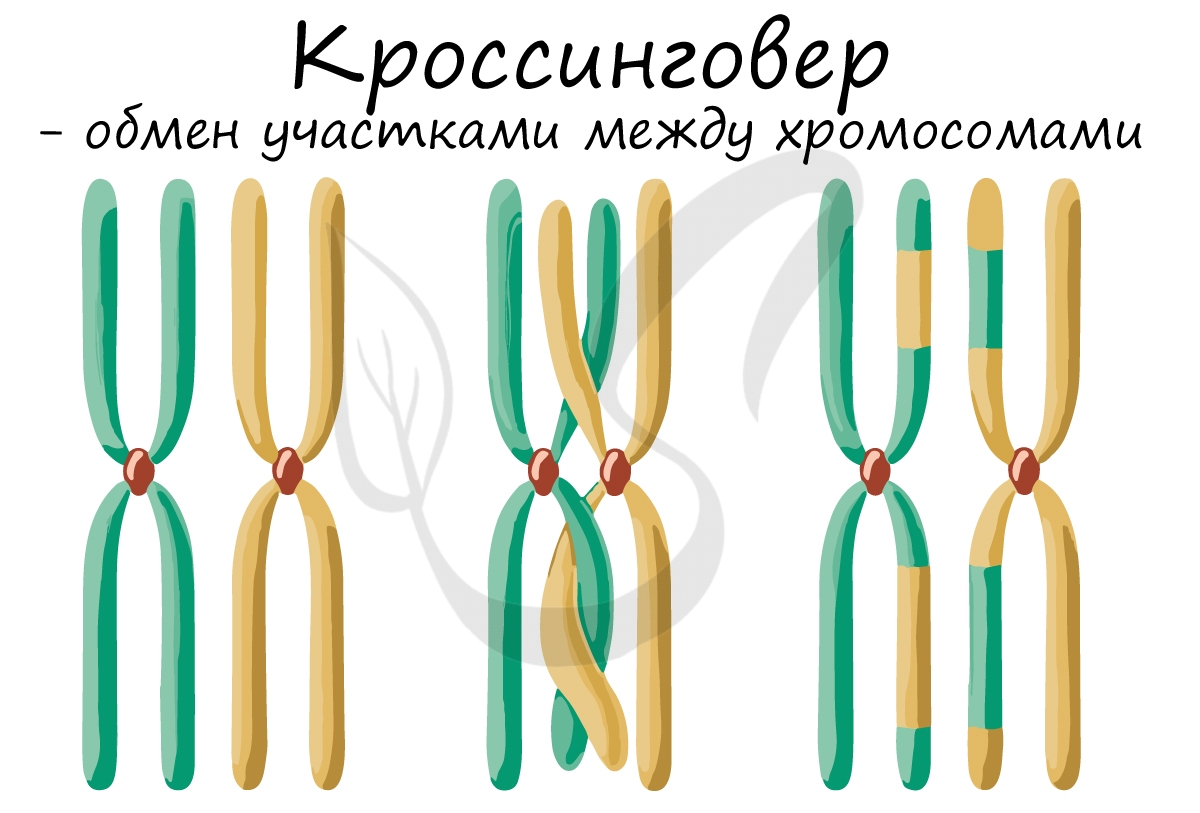
После конъюгации становится возможен следующий процесс — кроссинговер (от англ. crossing over — пересечение), в ходе которого происходит обмен участками между гомологичными хромосомами.
Кроссинговер является важнейшим процессом, в ходе которого возникают рекомбинации генов, что создает уникальный материал для эволюции, последующего естественного отбора. Кроссинговер приводит к генетическому разнообразию потомства.
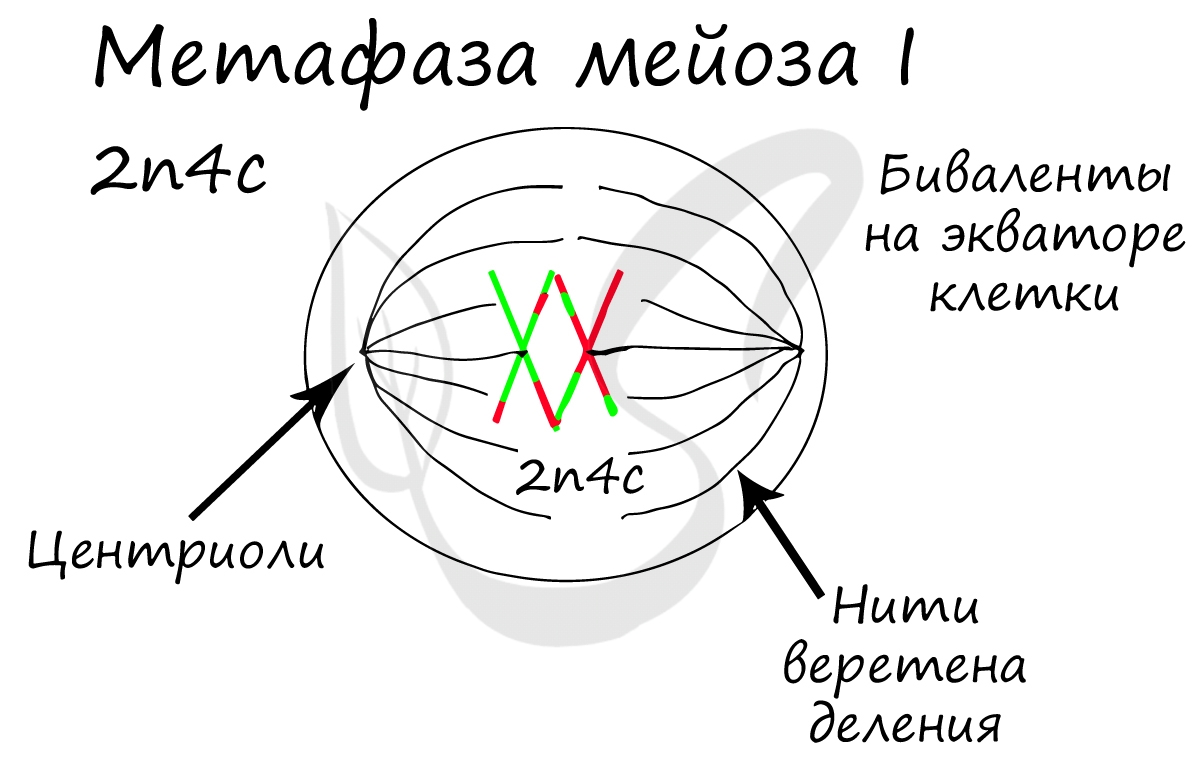
Биваленты (комплексы из двух хромосом) выстраиваются по экватору клетки. Формируется веретено деления, нити которого крепятся к центромере (кинетохору) каждой хромосомы, составляющей бивалент.
Нити веретена деления сокращаются, вследствие чего биваленты распадаются на отдельные хромосомы, которые и притягиваются к полюсам клетки. В результате у каждого полюса формируется гаплоидный набор будущей клетки — n2c, за счет чего мейоз I и называется редукционным делением.

Происходит цитокинез — деление цитоплазмы. Формируются две клетки с гаплоидным набором хромосом. Очень короткая интерфаза после мейоза I сменяется новым делением — мейозом II.
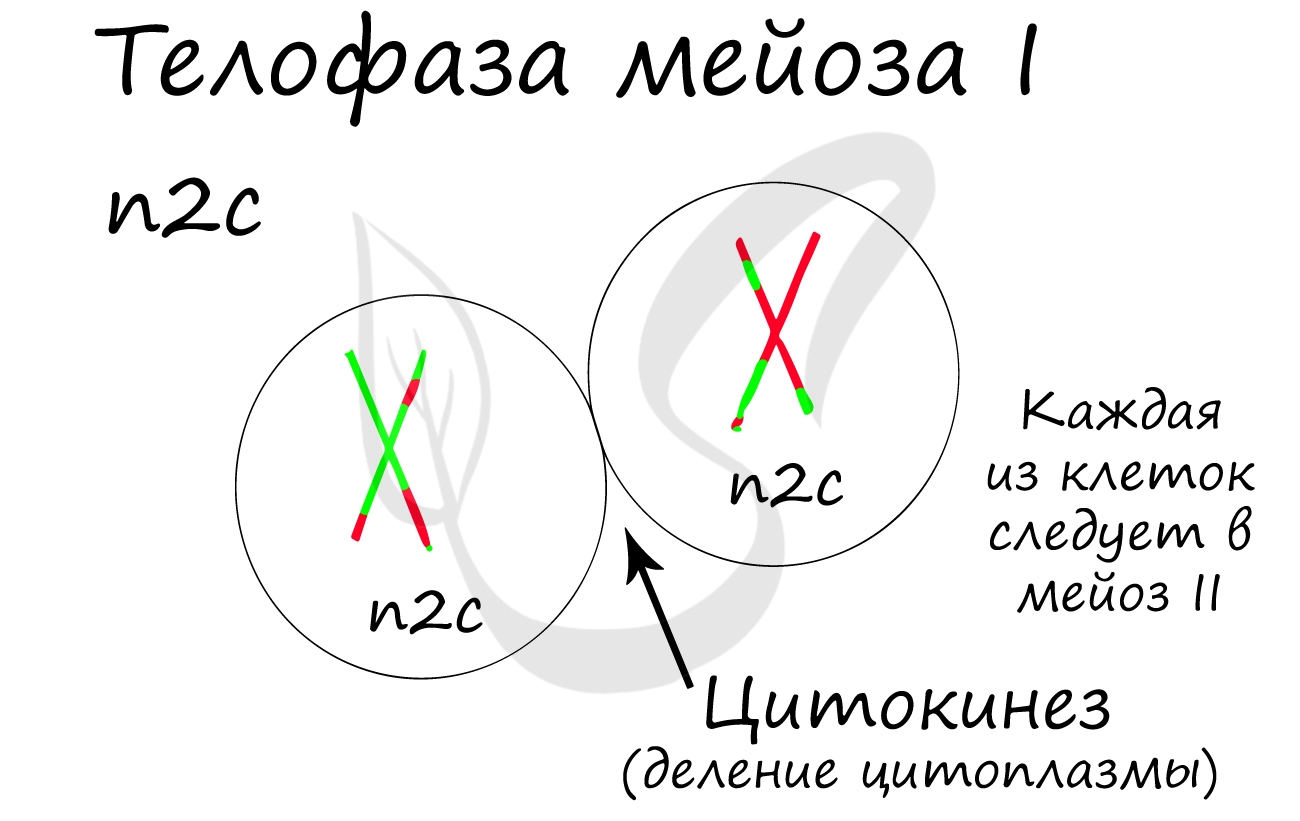
Мейоз II весьма напоминает митоз по всем фазам, поэтому если вы что-то подзабыли: поищите в теме про митоз. Главное отличие мейоза II от мейоза I в том, что в анафазе мейоза II к полюсам клетки расходятся не хромосомы, а хроматиды (дочерние хромосомы).
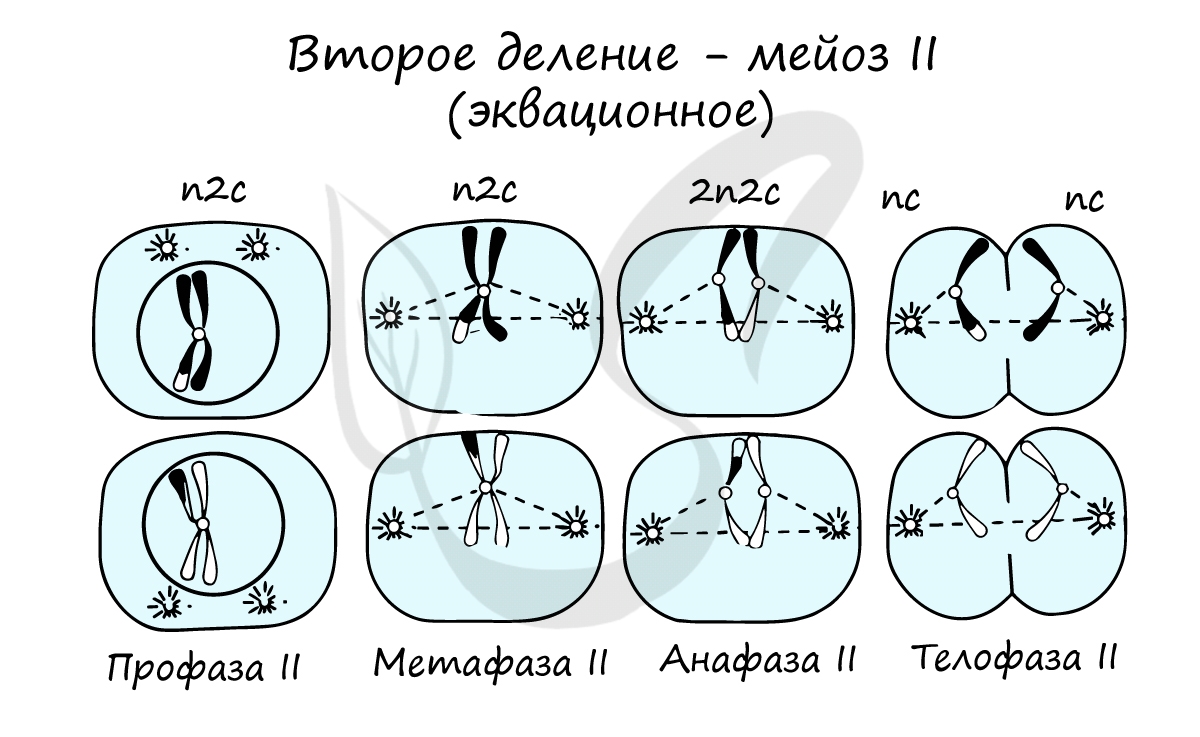
В результате мейоза I и мейоза II мы получили из диплоидной клетки 2n4c гаплоидную клетку — nc. В этом и состоит сущность мейоза — образование гаплоидных (половых) клеток. Вспомнить набор хромосом и ДНК в различных фазах мейоза нам еще предстоит, когда будем изучать гаметогенез, в результате которого образуются сперматозоиды и яйцеклетки — половые клетки (гаметы).
Сейчас мы возьмем клетку, в которой 4 хромосомы. Попытайтесь самостоятельно описать фазы и этапы, через которые она пройдет в ходе мейоза. Проговорите и осмыслите набор хромосом в каждой фазе.
Помните, что до мейоза происходит удвоение ДНК в синтетическом периоде. Из-за этого уже в начале мейоза вы видите их увеличенное число — 2n4c (4 хромосомы, 8 молекул ДНК). Я понимаю, что хочется написать 4n8c, однако это неправильная запись!) Ведь наша исходная клетка диплоидна (2n), а не тетраплоидна (4n) 😉
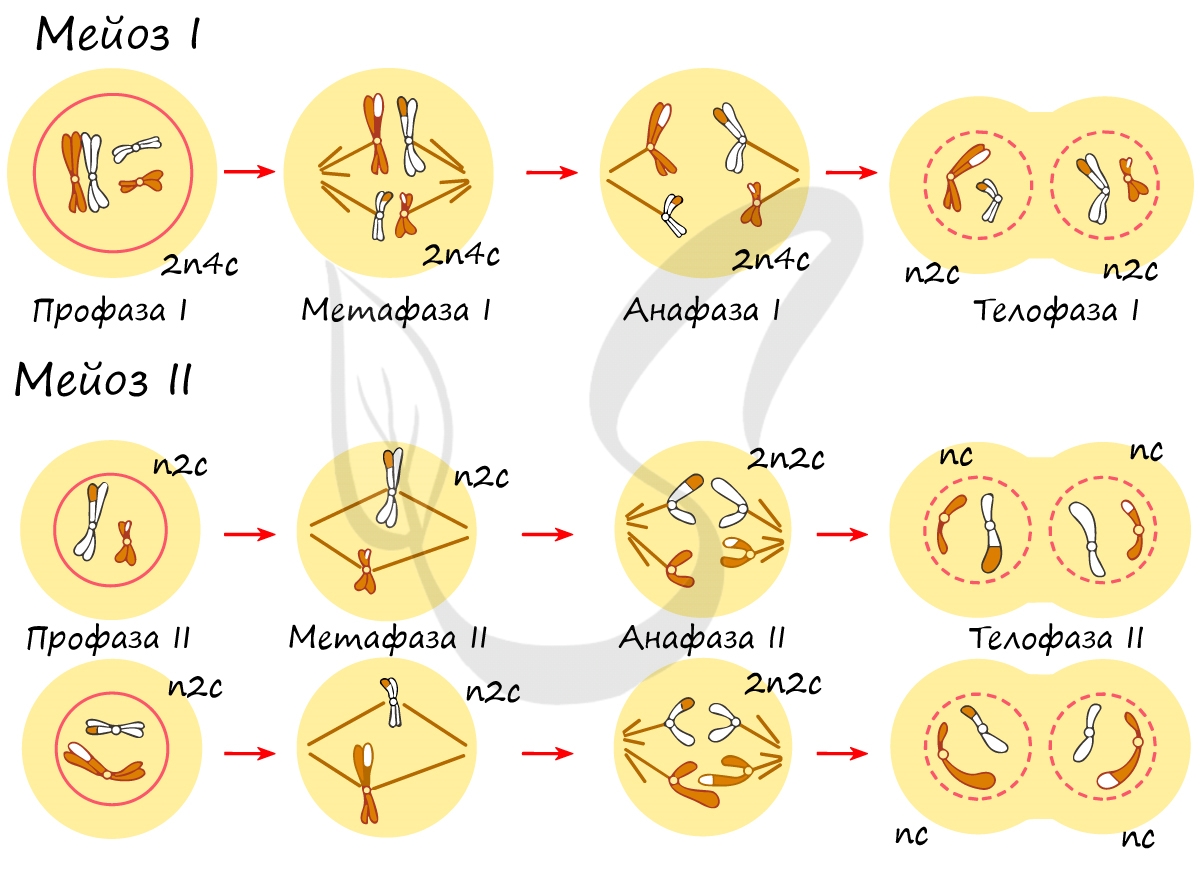
Итак, самое время обсудить биологическое значение мейоза:
- Поддерживает постоянное число хромосом во всех поколениях, предотвращает удвоение числа хромосом
- Благодаря кроссинговеру возникают новые комбинации генов, обеспечивается генетическое разнообразие состава гамет
- Потомство с новыми признаками — материал для эволюции, который проходит естественный отбор
Бинарное деление надвое
Митоз и мейоз возможен только у эукариот, а как же быть прокариотам — бактериям? Они изобрели несколько другой способ и делятся бинарным делением надвое. Оно встречается не только у бактерий, но и у ряда ядерных организмов: амебы, инфузории, эвглены зеленой.
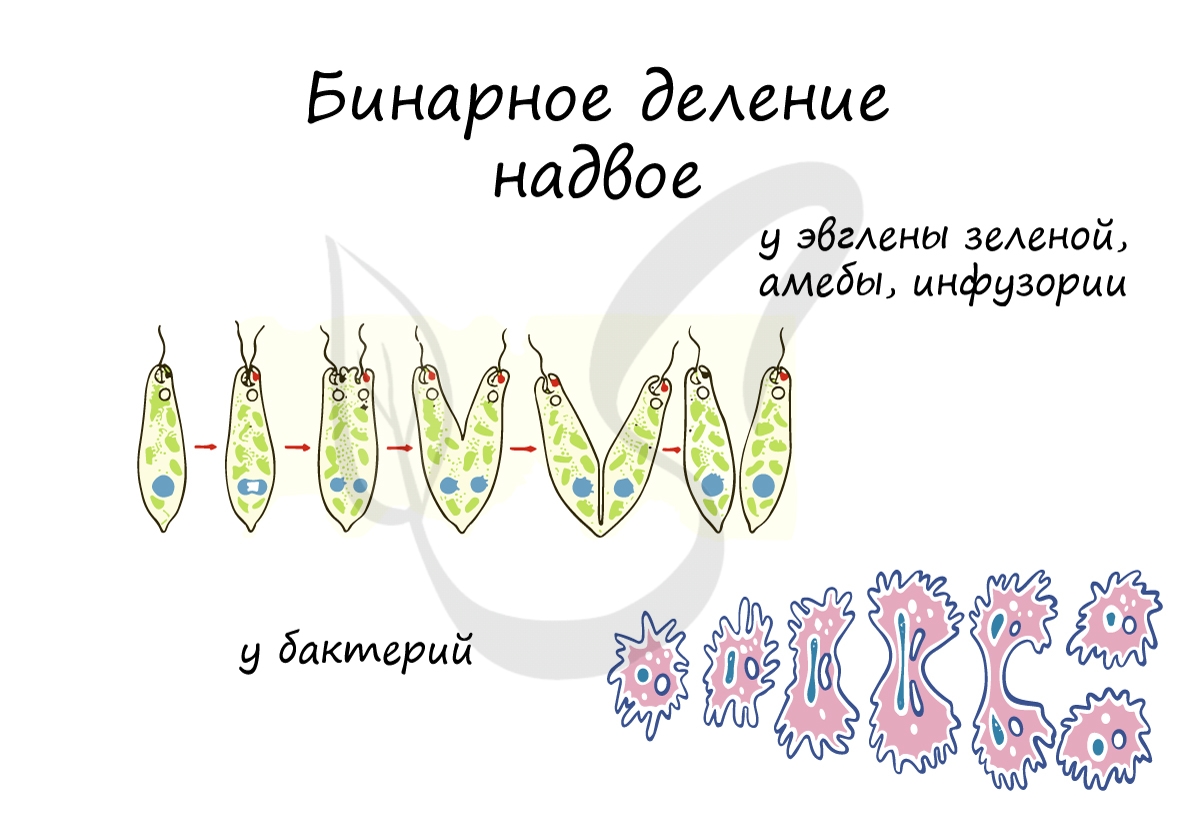
При благоприятных условиях бактерии делятся каждые 20 минут. В случае, если условия не столь благоприятны, то больше времени уходит на рост и развитие, накопление питательных веществ. Интервалы между делениями становятся длиннее.
Амитоз (от греч. ἀ — частица отрицания и μίτος — нить)
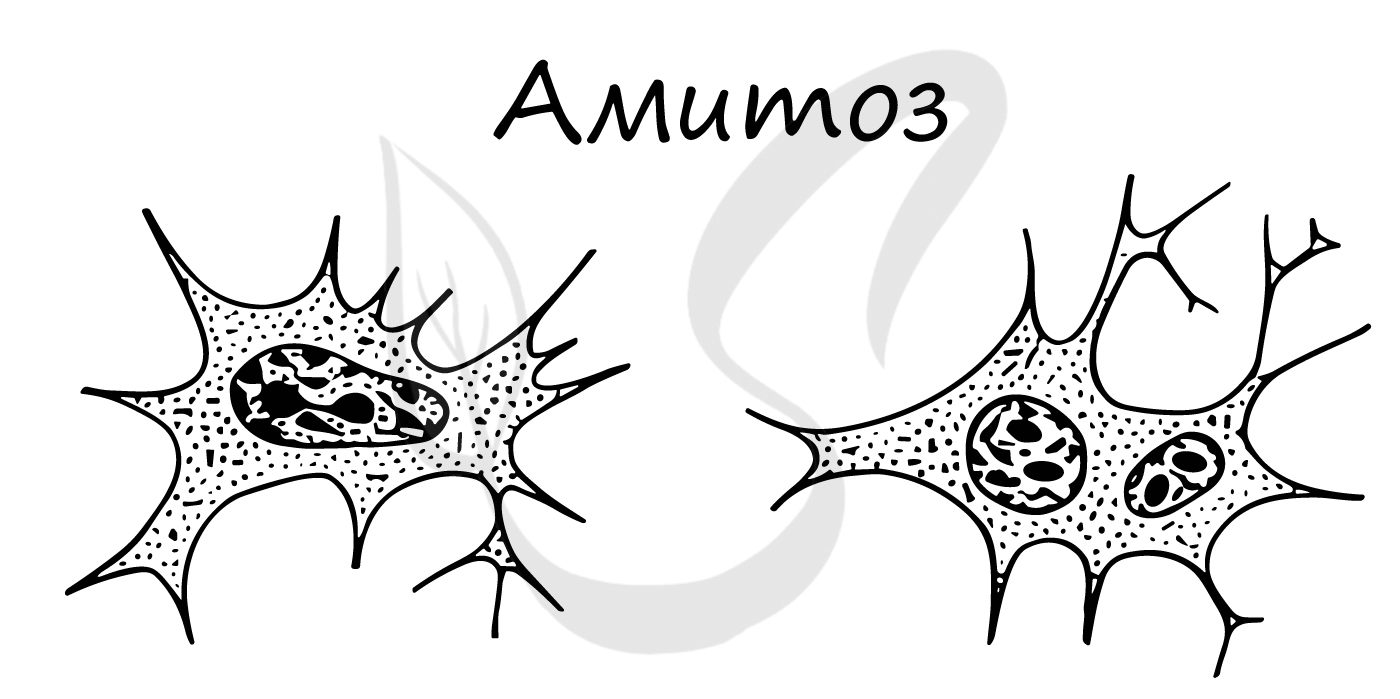
Амитоз встречается в раковых (опухолевых) клетках, воспалительно измененных, в старых клетках.
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Разные способы © 2022
Внимание! Информация, опубликованная на сайте, носит исключительно ознакомительный характер и не является рекомендацией к применению.
Читайте также: