
Закон кетле в генетике кратко
Обновлено: 04.07.2024
При рассмотрении основных законов генетики необходимо отметить, что они носят статистический характер, т.е. эти законы можно обнаружить при изучении очень большого количества объектов. Так, изучив 10 особей данного вида, обнаружить тот или иной закон нельзя — слишком мало параллельных наблюдений. Чем больше параллельных наблюдений будет сделано, тем четче и рельефнее будет проявляться тот или иной генетический закон.
Обзор законов генетики, открытых Г. Менделем
Используя гибридологический метод исследования, Г. Мендель открыл законы независимого наследования признаков. Эти законы были открыты при изучении закономерностей наследования у растений гороха, при этом применялось моногибридное и дигибридное скрещивание.
1. Первый закон Менделя — закон единообразия всех особей первого поколения для любого вида скрещивания (как моно-, так и полигибридного скрещивания): при любом скрещивании все особи первого поколения (F1) характеризуются одинаковым фенотипом по скрещиваемому признаку.
Этот фенотип определяется либо доминантным признаком, либо возникают промежуточные признаки, либо появляются новые признаки, как результат взаимодействия генов. Так, при скрещивании гороха с желтыми и зелеными семенами в первом поколении все растения имеют желтые семена (доминантно-рецессивный характер наследования). При скрещивании фиалки Ночная красавица с белыми и красным цветами все растения первого поколения имеют розовые цветы (промежуточный характер наследования).
Для скрещивания берут гомозиготные организмы. Например, материнский организм имеет гены желтого цвета семени (обозначим АА), а отцовский — гены зеленого цвета семени (обозначим аа). Тогда гаметы матери (яйцеклетки) содержат по одному гену желтого цвета семени (А) и гаметы отца (спермии) — по одному гену зеленого цвета семени (а). При оплодотворении образуется гетерозигота, содержащая гены рассмотренных выше альтернативных признаков; обозначается Аа. Так как в данном случае наблюдается доминантно-рецессивный характер наследования признаков, все особи первого поколения (F1) имеют семена желтого цвета, т. е. характеризуются одинаковым фенотипом по данному признаку.
В случае равноценного характера взаимодействия генов наблюдается промежуточный характер наследования. В этом случае в тоже возникают гетерозиготные организмы (обозначенные А1А2) с одинаковым фенотипом по конкретному признаку. Так, при скрещивании фиалки Ночная красавица с белыми и красными цветами в F1 все растения имеют розовые цветы.
2. Второй закон Менделя — закон расщепления признаков (закон моногибридного скрещивания) — иногда его называют правилом расщепления признаков. Этот закон справедлив для моногибридного скрещивания и проявляется при скрещивании разных особей, полученных при моногибридном скрещивании во втором поколении (F2): при скрещивании особей первого поколения, полученного после моногибридного скрещивания, у потомства наблюдается расщепление признаков в определенном количественном отношении, которое для доминантно-рецессивного наследования составляет 3:1, а для промежуточного наследования 1:2:1 (цифра 2 означает, что гибриды относятся к особям с промежуточным признаком).
1. Скрещивая растения гороха с гладкими семенами (F1 полученное после скрещивания растений с гладкими и морщинистыми семенами), получаем второе поколение (F2), при этом 3/4 потомства имеют гладкие семена, а 1/4 — морщинистые.
Это явление можно объяснить так. Растения первого поколения гетерозиготны (обозначим их Аа). Они дают два вида гамет (обозначим их А и а). Эти гаметы характерны и для отцовского, и для материнского организмов. При реализации процессов оплодотворения возможны четыре сочетания (в них на первом месте стоит ген, полученный от организма матери, его выделим): АА, Аа, аА и аа. Сочетание АА соответствует гомозиготе по доминантному признаку (гладкие семена); сочетания Аа и аА соответствуют гетерозиготе (гладкие семена), а последнее сочетание аа гомозиготно по рецессивному признаку. Таким образом, во втором поколении возникают три разных генотипа по данному признаку и им соответствует только два фенотипа.
2. Скрещивая растения фиалки с розовыми цветками (F1), получаем F2, в котором 1/4 часть потомства имеет белые цветки, 1/4 часть — красные, а половина потомства (2/4) — розовые. Объяснение этого явления такое же, как и для примера 1, но здесь наблюдаем разницу — трем генотипам по данному признаку (А1A1, А1А2 и А2А1; А2А2) соответствуют три фенотипа (белый, розовый и красный цветки).
3. Третий закон Менделя — закон полигибридного скрещивания или закон независимого расщепления признаков.
Этот закон проявляется во втором поколении при дигибридном и полигибридном (три-, тетра- и др.) скрещивании: при скрещивании особей первого поколения, полученного при скрещивании по дигибридному (полигибридному) типу, в потомстве (во втором поколении) происходит расщепление признаков (для доминантно-рецессивного характера наследования) в количественном отношении, выражаемом формулой (3 + 1) n , где n = 2, 3, 4 и т. д.
Для цитологического объяснения удобно применять решетку Пеннета. Проанализируем сведения, приведенные на рисунке. Сначала скрещивали растения вида горох обыкновенный с желтыми и гладкими семенами (ген желтого цвета семени обозначим А, а гладкой формы — В) с растениями, у которых были зеленые морщинистые семена (ген зеленого цвета семени обозначим а, ген морщинистой формы — b). Все полученные растения первого поколения гетерозиготны и имеют желтые гладкие семена (доминантно-рецессивное наследование, при котором гены желтого цвета и гладкой поверхности семян доминируют над генами зеленого цвета и морщинистой формы). Назовите закон, проявившийся в данном случае.
После скрещивания растений первого поколения получили F2 — второе поколение, у которых наблюдаем закон независимого расщепления признаков: 1/16 часть всех растений имеет зеленые морщинистые семена, 3/16 — зеленые и гладкие; 3/16 — желтые и морщинистые, а остальные (9/16) — желтые и гладкие. Следовательно, при дигибридном скрещивании наблюдается появление в F2 четырех фенотипов (по данным признакам).
При дигибридном скрещивании каждое растение образует четыре вида гамет, а для двух родителей эти гаметы могут дать 16 сочетаний. В результате получается, что 1/16 часть поколения является гомозиготной по рецессивному и столько же — по доминантному признакам, а все остальные особи гетерозиготны хотя бы по одному признаку; абсолютно гетерозиготных (по двум признакам) только 4/16 части поколения.
Подсчет показывает, что четырем фенотипам при дигибридном скрещивании соответствует девять фенотипов (сделайте этот подсчет самостоятельно).
Необходимо отметить, что третий закон Менделя справедлив, если гены, ведающие данными признаками, находятся в разных парах хромосом; так, ген окраски семени располагается в одной паре гомологических хромосом, а ген, определяющий форму семян, — в другой.
Вероятно, существуют случаи, когда гены, ведающие теми или иными признаками, содержатся в одной паре хромосом. Для таких вариаций законы Менделя (кроме первого) не применимы. Эти случаи подчиняются закону Моргана.
Закон Моргана — закон сцепленного наследования признаков
Ряд организмов имеет небольшое число хромосом, поэтому многие гены, определяющие различные группы альтернативных признаков, находятся в одной гомологичной паре хромосом, т.е. являются сцепленными и передаются потомству вместе. Так, у плодовой мушки дрозофилы ген, определяющий длину крыльев, и ген, ответственный за цвет тела, находятся в гомологичных хромосомах.
Дигибридное скрещивание, проведенное по данным признакам во втором поколении, не даст независимого расщепления признаков, т. е. не будет соответствовать третьему закону Менделя. Это явление обнаружил Т. Морган и сформулировал его в форме закона сцепленного наследования:
При дигибридном скрещивании организмов, у которых гены находятся в одной паре гомологичных хромосом, во втором поколении наблюдается расщепление признаков не по третьему, а по второму закону Менделя.
Скрещивая мушек дрозофил с темным цветом тела и нормальными крыльями (доминирующие признаки) с мушками, имеющими укороченные крылья и серое тело (рецессивные признаки), было получено гетерозиготное поколение (F1) с темными телами и нормальными крыльями.
При скрещивании особей первого поколения получили организмы, у которых 1/4 часть поколения имела укороченные крылья и серое тело, а 1/3 часть поколения — нормальные крылья и темное тело. Это объясняется тем, что гены окраски тела и длины крыльев располагаются в одной паре гомологических хромосом, т. е. являются сцепленными. Однако среди особей F2 наблюдали и насекомых, имеющих темное тело и укороченные крылья, и особей с серым телом и нормальными крыльями. Это объясняется кроссинговером, при котором хромосомы в результате конъюгации и перекрещивания обмениваются участками гомологических хромосом. Но эти явления носят случайный характер и не подчиняются математическим закономерностям.
Закон гомологических рядов наследственной изменчивости
В процессе изучения закономерностей наследования мутационной (наследственной) изменчивости Н. И. Вавилов открыл закон, известный в науке под названием закона гомологических рядов наследственной изменчивости, который был сформулирован следующим образом:
Если виды и роды генотипически связаны друг с другом, единством происхождения, то они образуют ряды форм организмов, сходных по своим признакам, т. е. гомологические ряды.
остистые формы → малоостистые формы → безостые формы.
Эти формы характерны и для пшеницы, и для ржи, и для ячменя.
Гомологические ряды известны не только для злаков, но и для других растений.
Анализирующее скрещивание
Как было показано выше, для выявления закономерностей наследования признаков необходимо первичному скрещиванию подвергать гомозиготные особи. Однако фенотип по данному признаку не всегда является признаком гомозиготности данного организма, например горох с желтыми семенами может быть как гомозиготным (АА) по доминантному признаку, так и гетерозиготным (Аа). Поэтому необходим метод выявления гомозиготности, которым является анализирующее скрещивание.
Для анализирующего скрещивания используют организмы, обладающие рецессивным альтернативным признаком, и эти организмы скрещивают с организмами, гомозиготность которых необходимо установить. Если в первом поколении не происходит расщепления признаков, то данные организмы являются гомозиготными по доминантному признаку, в противном случае (в этом поколении появятся организмы, обладающие рецессивными признаками) — исследуемые организмы гетерозиготны.
Рассмотрим пример. При скрещивании морских свинок с короткой шерстью (рецессивный признак) (аа — обозначение родительского организма, который дает гаметы а) со свинками с длинной шерстью (доминантный признак) в первом поколении получили потомство с длинной шерстью. Вывод — длинношерстные свинки являются гомозиготными (АА — обозначение родительского организма, который дает гаметы А), так как зигота первого поколения будет соответствовать обозначению Аа. Случай, когда длинношерстные свинки были гетерозиготными, охарактеризуйте самостоятельно. Ответьте также на вопрос: можно ли для анализирующего скрещивания использовать гомозиготные организмы, обладающие доминантными признаками и почему? Докажите свой ответ, используя цитологические представления.
Взаимодействие генов
При изучении закономерностей наследования признаков необходимо учитывать характер воздействия одних генов на другие. В предыдущих подразделах было показано, что аллельные гены оказывают определенное воздействие друг на друга, при котором наблюдается или доминантно-рецессивный характер взаимодействия, или при воздействии аллельных генов друг на друга возникает новый признак, промежуточный между исходными признаками — при одинаковом воздействии генов друг на друга.
В генетических исследованиях было обнаружено, что взаимодействовать между собой могут и неаллельные гены, и при их взаимодействии у организма появляются новые признаки, т.е. возникает новый фенотип. Так, при скрещивании кур с розовидным и ореховидным гребнями получили первое потомство кур с гороховидными гребнями. Скрещивание особей друг с другом привело к расщеплению признаков не по второму закону Менделя (как это предполагалось, ведь внешне скрещивание было моногибридным), а по третьему закону — закону независимого расщепления признаков.
Было обнаружено, что 1/16 часть потомства имела простой гребень, 3/16 — розовидный, 3/16 — ореховидный, а остальные (9/16) — гороховидный. Следовательно, розовидная и ореховидная формы гребня определяются не одним геном, а являются результатом взаимодействия двух неаллельных генов, так как характер расщепления признаков соответствует дигибридному скрещиванию.
Множественное действие генов
Генетиками было установлено, что один ген может влиять либо на отдельный конкретный признак, либо оказывать влияние на несколько признаков, т.е. иметь множественное действие. Так, у водосбора имеется ген окраски цветка, при этом ген красной окраски оказывает влияние на окраску листьев (у водосбора с красными цветами листья фиолетовые). Кроме того, этот ген оказывает влияние на длину стебля и массу семян — стебель у водосбора с красными цветами более длинный, а семена имеют большую массу, чем семена у водосбора с другой окраской цветка.
Мушка дрозофила имеет ген, определяющий цвет глаз. Если у дрозофилы содержится ген, вызывающий отсутствие пигмента в глазе, то эти мушки имеют малую плодовитость, более короткую продолжительность жизни и специфическую окраску внутренних органов.
Наследственность — свойство организмов передавать свои признаки от одного поколения к другому.
Изменчивость — свойство организмов приобретать новые по сравнению с родителями признаки. В широком смысле под изменчивостью понимают различия между особями одного вида.
Признак — любая особенность строения, любое свойство организма. Развитие признака зависит как от присутствия других генов, так и от условий среды, формирование признаков происходит в ходе индивидуального развития особей. Поэтому каждая отдельно взятая особь обладает набором признаков, характерных только для нее.
Фенотип — совокупность всех внешних и внутренних признаков организма.
Ген — функционально неделимая единица генетического материала, участок молекулы ДНК, кодирующий первичную структуру полипептида, молекулы транспортной или рибосомной РНК. В широком смысле ген — участок ДНК, определяющий возможность развития отдельного элементарного признака.
Генотип — совокупность генов организма.
Локус — местоположение гена в хромосоме.
Аллельные гены — гены, расположенные в идентичных локусах гомологичных хромосом.
Гомозигота — организм, имеющий аллельные гены одной молекулярной формы.
Гетерозигота — организм, имеющий аллельные гены разной молекулярной формы; в этом случае один из генов является доминантным, другой — рецессивным.
Рецессивный ген — аллель, определяющий развитие признака только в гомозиготном состоянии; такой признак будет называться рецессивным.
Доминантный ген — аллель, определяющий развитие признака не только в гомозиготном, но и в гетерозиготном состоянии; такой признак будет называться доминантным.
Методы генетики
Основным является гибридологический метод — система скрещиваний, позволяющая проследить закономерности наследования признаков в ряду поколений. Впервые разработан и использован Г. Менделем. Отличительные особенности метода: 1) целенаправленный подбор родителей, различающихся по одной, двум, трем и т. д. парам контрастных (альтернативных) стабильных признаков; 2) строгий количественный учет наследования признаков у гибридов; 3) индивидуальная оценка потомства от каждого родителя в ряду поколений.
Скрещивание, при котором анализируется наследование одной пары альтернативных признаков, называется моногибридным, двух пар — дигибридным, нескольких пар — полигибридным. Под альтернативными признаками понимаются различные значения какого-либо признака, например, признак — цвет горошин, альтернативные признаки — желтый цвет, зеленый цвет горошин.
Кроме гибридологического метода, в генетике используют: генеалогический — составление и анализ родословных; цитогенетический — изучение хромосом; близнецовый — изучение близнецов; популяционно-статистический метод — изучение генетической структуры популяций.
Генетическая символика
Предложена Г. Менделем, используется для записи результатов скрещиваний: Р — родители; F — потомство, число внизу или сразу после буквы указывает на порядковый номер поколения (F1 — гибриды первого поколения — прямые потомки родителей, F2 — гибриды второго поколения — возникают в результате скрещивания между собой гибридов F1); × — значок скрещивания; G — мужская особь; E — женская особь; A — доминантный ген, а — рецессивный ген; АА — гомозигота по доминанте, аа — гомозигота по рецессиву, Аа — гетерозигота.
Закон единообразия гибридов первого поколения, или первый закон Менделя
Другие материалы по теме:
Успеху работы Менделя способствовал удачный выбор объекта для проведения скрещиваний — различные сорта гороха. Особенности гороха: 1) относительно просто выращивается и имеет короткий период развития; 2) имеет многочисленное потомство; 3) имеет большое количество хорошо заметных альтернативных признаков (окраска венчика — белая или красная; окраска семядолей — зеленая или желтая; форма семени — морщинистая или гладкая; окраска боба — желтая или зеленая; форма боба — округлая или с перетяжками; расположение цветков или плодов — по всей длине стебля или у его верхушки; высота стебля — длинный или короткий); 4) является самоопылителем, в результате чего имеет большое количество чистых линий, устойчиво сохраняющих свои признаки из поколения в поколение.
Опыты Менделя были тщательно продуманы. Если его предшественники пытались изучить закономерности наследования сразу многих признаков, то Мендель свои исследования начал с изучения наследования всего лишь одной пары альтернативных признаков.
Мендель взял сорта гороха с желтыми и зелеными семенами и произвел их искусственное перекрестное опыление: у одного сорта удалил тычинки и опылил их пыльцой другого сорта. Гибриды первого поколения имели желтые семена. Аналогичная картина наблюдалась и при скрещиваниях, в которых изучалось наследование других признаков: при скрещивании растений, имеющих гладкую и морщинистую формы семян, все семена полученных гибридов были гладкими, от скрещивания красноцветковых растений с белоцветковыми все полученные — красноцветковые. Мендель пришел к выводу, что у гибридов первого поколения из каждой пары альтернативных признаков проявляется только один, а второй как бы исчезает. Проявляющийся у гибридов первого поколения признак Мендель назвал доминантным, а подавляемый — рецессивным.
При моногибридном скрещивании гомозиготных особей, имеющих разные значения альтернативных признаков, гибриды являются единообразными по генотипу и фенотипу.
( А — желтый цвет горошин, а — зеленый цвет горошин)
| Р | ♀ AA желтые | × | ♂ аа зеленые | ||
| Типы гамет | А | а | |||
| F1 | Aа желтые 100% | ||||
Закон расщепления, или второй закон Менделя
Г. Мендель дал возможность самоопылиться гибридам первого поколения. У полученных таким образом гибридов второго поколения проявился не только доминантный, но и рецессивный признак. Результаты опытов приведены в таблице.
| Признаки | Доминантные | Рецессивные | Всего | ||
|---|---|---|---|---|---|
| Число | % | Число | % | ||
| Форма семян | 5474 | 74,74 | 1850 | 25,26 | 7324 |
| Окраска семядолей | 6022 | 75,06 | 2001 | 24,94 | 8023 |
| Окраска семенной кожуры | 705 | 75,90 | 224 | 24,10 | 929 |
| Форма боба | 882 | 74,68 | 299 | 25,32 | 1181 |
| Окраска боба | 428 | 73,79 | 152 | 26,21 | 580 |
| Расположение цветков | 651 | 75,87 | 207 | 24,13 | 858 |
| Высота стебля | 787 | 73,96 | 277 | 26,04 | 1064 |
| Всего: | 14949 | 74,90 | 5010 | 25,10 | 19959 |
Анализ данных таблицы позволил сделать следующие выводы:
- единообразия гибридов во втором поколении не наблюдается: часть гибридов несет один (доминантный), часть — другой (рецессивный) признак из альтернативной пары;
- количество гибридов, несущих доминантный признак, приблизительно в три раза больше, чем гибридов, несущих рецессивный признак;
- рецессивный признак у гибридов первого поколения не исчезает, а лишь подавляется и проявляется во втором гибридном поколении.
Явление, при котором часть гибридов второго поколения несет доминантный признак, а часть — рецессивный, называют расщеплением. Причем, наблюдающееся у гибридов расщепление не случайное, а подчиняется определенным количественным закономерностям. На основе этого Мендель сделал еще один вывод: при скрещивании гибридов первого поколения в потомстве происходит расщепление признаков в определенном числовом соотношении.
При моногибридном скрещивании гетерозиготных особей у гибридов имеет место расщепление по фенотипу в отношении 3:1, по генотипу 1:2:1.
Генетическая схема закона расщепления Менделя
( А — желтый цвет горошин, а — зеленый цвет горошин):
| P | ♀ Aa желтые | × | ♂ Aa желтые | ||
| Типы гамет | A | a | A | a | |
| F2 | AA желтые | Aa желтые 75% | Aa желтые | aa зеленые 25% | |
Закон чистоты гамет
Купить проверочные работы
и тесты по биологии




- за формирование признаков отвечают какие-то дискретные наследственные факторы;
- организмы содержат два фактора, определяющих развитие признака;
- при образовании гамет в каждую из них попадает только один из пары факторов;
- при слиянии мужской и женской гамет эти наследственные факторы не смешиваются (остаются чистыми).
В 1909 году В. Иогансен назовет эти наследственные факторы генами, а в 1912 году Т. Морган покажет, что они находятся в хромосомах.
| Р | ♀ Аа желтые | × | ♂ aа зеленые |
| Типы гамет | A a | a | |
| F | Аа желтые 50% | аa зеленые 50% |
Цитологические основы первого и второго законов Менделя
Во времена Менделя строение и развитие половых клеток не было изучено, поэтому его гипотеза чистоты гамет является примером гениального предвидения, которое позже нашло научное подтверждение.
Явления доминирования и расщепления признаков, наблюдавшиеся Менделем, в настоящее время объясняются парностью хромосом, расхождением хромосом во время мейоза и объединением их во время оплодотворения. Обозначим ген, определяющий желтую окраску, буквой А , а зеленую — а . Поскольку Мендель работал с чистыми линиями, оба скрещиваемых организма — гомозиготны, то есть несут два одинаковых аллеля гена окраски семян (соответственно, АА и аа ). Во время мейоза число хромосом уменьшается в два раза, и в каждую гамету попадает только одна хромосома из пары. Так как гомологичные хромосомы несут одинаковые аллели, все гаметы одного организмы будут содержать хромосому с геном А , а другого — с геном а .
При оплодотворении мужская и женская гаметы сливаются, и их хромосомы объединяются в одной зиготе. Получившийся от скрещивания гибрид становится гетерозиготным, так как его клетки будут иметь генотип Аа ; один вариант генотипа даст один вариант фенотипа — желтый цвет горошин.
У гибридного организма, имеющего генотип Аа во время мейоза, хромосомы расходятся в разные клетки и образуется два типа гамет — половина гамет будет нести ген А , другая половина — ген а . Оплодотворение — процесс случайный и равновероятный, то есть любой сперматозоид может оплодотворить любую яйцеклетку. Поскольку образовалось два типа сперматозоидов и два типа яйцеклеток, возможно возникновение четырех вариантов зигот. Половина из них — гетерозиготы (несут гены А и а ), 1/4 — гомозиготы по доминантному признаку (несут два гена А ) и 1/4 — гомозиготы по рецессивному признаку (несут два гена а ). Гомозиготы по доминанте и гетерозиготы дадут горошины желтого цвета (3/4), гомозиготы по рецессиву — зеленого (1/4).
Закон независимого комбинирования (наследования) признаков, или третий закон Менделя
Организмы отличаются друг от друга по многим признакам. Поэтому, установив закономерности наследования одной пары признаков, Г. Мендель перешел к изучению наследования двух (и более) пар альтернативных признаков. Для дигибридного скрещивания Мендель брал гомозиготные растения гороха, отличающиеся по окраске семян (желтые и зеленые) и форме семян (гладкие и морщинистые). Желтая окраска ( А ) и гладкая форма ( В ) семян — доминантные признаки, зеленая окраска ( а ) и морщинистая форма ( b ) — рецессивные признаки.
Скрещивая растение с желтыми и гладкими семенами с растением с зелеными и морщинистыми семенами, Мендель получил единообразное гибридное поколение F1 с желтыми и гладкими семенами. От самоопыления 15-ти гибридов первого поколения было получено 556 семян, из них 315 желтых гладких, 101 желтое морщинистое, 108 зеленых гладких и 32 зеленых морщинистых (расщепление 9:3:3:1).
Анализируя полученное потомство, Мендель обратил внимание на то, что: 1) наряду с сочетаниями признаков исходных сортов (желтые гладкие и зеленые морщинистые семена), при дигибридном скрещивании появляются и новые сочетания признаков (желтые морщинистые и зеленые гладкие семена); 2) расщепление по каждому отдельно взятому признаку соответствует расщеплению при моногибридном скрещивании. Из 556 семян 423 были гладкими и 133 морщинистыми (соотношение 3:1), 416 семян имели желтую окраску, а 140 — зеленую (соотношение 3:1). Мендель пришел к выводу, что расщепление по одной паре признаков не связано с расщеплением по другой паре. Для семян гибридов характерны не только сочетания признаков родительских растений (желтые гладкие семена и зеленые морщинистые семена), но и возникновение новых комбинаций признаков (желтые морщинистые семена и зеленые гладкие семена).
При дигибридном скрещивании дигетерозигот у гибридов имеет место расщепление по фенотипу в отношении 9:3:3:1, по генотипу в отношении 4:2:2:2:2:1:1:1:1, признаки наследуются независимо друг от друга и комбинируются во всех возможных сочетаниях.
| Р | ♀ АABB желтые, гладкие | × | ♂ aаbb зеленые, морщинистые |
| Типы гамет | AB | ab | |
| F1 | AaBb желтые, гладкие, 100% | ||
| P | ♀ АaBb желтые, гладкие | × | ♂ AаBb желтые, гладкие |
| Типы гамет | AB Ab aB ab | AB Ab aB ab | |
Генетическая схема закона независимого комбинирования признаков:
| Гаметы: | ♂ | AB | Ab | aB | ab |
| ♀ | |||||
| AB | AABB желтые гладкие | AABb желтые гладкие | AaBB желтые гладкие | AaBb желтые гладкие | |
| Ab | AABb желтые гладкие | AАbb желтые морщинистые | AaBb желтые гладкие | Aabb желтые морщинистые | |
| aB | AaBB желтые гладкие | AaBb желтые гладкие | aaBB зеленые гладкие | aaBb зеленые гладкие | |
| ab | AaBb желтые гладкие | Aabb желтые морщинистые | aaBb зеленые гладкие | aabb зеленые морщинистые | |
Анализ результатов скрещивания по фенотипу: желтые, гладкие — 9/16, желтые, морщинистые — 3/16, зеленые, гладкие — 3/16, зеленые, морщинистые — 1/16. Расщепление по фенотипу 9:3:3:1.
Анализ результатов скрещивания по генотипу: AaBb — 4/16, AABb — 2/16, AaBB — 2/16, Aabb — 2/16, aaBb — 2/16, ААBB — 1/16, Aabb — 1/16, aaBB — 1/16, aabb — 1/16. Расщепление по генотипу 4:2:2:2:2:1:1:1:1.
Если при моногибридном скрещивании родительские организмы отличаются по одной паре признаков (желтые и зеленые семена) и дают во втором поколении два фенотипа (2 1 ) в соотношении (3 + 1) 1 , то при дигибридном они отличаются по двум парам признаков и дают во втором поколении четыре фенотипа (2 2 ) в соотношении (3 + 1) 2 . Легко посчитать, сколько фенотипов и в каком соотношении будет образовываться во втором поколении при тригибридном скрещивании: восемь фенотипов (2 3 ) в соотношении (3 + 1) 3 .
Если расщепление по генотипу в F2 при моногибридном поколении было 1:2:1, то есть было три разных генотипа (3 1 ), то при дигибридном образуется 9 разных генотипов — 3 2 , при тригибридном скрещивании образуется 3 3 — 27 разных генотипов.
Третий закон Менделя справедлив только для тех случаев, когда гены анализируемых признаков находятся в разных парах гомологичных хромосом.
Цитологические основы третьего закона Менделя
Пусть А — ген, обусловливающий развитие желтой окраски семян, а — зеленой окраски, В — гладкая форма семени, b — морщинистая. Скрещиваются гибриды первого поколения, имеющие генотип АаВb . При образовании гамет из каждой пары аллельных генов в гамету попадает только один, при этом в результате случайного расхождения хромосом в первом делении мейоза ген А может попасть в одну гамету с геном В или с геном b , а ген а — с геном В или с геном b . Таким образом, каждый организм образует четыре сорта гамет в одинаковом количестве (по 25%): АВ , Ab , aB , ab . Во время оплодотворения каждый из четырех типов сперматозоидов может оплодотворить любую из четырех типов яйцеклеток. В результате оплодотворения возможно появление девяти генотипических классов, которые дадут четыре фенотипических класса.
Новая наука об обществе зарождалась не только на базе общих рассуждений о сущности социальных явлений, сохранения теории теологического происхождения общества, но и на основе анализа статистических данных, характеризующих общественные явления и процессы (Л.Кетле, К.Маркс, Д.Милль и др.); на основе более тщательной проработки самих логических приемов, с помощью которых анализировались эти явления: формулирование основных положений материалистической диалектики и ее применения к анализу современного капиталистического общества XIX века (К.Маркс, Ф.Энгельс, формулирования концепции преобладания социального над психологическим (Э.Дюркгейм) и наоборот (Г.Лебон, Г.Тард, Ф.Тённис), концепции преобладания смысловых, значимых элементов социальной жизни над другими (понимающая социология В.Дильтея, Г.Зиммеля, М.Вебера) и др.; на основе аналогии с законами естественных наук (социальный органицизм Г.Спенсера, социальный дарвинизм У.Беджгота, Л. Гумпловича, расово-антропологическая школа Ж.Габино и др.).
Ломбер Адольф Жак Кетле (1796 – 1874)
Выдающийся франко-бельгийский ученый: математик, астроном, географ, социолог, основатель национальных статистических обществ Англии и Франции, организатор Первого международного статистического конгресса (Брюссель, 1853 г.). Основные социологические идеи Кетле изложены в трудах: “Социальная физика или опыт исследования о развитии человеческих способностей” (1835 г., т.1-2), “Социальная система и законы, ею управляющие” (1866 г.).
Кетле не считал себя позитивистом и, по-видимому, не был знаком с трудами Конта. Тем не менее, основные идеи “социальной физики” Кетле лежали в русле позитивизма. В отличие от Конта и многих других социальных философов, разрабатывавших глобальные реформаторские проекты XIX века, Кетле скорее был социологом-прикладником, сознательно ограничивая область своих интересов частными, но социально значимыми проблемами.
Не претендуя на роль великого реформатора, Кетле привнес в социологию не только новые теоретико-методологические идеи, но и новый образец исследовательской деятельности, ориентированные на конкретные прикладные проблемы. Возражая против противопоставления природных и социальных явлений, он приводит множество примеров, свидетельствующих о том, что даже самые “интимные” события (супружество, преступления, самоубийства и т.п.) повторяются в общественной жизни с неизменной закономерностью.
Кетле существенным образом обогатил возможности применения статистических методов в социологии, первым перешел от анализа простых распределений к исследованию корреляционных связей. Исходя из статистической трактовки социальных законов, Кетле выделил два основных вида причинно-следственных связей и действующих факторов: социальные константы, т.е. факторы, воздействующие на социальную ситуацию постоянным образом (пол, возраст, профессия, религиозные и хозяйственные институты и т.д.), и случайные факторы, действия которых вносят отклонения от средней тенденции. Задача научной социологии состоит в том, чтобы методами статистики изменить границы этих отклонений и выявить определяющие их факторы. Для этих целей Кетле разрабатывал оригинальные методики многомерного корреляционного анализа. Социальные законы Кетле понимал, таким образом, как законы больших чисел, статистическую зависимость между случайными социальными событиями.
Статистический метод у Кетле выступал как метод социологии, лучше всех других выражающий специфику ее предмета исследования. Отсюда и знаменитая “концепция статистического человека”, некого среднего представителя народа, класса социальной группы. В социологическом плане “статистический человек” представлял норму, т.е. более желательное состояние совокупности людей. Полнота и точность знаний об обществе и человеке в трактовке Кетле зависят от технических возможностей статистики, а не от принципиальных трудностей, связанных с отличием социальных явлений от природных.

Кетле внес исключительно большой вклад в разработку методов статистического анализа в социологии, социальной, демографической, медицинской статистики, особенно методов измерения качественных характеристик. По сути, он создал - под именем “социальная физика” - социологию, значительно близкую к прикладной социологии XX века. В истории социальных учений Кетле вошел как один из основателей эмпирической, прикладной школы в социологии.
Классическая европейская социология середины XIX начала XX века
Ø Ломбер Адольф Жак Кетле (1796 – 1874),
Ø Карл Маркс (1818 –1883), Фридрих Энгельс (1820 – 1895),
Ø Герберт Спенсер (1820 – 1903),
Ø Людвиг Гумплович (1838 – 1909 гг.),
Ø Жозеф Артур Габино (1816-1882 гг.),
Ø Макс Вебер (1864 – 1920 гг.),
Ø Габриэль Тард (1843-1904 гг.),
Ø Фердинанд Тённис (1855-1936 гг.)
Новая наука об обществе зарождалась не только на базе общих рассуждений о сущности социальных явлений, сохранения теории теологического происхождения общества, но и на основе анализа статистических данных, характеризующих общественные явления и процессы (Л.Кетле, К.Маркс, Д.Милль и др.); на основе более тщательной проработки самих логических приемов, с помощью которых анализировались эти явления: формулирование основных положений материалистической диалектики и ее применения к анализу современного капиталистического общества XIX века (К.Маркс, Ф.Энгельс, формулирования концепции преобладания социального над психологическим (Э.Дюркгейм) и наоборот (Г.Лебон, Г.Тард, Ф.Тённис), концепции преобладания смысловых, значимых элементов социальной жизни над другими (понимающая социология В.Дильтея, Г.Зиммеля, М.Вебера) и др.; на основе аналогии с законами естественных наук (социальный органицизм Г.Спенсера, социальный дарвинизм У.Беджгота, Л. Гумпловича, расово-антропологическая школа Ж.Габино и др.).
Ломбер Адольф Жак Кетле (1796 – 1874)
Выдающийся франко-бельгийский ученый: математик, астроном, географ, социолог, основатель национальных статистических обществ Англии и Франции, организатор Первого международного статистического конгресса (Брюссель, 1853 г.). Основные социологические идеи Кетле изложены в трудах: “Социальная физика или опыт исследования о развитии человеческих способностей” (1835 г., т.1-2), “Социальная система и законы, ею управляющие” (1866 г.).
Кетле не считал себя позитивистом и, по-видимому, не был знаком с трудами Конта. Тем не менее, основные идеи “социальной физики” Кетле лежали в русле позитивизма. В отличие от Конта и многих других социальных философов, разрабатывавших глобальные реформаторские проекты XIX века, Кетле скорее был социологом-прикладником, сознательно ограничивая область своих интересов частными, но социально значимыми проблемами.
Не претендуя на роль великого реформатора, Кетле привнес в социологию не только новые теоретико-методологические идеи, но и новый образец исследовательской деятельности, ориентированные на конкретные прикладные проблемы. Возражая против противопоставления природных и социальных явлений, он приводит множество примеров, свидетельствующих о том, что даже самые “интимные” события (супружество, преступления, самоубийства и т.п.) повторяются в общественной жизни с неизменной закономерностью.
Кетле существенным образом обогатил возможности применения статистических методов в социологии, первым перешел от анализа простых распределений к исследованию корреляционных связей. Исходя из статистической трактовки социальных законов, Кетле выделил два основных вида причинно-следственных связей и действующих факторов: социальные константы, т.е. факторы, воздействующие на социальную ситуацию постоянным образом (пол, возраст, профессия, религиозные и хозяйственные институты и т.д.), и случайные факторы, действия которых вносят отклонения от средней тенденции. Задача научной социологии состоит в том, чтобы методами статистики изменить границы этих отклонений и выявить определяющие их факторы. Для этих целей Кетле разрабатывал оригинальные методики многомерного корреляционного анализа. Социальные законы Кетле понимал, таким образом, как законы больших чисел, статистическую зависимость между случайными социальными событиями.
Статистический метод у Кетле выступал как метод социологии, лучше всех других выражающий специфику ее предмета исследования. Отсюда и знаменитая “концепция статистического человека”, некого среднего представителя народа, класса социальной группы. В социологическом плане “статистический человек” представлял норму, т.е. более желательное состояние совокупности людей. Полнота и точность знаний об обществе и человеке в трактовке Кетле зависят от технических возможностей статистики, а не от принципиальных трудностей, связанных с отличием социальных явлений от природных.
Кетле внес исключительно большой вклад в разработку методов статистического анализа в социологии, социальной, демографической, медицинской статистики, особенно методов измерения качественных характеристик. По сути, он создал - под именем “социальная физика” - социологию, значительно близкую к прикладной социологии XX века. В истории социальных учений Кетле вошел как один из основателей эмпирической, прикладной школы в социологии.
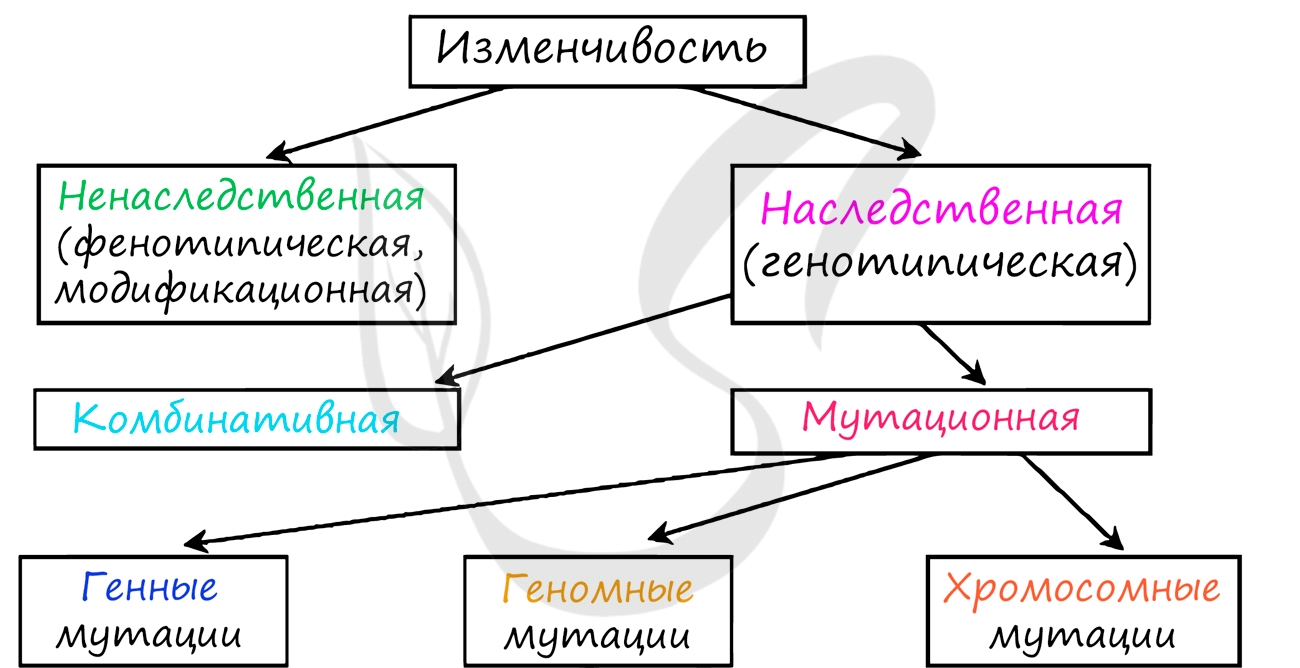
Под изменчивостью понимают способность организмов приобретать признаки и свойства, отличные от родительских, характерных для данного вида. Изменчивость является общим свойством всех живых систем и может выражаться в изменении как генотипа, так и фенотипа.
Традиционно различают ненаследственную и наследственную изменчивость.

Модификационная изменчивость
Модификационная (фенотипическая) изменчивость - изменения фенотипа организма, обусловленные влиянием факторов внешней среды. Данный вид изменчивости не приводит к изменениям генотипа особи - все изменения касаются только фенотипа.
Напомню, что генотипом называют генетическую конституцию - совокупность генов одного организма, полученных от родителей. Фенотип (греч. phаino - обнаруживаю) - совокупность наблюдаемых характеристик организма (любой морфологический, гистологический, биохимический, поведенческий признак).
Для модификационной изменчивости характерен групповой характер, она часто (но не всегда) служит приспособлением к условиям внешней среды. Известным примером модификационной изменчивости является изменение окраски шерсти у зайца-беляка в зависимости от сезона года.

Такое изменение окраски делает их более приспособленными, повышает выживаемость: заяц сливается с внешней средой и становится незаметен для хищников.
Однако не стоит забывать об относительности любой приспособленности: если среда резко изменится, то белый заяц на фоне темной земли станет легкой добычей для хищников.

Еще одним примером модификационной изменчивости служит изменение окраски шерсти у гималайских кроликов. Они рождаются полностью белыми, так как их эмбриональное развитие протекает в условиях повышенной температуры.
Однако в результате воздействия холода на разные участки их тела, шерсть начинает темнеть. В естественных условиях шерсть темная на ушах, носе, лапах и хвосте.
В эксперименте лед привязывают к спине, и через некоторое время шерсть на этом месте начинает темнеть. Это наглядно демонстрирует влияние внешней среды на проявление признака.

Вам известно, что человек, побывавший на солнце, получает его "отпечаток" - загар. Потемнение цвета кожи в данном случае связано с активной выработкой пигмента меланина, который защищает кожу и внутренние органы от УФ излучения.
Загар также является типичным примером модификационной изменчивости. Одни люди загорают быстро, у других этот процесс занимает гораздо больше времени - все дело в норме реакции.
Норма реакции
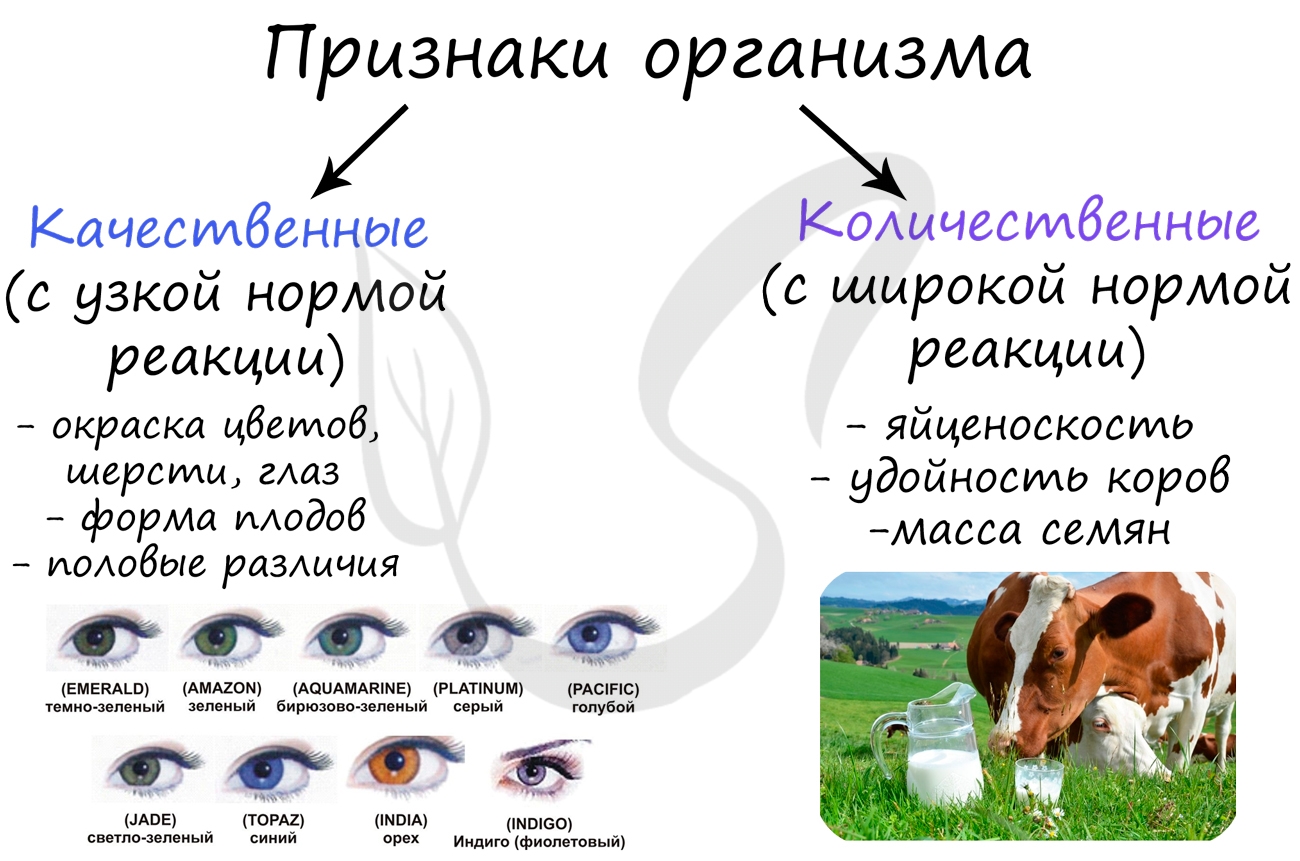
Нормой реакции называют генетически (наследственно) закрепленные пределы (границы) изменчивости признака. Принято говорить, что у каждого признака существует определенная норма реакции: она может быть узкой или широкой.
Узкая норма реакции характерна для признаков, которые относятся к качественным: форма глаза, желудка, сердца, размеры головного мозга, рост.
Количественные признаки имеют широкую норму реакцию и достаточно вариабельны в течение жизни: яйценоскость кур, удойность коров, вес, размер листьев.
- Причина изменения - влияние факторов внешней среды
- Изменения признаков организма не затрагивают генотип, происходят в соматических клетках и не передаются потомкам
- Изменение признаков ограничено в пределах нормы реакции, которая определяется генотипом
- Изменчивость носит групповой характер, характерна для многих особей (к примеру, сезонная изменчивость)
Наследственная изменчивость
Наследственная изменчивость (неопределенная, индивидуальная, генотипическая) - форма изменчивости, вызванная изменениями генотипа организма, которые могут быть связаны с мутационной или комбинативной изменчивостью.
В отличие от модификационной изменчивости, где затрагивается только фенотип (внешние проявления), генотипическая изменчивость затрагивает генотип, а это означает, что генетические изменения затрагивают и половые клетки, которые передаются потомству. Поэтому и называется она - наследственная.
Комбинативная изменчивость
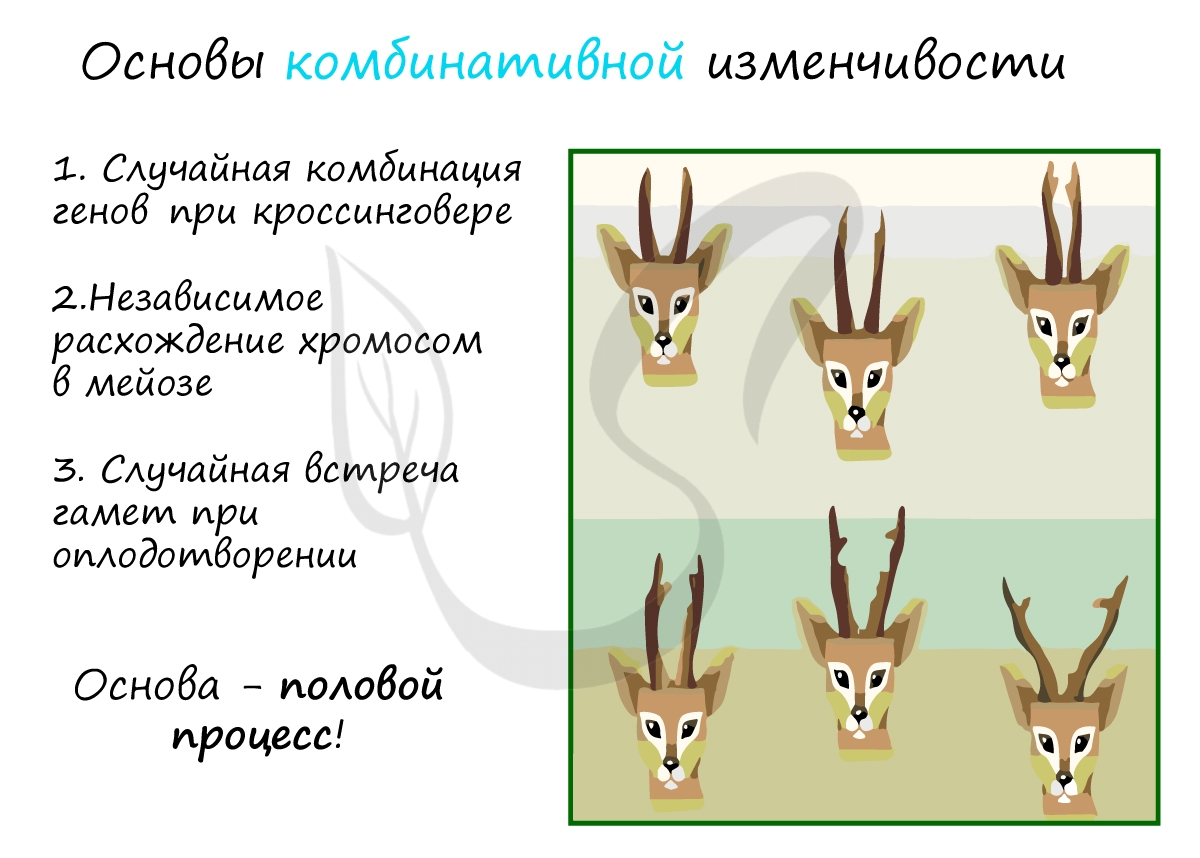
Комбинативная изменчивость возникает в результате появления у потомков новых сочетаний генов (комбинаций). Эти комбинации возникают во время мейоза в результате хорошо вам знакомого (я надеюсь!) кроссинговера - обмена участками между гомологичными хромосомами.
- Случайная комбинация генов в ходе кроссинговера
- Независимое расхождение хромосом в мейозе
- Случайная встреча гамет при оплодотворении
Я всегда говорю ученикам, что комбинативная изменчивость - это полная неопределенность: мы не знаем, какие комбинации возникнут между генами при кроссинговере, не знаем, какие хромосомы образуются и в какие гаметы они разойдутся, и, наконец, не знаем какие половые клетки (гаметы) встретятся при оплодотворении.
То, что мы отличаемся от своих родителей, и есть результат этих неопределенностей.

Мутационная изменчивость
Мутационная изменчивость связана с возникновением мутаций. Мутации (лат. mutatio - изменение) - внезапные, возникающие спонтанно или вызванные мутагенами наследуемые изменения генетического материала, приводящие к изменению тех или иных признаков организма.
- Мутации - резкие спонтанные изменения генотипа
- Стойкие, передаются потомкам через половые клетки (гаметы)
- Ненаправленные. Большинство мутаций - вредные (часть из них летальные), лишь очень небольшая часть носит полезный приспособительный характер, мутации также могут быть безразличными (нейтральными) для организма
- Носят индивидуальный характер

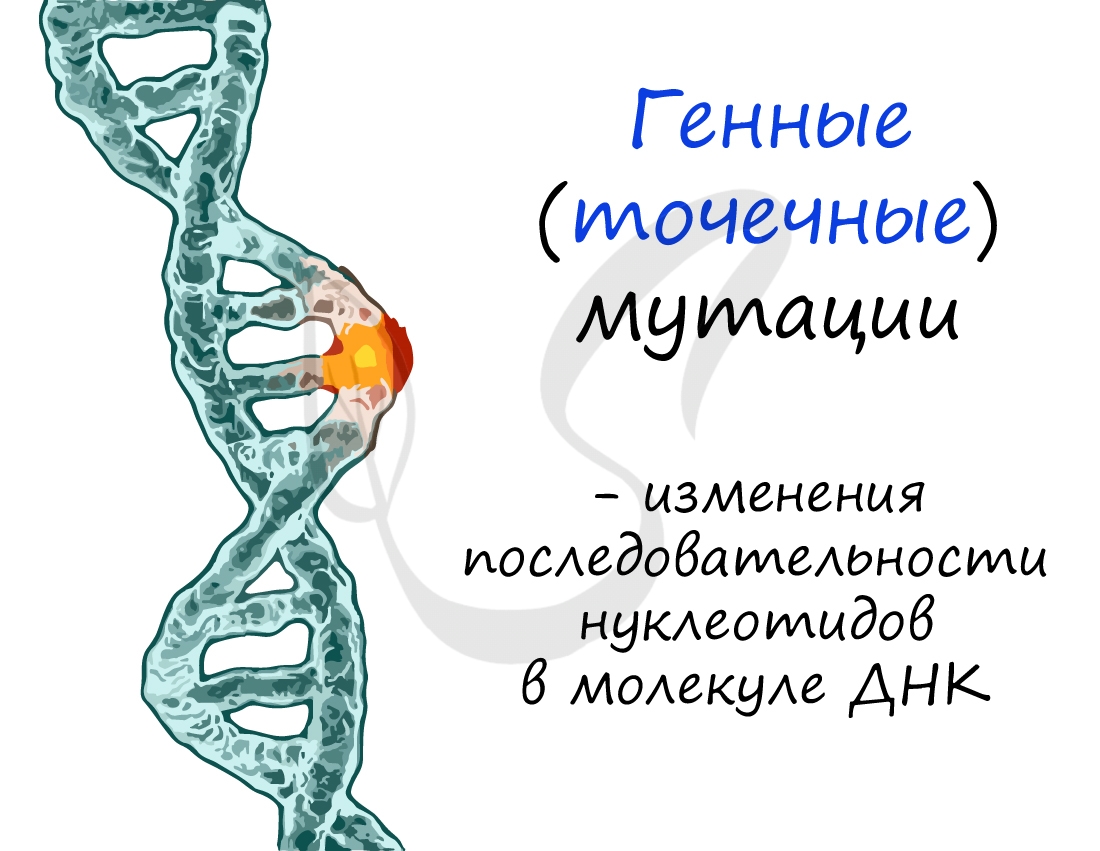
Изменения при генных мутациях происходят в последовательности нуклеотидов молекулы ДНК. Может случаться такое, что один или несколько нуклеотидов выпадают из ДНК (делеция), вставляются новые нуклеотиды, удваиваются имеющиеся нуклеотиды (дупликация).
Изменения ДНК ведут к тому, что в результате на рибосомах синтезируется белок с иной аминокислотной последовательностью. К примеру: изначально триплет ДНК "ТАЦ" кодировал аминокислоту "Мет", нуклеотид "Т" выпал из триплета произошла вставка нуклеотида "Г". В результате вместо аминокислоты "Мет" теперь синтезируется аминокислота Вал.
Новые аминокислоты могут поменять свойства белка, так что признак, за который он отвечает, будет меняться. Только что вы узнали об универсальной схеме - изменении фенотипа в результате изменений генотипа.
В результате хромосомных мутаций происходят структурные изменения хромосом (не следует путать с кроссинговером, который происходит в норме и подразумевает обмен участками между гомологичными хромосомами). Последствия хромосомных мутаций часто оказываются летальны.
В результате таких мутаций может происходить утрата (делеция) участка хромосомы, его удвоение (дупликация), поворот на 180° (инверсия), перенос участка одной хромосомы на другую (транслокация), перенос участка внутри одной хромосомы (транспозиция).

-
Автополиплоидию - кратное увеличение числа наборов хромосом
В результате таких мутаций количество хромосом увеличивается в кратное количество раз (2,3,4 и т.д.). В результате получаются организмы триплоиды, тетраплоиды и т.д. Иногда такие мутации вызывают искусственно, к примеру, в селекции растений. Известно, что у полиплоидов более крупные и сочные плоды.
В селекции полиплоидию у растений вызывают добавлением специального химического вещества - колхицина, который блокирует образование нитей веретена деления. Вследствие этого хромосомы не расходятся и остаются в одной клетке - набор хромосом увеличивается в 2 раза.

Имеет значение в процессе видообразования. Примером данной мутации может послужить отдаленная гибридизация (аутбридинг) пшеницы и ржи. Их генотип состоит из гаплоидного набора пшеницы (n) и гаплоидного набора ржи (m).
В результате такого скрещивания в 1875 году в Шотландии был получен первый искусственный стерильный гибрид - тритикале. Тритикале дает отличный урожай, в дальнейшем путем полиплоидии стерильность данного гибрида была преодолена.
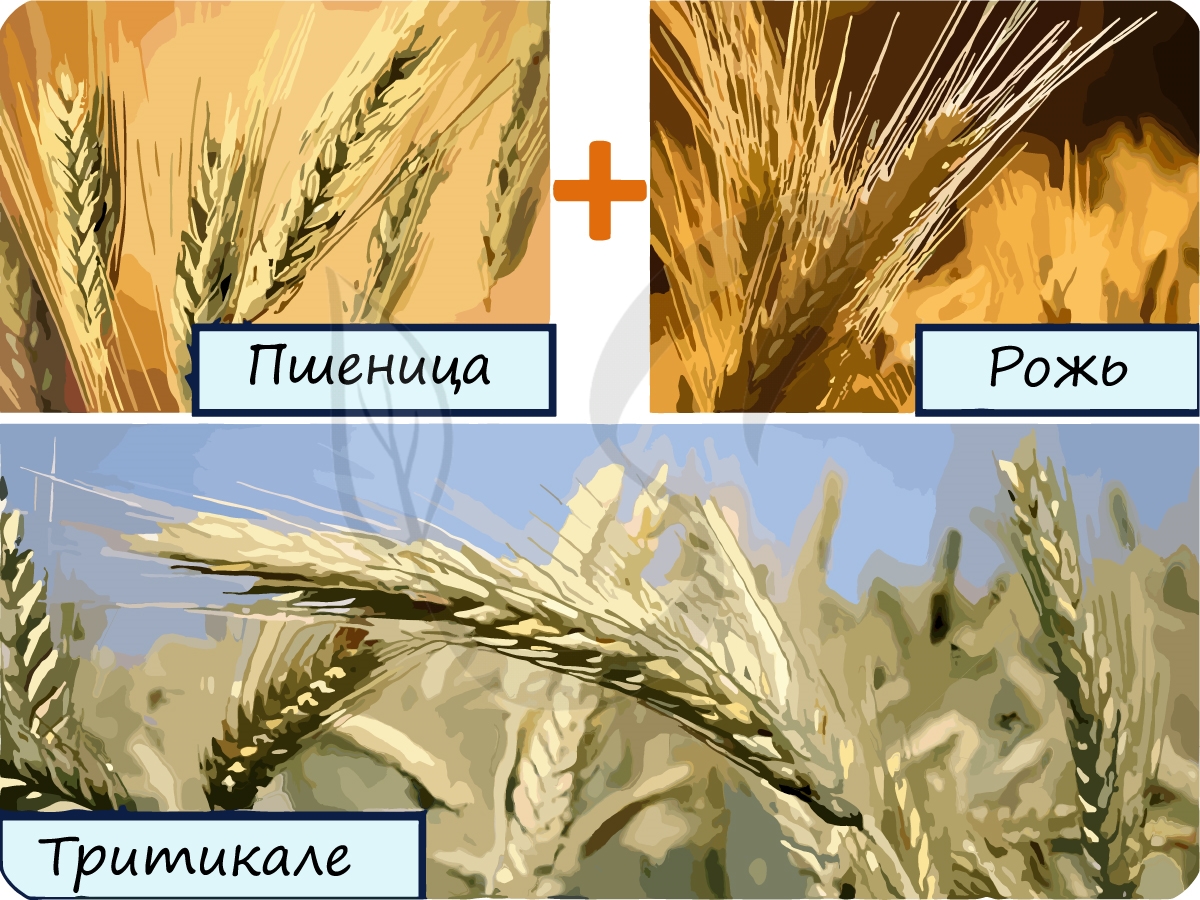
Также примером отдаленной гибридизации, соответственно и аллополиплоидии, является гибрид осла (самца) и лошади (самки) - мул. Это животное отличается большой выносливостью, но опять-таки бесплодное вследствие геномной мутации.

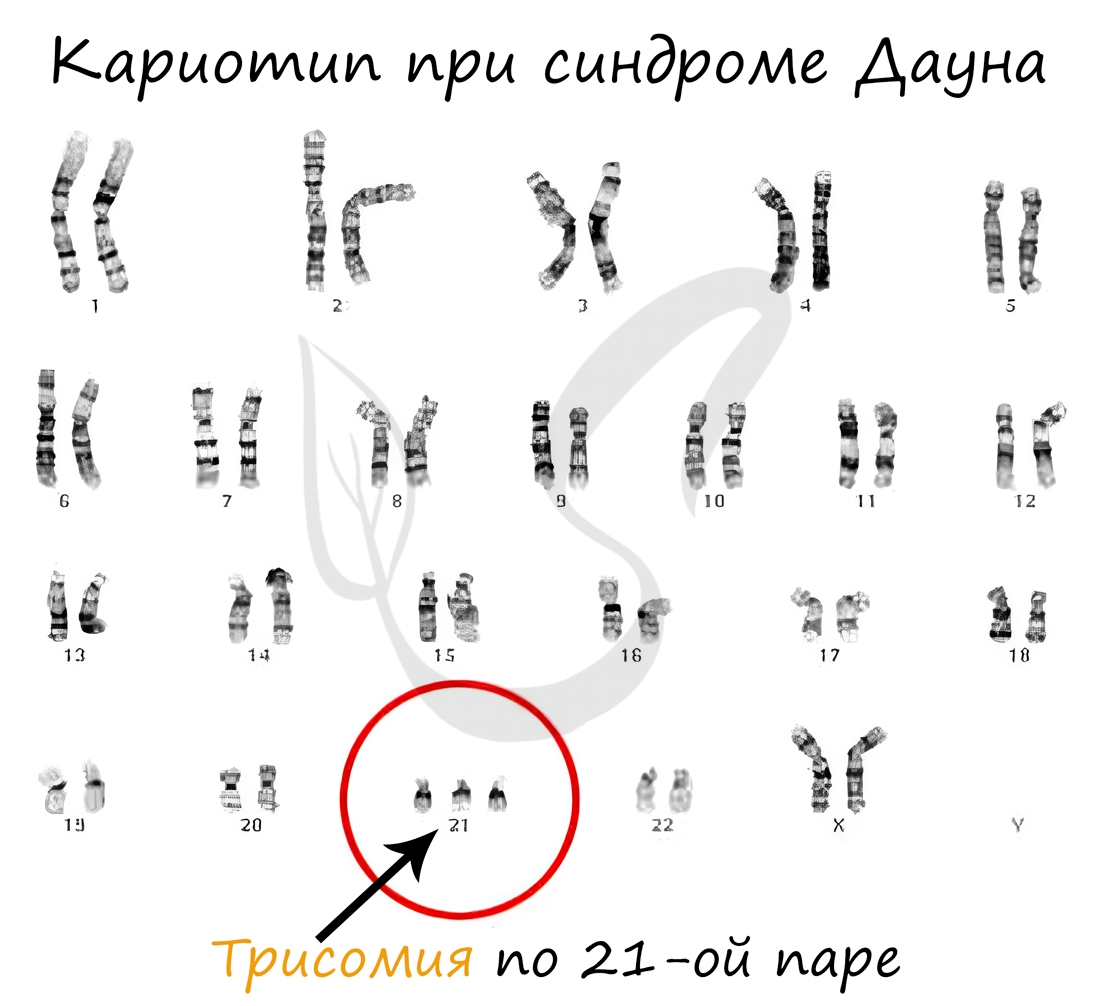
Анеуплоидия - изменение кариотипа (совокупность признаков хромосом), при котором число хромосом в клетках не кратно гаплоидному набору (n). Таким образом, в результате анеуплоидии отсутствует одна (или несколько) хромосом, либо же хромосомы имеются в избытке ("лишние" хромосомы).
В случае отсутствия в хромосомном наборе одной хромосомы говорят о моносомии, двух хромосом - нуллисомии. Если к паре хромосом добавляется одна лишняя, говорят о трисомии.
Наследственные болезни, в том числе связанные с геномными мутациями: синдром Шерешевского-Тёрнера, Дауна - мы более детально обсудим в следующей статье, которая посвящена наследственным заболеваниям.
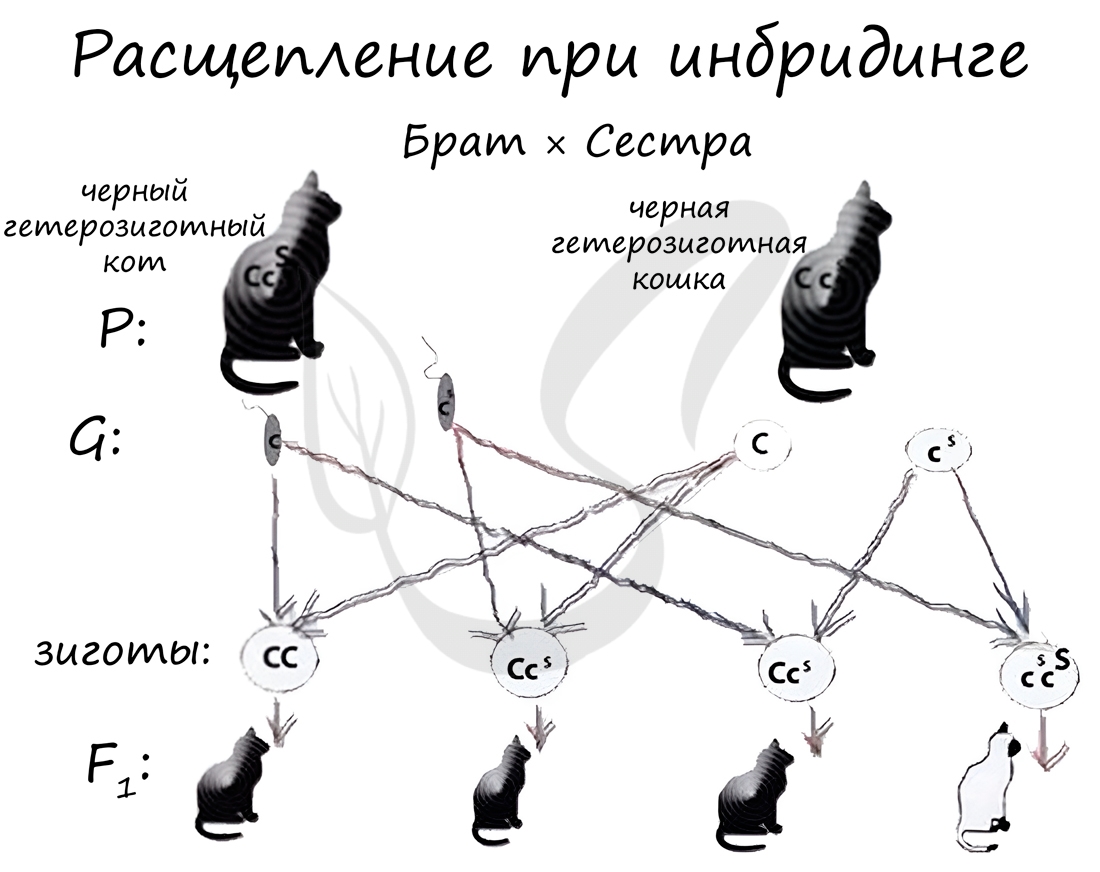
Раз уж мы затронули аутбридинг, то следует коснуться явления инбридинга и гетерозиса для их полного понимания.
Инбридинг (англ. in — в, внутри + breeding — разведение) - скрещивание близкородственных форм, в результате которого в ряду поколений увеличивается гомозиготность. С помощью инбридинга выводят чистые линии (AA, aa, BB, bb). Однако известно, что близкородственное скрещивание может приводить к проявлению рецессивных генов заболеваний и ослаблению потомства.
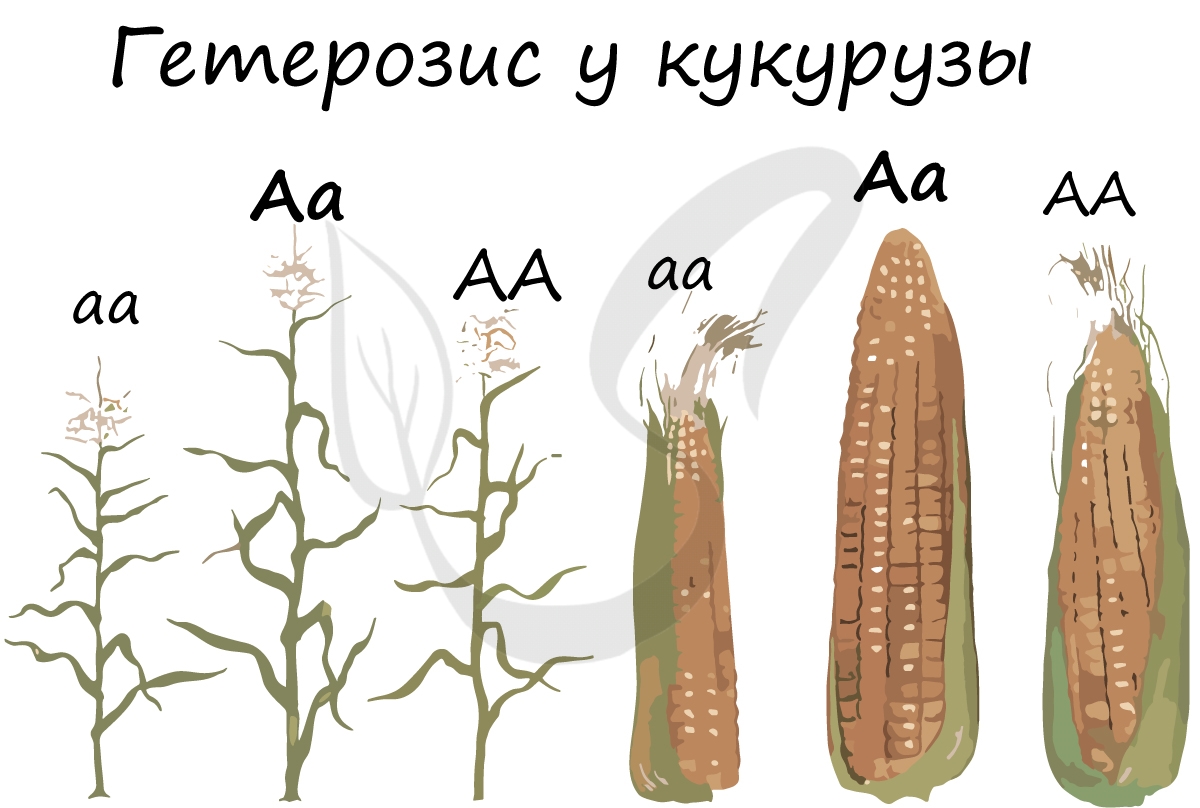
Гетерозис (греч. ἕτερος - другой + -ωσις - состояние) - явление увеличения жизнеспособности гибридов, вследствие унаследования ими различных вариантов аллельных генов от своих разнородных родителей. Увеличение жизнеспособности связывают с переходом генов в гетерозиготное состояние.
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Читайте также: