
Архей это в биологии кратко
Обновлено: 05.07.2024
История
Археи было идентифицировано 1977 Карл Вёзе (Carl Woese) и Джорджем Фоксом (George Fox), основываясь на их отличия от других прокариот (филогенетические деревья, основанные на 16S рРНК). Когда 1977 биолог Карл Вёзе изучал генетику микробов, производили метан, он понял, что они отличаются от всех известных бактерий. Стенка клетки была уникальной — производила уникальные энзимы и ее генетический ряд не был похож ни на что исследуемое раньше. Очень быстро — в течение часа — стало понятно, что есть еще что-то, третья группа. Это был момент открытия. Карл Вёзе открыл третью форму жизни — группу одноклеточных, которую он назвал археями. Он открыл форму жизни, которая способна существовать где угодно, включая самые невероятные места. Эта группа была сначала назван археобактериямы (Archaebacteria) в отличие от эубактерий (Eubacteria), и они рассматривались как царства или подцарства. Вёзе убеждал, что археи являются представителями совсем другой ветви живых существ. Он позже переименовал группы в архей и бактерий, чтобы подчеркнуть это, и аргументировал тем, что вместе с эукариотами они охватывают три группы живых организмов.
Биологический термин архея (археи) не следует путать с архейская геологической эрой, также известной как Археозойська эра. Последний срок ссылается на древний период земной истории, когда археи и бактерии были единственными клеточными организмами, живших на планете. Возможные окаменелости этих микробов были датированы почти 3800000000 лет назад.
Археи, бактерии и эукариоты
Археи подобные других прокариот в большинстве аспектов структуры клетки и метаболизма. Однако, их генетическая транскрипция и трансляция — два центральных процессы в молекулярной биологии — не проявляют типичных для бактерий особенностей, но чрезвычайно подобные этих процессов у эукариот. Например, система трансляции архей использует эукариотические факторы инициации и элонгации, а их система транскрипции применяет TATA-связываемые белки и TFIIB, как у эукариот. Гены тРНК и рРНК археи содержат уникальные археальни интроны, которые не похожи ни к эукариотических, ни к бактериальным интронов.
Некоторые другие характеристики также выделяют архей. Подобно бактерий и эукариот, археи имеют основанные на глицерине фосфолипиды. Однако, три особенности липидов архей необычны:
- Липиды архей уникальны, так как стереохимия глицерина — обратная той, что найдена в бактериях и эукариот. Это убедительное свидетельство другого биосинтетического пути.
- Большинство бактерий и эукариот имеют мембраны, составленные преимущественно из эфир-липидов глицерина, тогда как археи имеют мембраны, состоящие из эфир-липидов глицерина. Даже когда бактерии имеют эфир-связанные липиды, стереохимия глицерина — бактериальная форма. Эти различия, возможно, является адаптацией части архей к гипертермофилии. Однако, стоит отметить, что даже мезофилични археи имеют эфир-связанные липиды.
- Липиды архей основанные на изопреноидного боковом цепочке. Это пятиуглеродный химическая группа, также обычная для каучука и является компонентом некоторых витаминов, распространенная в бактериях и эукариот. Однако, только археи применяют эти вещества в своих клеточных липидах, часто как боковые цепочки C-20 (четыре мономера) или C-40 (восемь мономеров). В некоторых архей изопреноидного боковой цепочку C-40 — фактически достаточно длинный, чтобы пройти сквозь мембрану, формируя монослой вместо клеточной мембраны с группами фосфата глицерина на обоих концах.
Эта адаптация распространенная в чрезвычайно теплолюбивых археи. Хотя они и не уникальны, клеточные стенки архей также необычны. Например, клеточные стенки большинства архей образованные внешними слоями белков или S-слоем. S-слоя распространенные в бактериях, они служат единственным компонентом клеточной стенки в некоторых организмах (например, в Planctomyces) или внешний слой во многих организмах с пептидогликана. За исключением одной группы метаногенов, археи не имеют пептидогликановый стенки. Однако даже в этом случае, пептидогликаны очень отличаются от разновидности, найденного в бактериях. Археи также имеют жгутики, которые значительно отличаются по составу от очень похожих жгутиков бактерий. Бактериальные жгутики — это измененная система секреции III типа, тогда как жгутики архей напоминают ворсинки IV типа (type IV pili), которые используют sec-зависимую систему секреции, несколько подобную, но все же отличную от системы секреции II типа.
Среда
Многие архей — экстремофилы. Некоторые живут при очень высоких температурах, часто выше 100 ° C, как те, которых нашли в гейзерах и черных курильщиках. Других найдено в очень холодных средах или в чрезвычайно соленой, кислой или щелочной воде. Однако, некоторые археи — мезофилы, живут в средах, подобных болота, сточных вод и почвы. Многие метаногенных архей найдено в пищеварительных трактах животных, например, жвачных животных, термитов и людей. Археи не патогенны, и неизвестно, чтобы какие-либо из них вызвали болезнь.
Архей конечно разделяют на три группы, в соответствии с среды. Это — галофиты, метаногены и термофилы. Галофилы живут в чрезвычайно соленых окружающих средах (например, многие из них живут в Мертвом море и на юге залива Сан-Франциско, придавая ей ярких цветов: от красного до зеленого). Метаногены живут в анаэробных окружающих средах и производят метан. Их можно найти в отложениях или в кишечниках животных. Термофилы живут в местах с высокими температурами, как, например, горячие источники. Эти группы не обязательно согласуются с молекулярным филогенезом, не полные но не взаимоисключающие. Тем не менее, они — полезный начальный пункт для подробного изучения.
Недавно несколько исследований показали, что археи существуют не только в мезофильных окружающих средах, но также иногда присутствуют в значительном количестве при низких температурах. Становится все более принятым, что метаногены обычно присутствуют в низкотемпературных средах, как, например, холодные осадки. Некоторые исследования даже свидетельствовали, что в этих температур путь метаногенеза может изменяться благодаря термодинамическим ограничениям, обусловленным низкими температурами. Возможно, даже, важнее то, что большое количество архей найдено в большинстве океанов, в холодных средах (Giovannoni и Stingl, 2005). Эти археи, которые относятся к группам, ранее исследованных, могут случаться в чрезвычайно высоких количествах (до 40% микробной биомассы), хотя они не были изолированы в чистой культуре. Сейчас мы не имеем почти никакой информации о физиологии этих организмов, что означает, что их влияние на глобальные биогеохимические циклы неизвестен. Одно недавнее исследование (Könneke и другие, 2006) показало, однако, что одна группа морских Crenarchaeota способна к фиксации азота, а это — черта, до сих пор неизвестна среди архей.
Генетика
Как правило, археи имеют одиночную кольцевую хромосому, размер которой может достигать 5751492 пар нуклеотидов в Methanosarcina acetivorans, что самый известный геном среди архей. Одну десятую размера этого генома составляет геном с 490885 парами нуклеотидов в Nanoarchaeum equitans, что маленький геном среди архей; он содержит только 537 генов, кодирующих белки. Также в архей обнаружены более мелкие независимые молекулы ДНК, так называемые плазмиды. Вероятно, плазмиды могут передаваться между клетками при физическом контакте, во время процесса похожего на конъюгации бактерий.
Археи могут быть поражены вирусами, содержащими двухцепочечной ДНК. Эти виды не родственные с другими группами вирусов и имеют различные необычные формы, так как бутылки, крючки, капли и т. Эти вирусы были тщательно изучены на термофилы, в основном рядов Sulfolobales и Thermoproteales. В 2009 году был открыт вирус, содержащий одноцепочечную ДНК и поражает галофильные археи. Защитные реакции архей против вирусов могут включать механизмы близок к РНК-интерференции эукариот.
Археи генетически отличны от эукариот и бактерий, причем до 15% белков, кодируемых одним геном бактерии, являются уникальными для этого домена, хотя функции многих этих белков неизвестны. Большая часть уникальных белков, функция которых известна, принадлежит евриархеотам и задействована в метаногенеза. Белки, общие для архей, бактерий и эукариот, участвующих в основных клеточных функциях и касаются в основном транскрипции, трансляции и метаболизма нуклеотидов. К другим различий архей можно отнести организацию генов, выполняющих связанные функции (например, гены, отвечающие за различные этапы одного и того же метаболического процесса), в оперон и большие различия в строении тРНК и их аминоацил-тРНК синтетаз.
Транскрипция и трансляция архей больше напоминают эти процессы в клетках эукариот, чем бактерий, причем РНК-полимераза и рибосомы архей очень близки к аналогичным структур у эукариот. Хотя у архей есть только один тип РНК-полимеразы, по строению и функции она близка к РНК-полимеразы II эукариот, при этом подобные группы белков (главные факторы транскрипции) обеспечивают связывание РНК-полимеразы с промотором гена. в то же время другие факторы транскрипции архей более близки к таковым у бактерий. Процессинг РНК в архей проще, чем у эукариот, поскольку большинство генов не содержит интронов, хотя в генах их тРНК и рРНК их достаточно много, также они присутствуют в небольшом количестве генов, кодирующих белки.
Размножение
Археи размножаются бесполым путем: бинарным или множественным делением, фрагментацией или почкованием. Мейоза не происходит, поэтому даже у представителей конкретного вида архей существуют в более чем одной форме, все они имеют одинаковый генетический материал. Клеточное деление определяется клеточным циклом: после того, как хромосома реплицировать и две дочерние хромосомы разошлись, клетка делится. Детали были изучены только для рода Sulfolobus, но особенности его цикла очень схожи с особенностями у эукариот и у бактерий. Репликация хромосом начинается с многих точек начала репликации с помощью ДНК-полимеразы, похожей на аналогичные ферменты эукариот. Однако белки, управляющие клеточным делением, такие как FtsZ, которые формируют сжимающее кольцо вокруг клетки и компоненты септы, проходящей через центр клетки, схожие с их бактериальными эквивалентами.
Археи не образуют споры. Некоторые виды Haloarchaea могут претерпевать изменения фенотипа и существовать как клетки нескольких различных типов, включая толстостенными клетками, устойчивыми к осмотического шока, позволяющие археям выживать в воде с низкой концентрацией соли. Однако эти структуры не служат для размножения, а скорее помогают археям осваивать новые среды обитания.
Эволюция и классификация
Название говорит о том, что эти организмы в наше время многими исследователями считаются древнейшими живыми организмами на Земле.
Археи разделены на две основные группы, основанные на деревьях 16S рРНК: Euryarchaeota и Crenarchaeota. Две другие группы были также созданы для определенных экологических образцов и особого вида Nanoarchaeum equitans, найденных в 2002 Карлом Стеттером (Karl Stetter), но их сходство с других групп все еще исследовано очень плохо.
Карл Вёзе (Carl Woese) убедил, что бактерии, археи и эукариоты представляют собой уникальные наследственные линии, которые довольно рано отклонились от наследственного Прогенота (Progenote) с плохо развитым генетическим аппаратом. Эта гипотеза отражена в имени археи (Archaea, от Греческого archae или древний). Позже он формально рассматривал эти группы как империи, каждая из которых охватывает несколько царств. Этот раздел стал самым популярным, хотя идея Прогенота не всеми поддерживается. Некоторые биологи, однако, утверждают, что, археобактерии и эукариоты произошли от специализированных бактерий.
Взаимоотношения между археями и эукариотами остаются важной проблемой. Учитывая сходство, указанную выше, много генетических деревьев располагают эти две группы вместе. Некоторые ставят эукариот ближе к Eurarchaeota, чем к Crenarchaeota, хотя мембранная химия предлагает наоборот. Однако открытие архея-подобных генов в определенных бактериях, например, Thermotoga, делает их взаимоотношения трудным для определения. Некоторые исследователи считают, что эукариоты возникли из смешения археи и бактерии, они стали ядром и цитоплазмой, объясняют различную генетическую схожесть, но эта теория не может объяснить происхождение клеточных структур.
Внутренняя классификация
Архебактерии разнообразные по типу обмена веществ, физиологическими и экологическими особенностями: среди них встречаются аэробных и анаэробных, хемогетеротрофы и хемоавтотрофы, нейтрофилы и ацидофилы. Некоторым Архебактерии (галобактерии) присущ особый тип фотосинтеза, когда свет поглощается НЕ хлорофиллом, а бактериородопсинном. Другим археям властвуй энергетический процесс, в результате которого образуется метан. Описано более 40 видов археи (25 родов), относящихся к 5 различных групп: Метанобразующие бактерии, сиркоокислюючи термоацидофилы, сирковидновлюючи термофилы, галобактерии, термоплазма.
Расшифровка одиночных генов привела к расшифровке целых геномов, сейчас 24 архейни геномы завершения и 22 частично завершено (1).

История планеты Земля состоит из четырех эонов – отрезков времени, которые охватывают период от 4 млрд лет назад до современности. Самым первым из них является архей продолжительностью в 1,5 млрд лет. Термин для его обозначения был предложен американским геологом Джеймсом Дана в 1872 году.
Периодизация Архея
Архейской эре предшествовал катархей, но его осадочные породы неизвестны. Его нижней временной границей является время образования планеты Земля – 4,6 млрд лет назад, а закончился он 4 млрд лет назад.

Рис. 1. Архейская эра.
Периодов архейской эры геологи выделяют четыре:
- Эоархей. 4-3,6 млрд лет назад. Формировалась земная кора, вулканические кратеры, соленые водоемы с горячей водой. Появились организмы – цианобактерии.
- Палеоархей.
На период эоархей приходится так называемая “Поздняя тяжелая бомбардировка” – время формирования кратеров на Луне и Марсе.

Рис. 2. Архейская эра фауна.
Растения и организмы
Флора и фауна архея не особо разнообразны. В данный период на планете существовал только один вид растений – одноклеточные нитчатые водоросли (сфероморфид).
В дальнейшем водоросли привели к формированию лишайников. В архее появились одноклеточные безъядерные организмы, которые называются прокариоты. Они могли производить кислород и создавать условия для возникновения новых организмов. Существует спорная теория о зарождении в этот период эукариотов – организмов из царства грибов.
Цианобактерии архея найдены геологами в Канаде, Африке и в азиатской части России.
Они повлияли на возникновение кристаллов арагонита и на появление кремневых и карбонатных залежей.
Другие особенности архейского периода
Температура воды в архее составляла 90 градусов. Единственной сушей в мезоархее являлись вулканические острова, которые можно считать первыми горами. Других типов гор не могло появиться из-за отсутствия тектонической активности. В результате столкновения с крупным астероидом, 3 млрд лет назад на планете появился самый древний из кратеров. Он обнаружен в Гренландии. Первые снег и лед земного происхождения появились 2,9 млрд лет назад. Первое оледенение получило название – понгольское (в честь города Понгола в ЮАР).
К основным событиям завершающего этапа этого эона (неоархею) следует отнести превращение материка Ур в суперконтинент Кенорленд. Это произошло около 2,7 млрд лет назад. Позже, около 2,6 млрд лет назад, Кенорленд присоединился к суперконтиненту Арктида.
Следующей геологической эрой стал протерозой. Он продолжался 2 млрд лет и включал в себя: палеопротерозой, мезопротерозой и неопротерозой.

Рис. 3. Архей.
Что мы узнали?
АРХЕ́И (от греч. ἀ ρχαῖος – изначальный, древний), архебактерии (Archaea), группа прокариот, выделяемая в отдельную систематич. категорию наравне с истинными бактериями (эубактериями) и ядерными организмами – эукариотами. Деление прокариот на А. и истинные бактерии было предложено амер. биологом К. Везе (1980) на основании значит. различий в структуре генома, свидетельствующих о раннем расхождении этих групп организмов в ходе эволюции. А. объединяют в домен, который включает три царства: Euryarchaeota (метаногены, сульфатредуцирующие А., галофилы и некоторые гипертермофилы), Crenarchaeota (только гипертермофилы) и Nanoarchaeota (развиваются только в клетках других А. и неспособны к самостоят. существованию). Морфологически А. сходны с бактериями: их клетки размером в ср. 1–2 мкм представляют собой палочки или кокки, в некоторых случаях спириллы, иногда собраны в агрегаты, размножаются делением клетки на две дочерние, подвижные формы снабжены одним или несколькими жгутиками. В то же время спорообразование у А. неизвестно. Их клетки окружены одним или несколькими слоями белковых или гликопротеиновых молекул (т. н. S-слои), либо оболочкой, сходной с клеточной стенкой грамположительных бактерий, но с др. составом биополимеров, обеспечивающих её прочность. У некоторых А. она вообще отсутствует, а липиды их цитоплазматич. мембраны представлены не эфирами глицерина и жирных кислот, как у бактерий, а ди- или тетраизопреноидными эфирами. Геномы А. организованы так же, как у бактерий, как правило, в виде одной замкнутой в кольцо молекулы ДНК (хромосомы). Особенности А. проявляются также в нуклеотидном составе транспортной РНК, строении рибосом, структуре фермента ДНК-зависимой РНК-полимеразы, которая по своим характеристикам ближе к РНК-полимеразе эукариот. Отличия в строении клеточной стенки и аппарата синтеза белка обусловливают устойчивость А. к действию бактериальных антибиотиков. Более того, А. свойственны типы метаболизма и др. биохимич. особенности, не обнаруживаемые у представителей бактерий и эукариот.
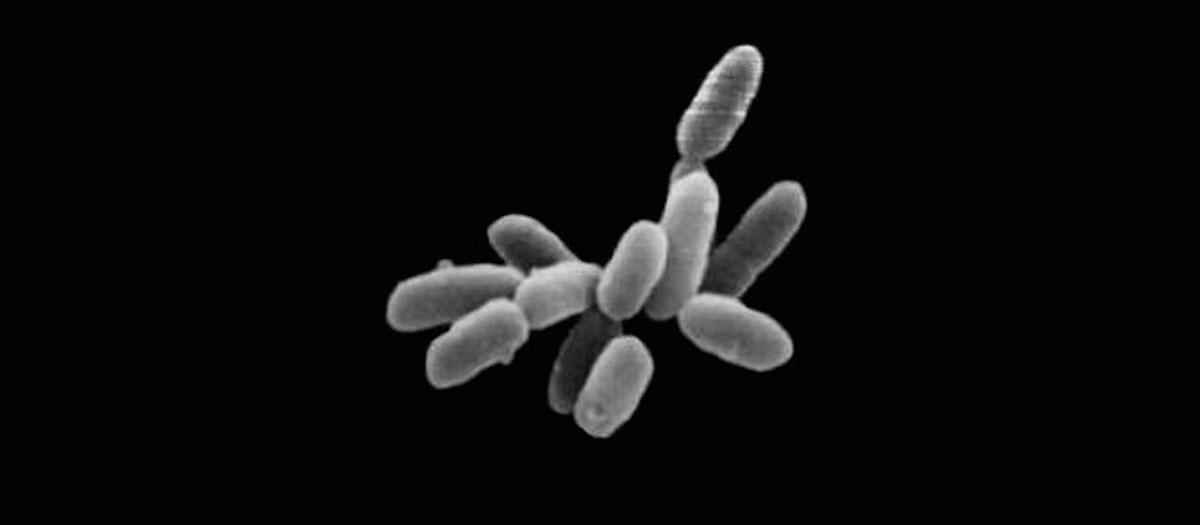
Археи – домен микроорганизмов, которые были обнаружены в начале 1970-х годов. Подобно бактериям, они являются одноклеточными прокариотами. Первоначально археи считались бактериями, пока анализ ДНК не показал, что это разные организмы. Они настолько различны, что открытие побудило ученых придумать новую систему классификации жизни.
Мы знаем, что многие из них являются экстремофилами, живущими и процветающими в самых экстремальных условиях, таких как чрезвычайно горячие, засушливые, кислотные или щелочные среды обитания.
Клетки Архей
Археи являются чрезвычайно маленькими микроорганизмами, которые нужно рассматривать под электронным микроскопом, чтобы идентифицировать их характеристики. Как и бактерии, они имеют различные формы, включая кокки (круглые), бациллы (стержнеобразные) и произвольные формы. Археи имеют типичную анатомию прокариотических клеток: плазмидная ДНК, клеточная стенка, клеточная мембрана, цитоплазма и рибосомы. У некоторых архей также есть длинные выросты, называемые жгутиками, которые помогают в движении.
Домен Археи
Живые организмы подразделяются на три домена и шесть царств. Домены включают:
- археи (Archaea);
- бактерии (Bacteria);
- эукариоты (Eukaryota).
Согласно последним данным, археи подразделяется на следующие основные типы:
Crenarchaeota
Crenarchaeota состоит в основном из гипертермофилов и термоацидофилов. Гипертермофильные микроорганизмы живут в чрезвычайно жарких или холодных условиях. Термоацидофилы представляют собой архей, которые обитают в чрезвычайно жарких и кислых средах. Их местообитания имеют рН от 5 до 1. Вы найдете эти организмы в гидротермальных жерлах и горячих источниках.
- Sulfolobus acidocaldarius – обнаружен вблизи горячих и кислотных вулканических источников, содержащих серу.
- Pyrolobus fumarii – живут при температуре от 90 до 113 градусов Цельсия.
Euryarchaeota
Euryarchaeota – тип архей, включающий галофилов и метаногенов. Экстремальные галофильные микроорганизмы живут в соленой среде. Эти организмы обитают в соленых озерах или в районах, где испарилась морская вода.
Метаногены нуждаются в свободных от кислорода (анаэробных) условиях, чтобы выжить. Они производят метан как побочный продукт метаболизма. Метаногены распространены в таких местах, как болота, водно-болотные угодья, ледяные озера, сточные воды, кишечники животных и людей.
- Halobacterium – насчитывает несколько видов галофильных организмов, которые встречаются в соляных озерах и в условиях сильного засоления.
- Methanococcus – Methanococcus jannaschii считается первым генетически упорядоченным археем. Этот метаноген живет вблизи гидротермальных отверстий.
- Methanococcoides burtonii – эти психофильные (холодолюбивые) метаногены обитают в Антарктиде и способны выдерживать чрезвычайно холодные температуры.
Nanoarchaeota
Nanoarchaeota – тип архей, который был включен в основную классификацию домена с открытием Nanoarchaeum equitans в 2002 году, обнаруженного в гидротермальных источниках у берегов Исландии.
Korarchaeota
Представители типа Korarchaeota считаются очень примитивными живыми микроорганизмами. На сегодняшний день мало известно об основных характеристиках этих архей. Мы знаем, что они термофильные и встречаются в горячих источниках и обсидиановых бассейнах.
Thaumarchaeota
Thaumarchaeota – предложенный в 2008 году тип архей, который на данный момент включает следующие виды: Nitrosopumilus maritimus, Cenarchaeum symbiosum, Nitrososphaera viennensis и Nitrososphaera gargensis. Все представитель этого типа архей принадлежат к хемолитотрофам, окисляющим аммиак.
Филогенез Архей
Археи – интересные с точки зрения науки организмы, поскольку у них есть гены, похожие на бактерий и эукариот. Филогенетически, археи и бактерии, как предполагают ученые, развились от одного общего предка. Считается, что эукариоты отделились от архей миллионы лет спустя. Это указывает на то, что археи более тесно связаны с эукариотами, чем с бактериями.
Читайте также: