
Строение и значение подкорковой области кратко
Обновлено: 02.07.2024
комплекс образований головного мозга, расположенных между корой больших полушарий и продолговатым мозгом; участвуют в формировании всех поведенческих реакций человека и животных. В анатомическом плане к П. с. м. относят Зрительные бугры, Гипоталамус, лимбическую систему (См. Лимбическая система), четверохолмие, бледный шар и др. базальные нервные узлы, а в функциональном — ретикулярную формацию (См. Ретикулярная формация) ствола мозга и таламуса. См. также Подкорковые функции.
Большая советская энциклопедия. — М.: Советская энциклопедия . 1969—1978 .
Смотреть что такое "Подкорковые структуры мозга" в других словарях:
ПОДКОРКОВЫЕ СТРУКТУРЫ МОЗГА — отделы головного мозга, расположенные между корой больших полушарий и продолговатым мозгом. Оказывают активирующее влияние на кору, участвуют в формировании всех поведенческих реакций человека и животных, в поддержании мышечного тонуса и др … Большой Энциклопедический словарь
подкорковые структуры мозга — отделы головного мозга, расположенные между корой больших полушарий и продолговатым мозгом. Оказывают активирующее влияние на кору, участвуют в формировании всех поведенческих реакций человека и животных, в поддержании мышечного тонуса и др. * *… … Энциклопедический словарь
ПОДКОРКОВЫЕ СТРУКТУРЫ МОЗГА — отделы головного мозга, расположенные между корой больших полушарий и продолговатым мозгом Оказывают активирующее влияние на кору, участвуют в формировании всех поведенческих реакций человека и животных, в поддержании мышечного тонуса и др … Естествознание. Энциклопедический словарь
подкорка (подкорковые структуры мозга) — часть головного мозга, расположенная между корой больших полушарий и продолговатым мозгом. В нее входят: зрительные бугры, гипоталамус, лимбическая система и другие базальные нервные узлы, ретикулярная формация ствола мозга, таламус. П. участвует … Энциклопедический словарь по психологии и педагогике
Подкорковые функции — совокупность физиологических процессов, связанных с деятельностью отдельных подкорковых структур мозга (См. Подкорковые структуры мозга) или с их системой. С анатомической точки зрения к подкорковым относят все ганглионарные образования,… … Большая советская энциклопедия
ПОДКОРКОВЫЕ ФУНКЦИИ — ПОДКОРКОВЫЕ ФУНКЦИИ. Учение о функциях П. образований, развившееся на базе анат. клинических (по преимуществу) сравнительно анатомических и экспериментально физиологических исследований, насчитывает i.e много лет давности и не может считаться за … Большая медицинская энциклопедия
Кора больших полушарий головного мозга — слой серого вещества толщиной 1 5 мм, покрывающий полушария большого мозга млекопитающих животных и человека. Эта часть головного мозга (См. Головной мозг), развившаяся на поздних этапах эволюции животного мира, играет исключительно… … Большая советская энциклопедия
Кора головного мозга — Центральная нервная система (ЦНС) I. Шейные нервы. II. Грудные нервы. III. Поясничные нервы. IV. Крестцовые нервы. V. Копчиковые нервы. / 1. Головной мозг. 2. Промежуточный мозг. 3. Средний мозг. 4. Мост. 5. Мозжечок. 6. Продолговатый мозг. 7.… … Википедия
Префронтальная кора головного мозга — Префронтальная кора головного мозга … Википедия
ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ — (ВНД), интегративная деятельность высш. отделов центр. нервной системы (ЦНС), обеспечивающая поведение, т.е. оптимальное приспособление организма как целого к внеш. миру. Структурная основа ВНД у всех млекопитающих (в т. ч. у человека) кора… … Российская педагогическая энциклопедия

Весь контент iLive проверяется медицинскими экспертами, чтобы обеспечить максимально возможную точность и соответствие фактам.
У нас есть строгие правила по выбору источников информации и мы ссылаемся только на авторитетные сайты, академические исследовательские институты и, по возможности, доказанные медицинские исследования. Обратите внимание, что цифры в скобках ([1], [2] и т. д.) являются интерактивными ссылками на такие исследования.
Если вы считаете, что какой-либо из наших материалов является неточным, устаревшим или иным образом сомнительным, выберите его и нажмите Ctrl + Enter.
К подкорковым отделам головного мозга относятся зрительный бугор, базальные ядра в основании мозга (хвостатое ядро, чечевицеобразное ядро, состоящее из скорлупы, латерального и медиального бледных шаров); белое вещество головного мозга (полуовальный центр) и внутренняя капсула, а также гипоталамус. Патологические процессы (кровоизлияние, ишемия, опухоли и др.) часто развиваются одновременно в нескольких перечисленных образованиях, однако возможно и вовлечение только одного из них (полное или частичное).
Таламус (зрительный бугор). Важный подкорковый отдел афферентных систем; в нем прерываются проводящие пути всех видов чувствительности. Корковые отделы всех анализаторов имеют также обратные связи с таламусом. Афферентные и эфферентные системы обеспечивают взаимодействие с корой полушарий большого мозга. Таламус состоит из многочисленных ядер (всего их около 150), которые объединены в группы, различные по своему строению и функции (передние, медиальные, вентральные и задние группы ядер).
Таким образом, в таламусе можно выделить три основные функциональные группы ядер.
- Комплекс специфических, или релейных таламических ядер, через которые проводятся афферентные импульсы определенной модальности. К этим ядрам относятся передне-дорсальное и передне-вентральное ядра, группа вентральных ядер, латеральное и медиальное коленчатые тела, уздечка.
- Неспецифические таламические ядра не связаны с проведением афферентных импульсов какой-либо определенной модальности. Нейрональные связи ядер спроецированы в коре больших полушарий более диффузно, чем связи специфических ядер. К неспецифическим ядрам относятся: ядра средней линии и прилегающие к ним структуры (медиальное, субмедиальное и медиальное центральное ядра); медиальная часть вентрального ядра, медиальная часть переднего ядра, внутрипластинчатые ядра (парацентральное, латеральное центральное, парафасцикулярное и центральное срединное ядра); ядра, лежащие в параламинарной части (дорсальное медиальное ядро, переднее вентральное ядро), а также сетчатый комплекс таламуса,
- Ассоциативные ядра таламуса - это те ядра, которые получают раздражение от других ядер таламуса и передают эти влияния на ассоциативные области коры головного мозга. К этим образованиям таламуса относятся дорсальное медиальное ядро, латеральная группа ядер, подушка таламуса.
Таламус имеет многочисленные связи с другими отделами мозга. Корково-таламические связи образуют так называемые ножки таламуса. Передняя ножка таламуса образована волокнами, связывающими таламус с корой лобной области. Через верхнюю или среднюю ножку к таламусу идут пути от лобно-теменной области. Задняя ножка таламуса образована из волокон, идущих от подушки и наружного коленчатого тела к полю 17, а также височно-таламического пучка, соединяющего подушку с корой височно-затылочной области. Нижнее-внутренняя ножка состоит из волокон, соединяющих кору височной области с таламусом. Подбугорное ядро (люисово тело) относится к субталамической области промежуточного мозга. Оно состоит из однотипных мультиполярных клеток. К субталамической области относятся также поля Фореля и неопределенная зона (zona incetta). Поле Н 1 Фореля располагается под таламусом и включает волокна, соединяющие гипоталамус с полосатым телом - fasciculis thalami. Под полем Н 1 Фореля находится неопределенная зона, переходящая в перивентрикулярную зону желудочка. Под неопределенной зоной лежит поле H 2 Фореля, или fasciculus lenticularis, соединяющий бледный шар с подбугорным ядром и перивентрикулярными ядрами гипоталамуса.
К гипоталамусу (подбугорью) относятся поводок со спайкой, эпиталамическая спайка и эпифиз. В trigonum habenulae располагается gangl, habenulae, в котором выделяют два ядра: внутреннее, состоящее из мелких клеток, и наружное, в котором преобладают крупные клетки.
Поражения зрительного бугра вызывают прежде всего нарушения кожной и глубокой чувствительности. Возникает гемианестезия (или гипестезия) всех видов чувствительности: болевой, термической, суставно-мышечной и тактильной, больше в дистальных отделах конечностей. Гемигипестезии часто сочетаются с гиперпатией. Поражения таламуса (особенно медиальных его отделов) могут сопровождаться интенсивной болью - гемиалгией (мучительные ощущения колода, жжения) и различными вегетативно-кожными расстройствами.
Грубое нарушение суставно-мышечного чувства, а также нарушение мозжечково-таламических связей вызывают появление атаксии, которая обычно носит смешанный характер (сенситивный и мозжечковый).
Следствием поражения подкорковых отделов зрительного анализатора (латеральных коленчатых тел, подушки таламуса) объясняется возникновение гемианопсии - выпадения противоположных половин полей зрения.

[1], [2], [3], [4], [5], [6]

8 (495) 120-07-03
Заказать обратный звонок
Запись на прием к специалисту Центра
Психотерапевт Мартынов Сергей Егорович
Психолог, семейный психолог, клинический психолог Теперик Римма Фёдоровна
Психолог Копьёв Андрей Феликсович
Конфликтолог, психолог, клинический психолог Цуранова Наталья Александровна
Ночевкина Алёна Игоревна
Психолог, клинический психолог Алиева Лейла. Телесная терапия и арт-терапия.
Психолог Михайлова Анна Дмитриевна
Психолог, семейный психолог, коуч Волкова Татьяна
Детский психолог Горина Екатерина
Психолог Светлана Ткачева
Клинический психолог, психолог, психотерапевт Прокофьева Анна Вячеславовна
Основатель беатотерапии, психолог, клинический психолог Спиваковская Алла Семеновна
Психиатр Фролов Алексей Михайлович
Подростковый психолог Каравашкина Елена
Ведущий логопед Кухтина Алла Юрьевна
Подростковый психолог Максимов Алексей Вячеславович
Детский психолог Таранова Ирина Юрьевна
Психолог, юнгианский аналитик Юзьвак Екатерина Григорьевна
Психолог, клинический психолог, психоаналитический психотерапевт Ермушева Анастасия Алексеевна
Психиатр Медведев Владимир Эрнстович
Клинический психолог, психотерапевт, нейропсихолог Баринская Янина Сергеевна
Врач-психотерапевт Сивков Евгений Евгеньевич
Т.В. Мельникова
Нижний Новгород, Россия
Изучение роли подкорковых структур и корково-подкоркового взаимодействия в организации высших психических функций (ВПФ) остается актуальной проблемой.
Вместе с этим нейропсихологические исследования указывают на неоднородность активирующих систем, проявляющуюся снижением моторной, речевой, интеллектуальной активности при поражении левого полушария, особенностями эмоционально-личностной сферы при поражении правого.
Нарушения когнитивных функций при подкорковых поражениях зависят от распространенности патологического процесса и его этиологии (сосудистые или опухолевые заболевания мозга).
Данные о специфическом участии подкорковых ядер в психической деятельности, формировании межполушарной асимметри на подкорковом уровне получены при изучении ВПФ у больных паркинсонсонизмом до и после стереотаксических деструкций (Корсакова, Московичюте 1985).
Роль подкорковых структур в формировании когнитивных нарушений анализируется при прогрессирующей деменции (Корсакова и др., 1991), при болезни Паркинсона (Захаров и др., 1999), артери-венозных глубинных структурах мозга (Буклина, 1999).
Преобладание дисрегуляторных расстройств (при лакунарных инсультах зрительного бугра) выявлено при сопоставлении нарушений высших психических функций и МРТ (Яхно и др., 2002).
Зрительный бугор считается стратегически важной областью для реализации когнитивных функций, поэтому изучение нейропсихологического дефицита при поражениях различных его отделов представляет интерес для исследователей.
Нейропсихологическим исследованием при ограниченных таламических инсультах в отечественной и зарубежной литературе посвящено небольшое число публикаций. Во всех работах прослеживаются тенденции связывать возникновение когнитивных нарушений с поражением определенных сосудистых бассейнов и ядер зрительного бугра (Калашникова и др., 1998, Яхно и др., 2002).
В остром периоде, а также при двусторонних парамедианных поражениях отмечают снижение уровня бодрствования, сопор, нарушение памяти типа Корсаковского синдрома, галлюцинаторный синдром. Клинические проявления, наблюдаемые в подростковом периоде, рассматриваются как следствие нейродинамических нарушений.
При лакунаторных поражениях зрительного бугра в левом полушарии отмечают нарушения речевой функции (плохую артикуляцию, замедленность речи, парафазии), интеллектуальные, зрительно-пространственные, мнестические нарушения.
Сложная функция зрительного бугра обусловлена наличием специфических и неспецифических, вторичных и третичных групп ядер, а также многочисленными связями: соматосенсорными и ретикулотамическими путями, связями с орбитальной и префронтальной, теменной и затылочной корой, поясной извилиной, маммилярными телами. Типичный (задне-латеральный) таламический синдром Джерина-Русси, возникающий в результате закупорки таламо-коленчатой артерии, характеризуется преходящими контрлатеральным гемипарезом, контрлатеральной гемианестезией для тактильной и особенно глубокой чувствительности, гемигипестезией болевой и температурной чувствительности, спонтанной болью в вовлеченных областях, гемиатаксией, астереогнозом, контрлатеральным хореоатетозом.
Передне-боковой таламический синдром связан с закупоркой таламо-перфорирующей артерии и включает интенционный тремор, хореоатетоз, таламическую кисть.
Сопоставление результатов использования высокочувствительного нейропсихологического метода и данных КТ при лакунарных инсультах зрительного бугра позволяет оценить основные составляющие формирования нейропсихологического синдрома при этой патологии и возможность латерализации функций на этом уровне.
Лакунарные таламические инсульты по данным компьютерной томографии выявлены у 7 больных (5 женщин и 2 мужчин). Среднии возраст обследованных – 57 лет.
В трех случаях при компьютерной томографии головного мозга имелись указания на локализацию процесса в подушке зрительного бугра, в боковых и средних отделах, в 4 случаях локализация очага в пределах зрительного бугра точно не была определена. Правополушарная локализация была установлена у 5 больных, левополушарная у 2.
Обследование проводилось через 1-3 недели после острого инсульта. Ни в одном случае не было отмечено грубых нарушений памяти, дезориентации в месте и времени, галлюцинаторного синдрома. Только у одного больного был выявлен синдром Дежерина-Русси, с явлениями гемигипестезии, атаксии, жгучими болями в контрлатеральной половине тела. У остальных – очаговая симптоматика была легко выражена и проявлялась легкими преходящими гемипарезами, непостоянными нарушениями чувствительности, атаксией.
Строение коры головного мозга: слои, клетки
Толщина коры больших полушарий (от греч. pallium — мантия) варьирует от 2 до 4 мм, наиболее тонкая ее часть находится в области первичной зрительной коры, а наиболее утолщенная — в первичной моторной области. Больше половины общей поверхности коры скрыто стенками борозд. Мозг содержит приблизительно 86 млрд, нейронов (кора больших полушарий содержит только 19% общего количества, но составляет 81% массы мозга), аналогичное число глиальных клеток и разветвленную капиллярную сеть.
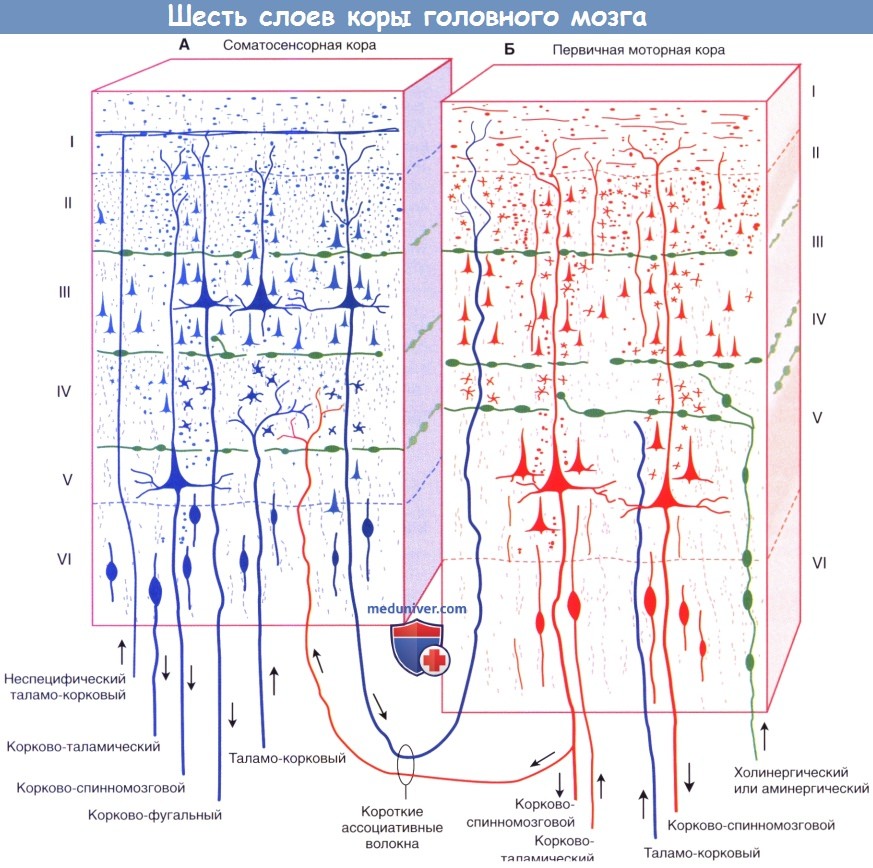
Шестислойное строение изокортекса коры головного мозга.
(А) Соматосенсорная кора. (Б) Первичная моторная кора, кортикальные пластинки обозначены цифрами от I до VI.
Клеточные пластинки новой коры (неокортекса):
I. Молекулярный слой образован концами апикальных дендритов пирамидных клеток и наиболее дистальными ветвями аксонов, проходящих к коре от интраламинарных ядер таламуса.
II. Наружный зернистый слой образован мелкими пирамидными и звездчатыми клетками.
III. Наружный пирамидный слой состоит из средних пирамидных клеток и звездчатых клеток.
IV. Внутренний зернистый слой образован звездчатыми клетками, к которым подходят восходящие волокна от релейных таламических ядер. Звездчатые (зернистые) клетки особенно многочисленны в первичной соматосенсорной коре, первичной зрительной коре и первичной слуховой коре; к ним подходят афферентные чувствительные волокна. Эти отделы обозначают как гранулярная кора. Наоборот, первичная моторная кора — место начала корково-спинномозгового и корково-бульбарного путей, имеет относительно небольшое количество зернистых клеток в слое IV и большое число пирамидных клеток в слоях III и V, которые смазывают (перекрывают) отдельные слои. Эту область обозначают как агранулярная кора.
V. Внутренний пирамидный слой образован крупными пирамидными клетками, волокна которых направляются к полосатому телу, стволу мозга и спинному мозгу.
VI. Слой веретеновидных клеток образован видоизмененными пирамидными клетками, волокна которых направляются к таламусу.
Взаимодействие между группами колонн позволяет осуществлять более сложную деятельность, модели поведения или решать когнитивные задачи.
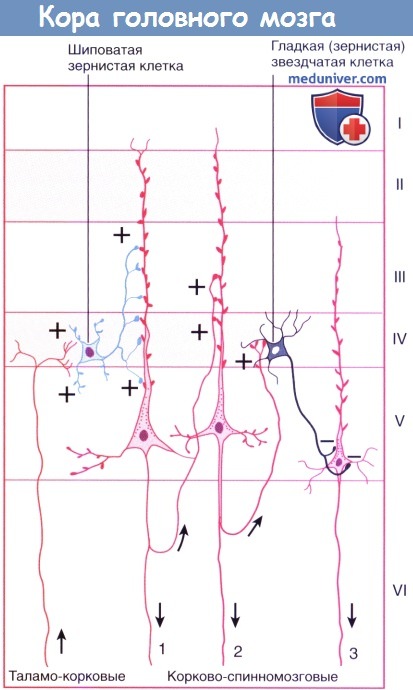
Восходящие/нисходящие проводящие пути. Стрелки указывают направление проведения импульса. Знаки +/- обозначают возбуждение/торможение.
Пирамидная клетка 1 возбуждается шиповатой зернистой клеткой; она возбуждает клетку 2 внутри своей колонны клеток; клетка 3 в соседней колонне тормозится гладкой зернистой клеткой.
в) Типы клеток. Морфологически кортикальные нейроны разделяют на две большие группы. Большая часть (60-85%) представлена пирамидными нейронами (из-за их формы) — единственными, волокна которых покидают кору (и к которым подходит большая часть волокон), что объясняет их альтернативное название — корковые проекционные нейроны; их волокна — возбуждающие глутаматергические. К оставшимся 15-40% нейронам относят непирамидные, или вставочные, нейроны; несмотря на то, их взаимосвязи не выходят за пределы коры, они осуществляют регуляцию и значительное влияние на ее деятельность; их тип — преимущественно тормозной ГАМК-ергический.
Внутри каждой группы можно выделить множество подгрупп в зависимости от морфологии, взаимосвязей, электрофизиологических свойств, типа развития, физиологических характеристик, молекулярных маркеров и т. д. (Примерами основных морфологических и функциональных клеточных типов служат пирамидные клетки, шиповатые зернистые клетки (измененные пирамидные клетки) и группа непирамидных тормозных вставочных нейронов.)
• Шиповатые зернистые клетки — один из вариантов атипичных пирамидных клеток, лежащих в слое IV и наиболее многочисленных в первичной сенсорной коре. Их шиповатые дендриты не выходят за пределы слоя IV, а аксоны могут подниматься или опускаться, образуя возбуждающие глутаматергические синаптические контакты с пирамидными клетками. На них переключается большая часть волокон восходящих таламических путей к слою IV с дальнейшим радиальным распространением.
Для организации их можно разделить на три крупных семейства в зависимости от экспрессируемых этими вставочными нейронами биомаркеров: парвальбумин, соматостатин и серотонин (5-гидрокситриптамин, 5НТ) 3а-рецептор (5HT3aR).
• Парвальбумин-экспрессирующие вставочные нейроны не имеют шигшков на дендритах. К ним подходят возбуждающие волокна от таламуса и коры, а тормозные — от других вставочных нейронов того же типа. Считают, что они играют роль в стабилизации активности корковых нейронных сетей. Как и в коре мозжечка, эти нейроны обладают фокусирующим действием на кору больших полушарий, тормозя слабо возбужденные колонны клеток. Клетки-канделябры (названы так из-за канделяброподобных групп аксоаксональных синаптических окончаний) наиболее многочисленны в слое II, образуют контакты с начальным сегментом аксона пирамидной клетки и играют ключевую роль в корково-корковых взаимодействиях.
• Соматостатин-экспрессирующие вставочные нейроны. Примером служат клетки Мартинотти, расположенные в пластинках V и VI, аксоны которых направляются к пластинке I. Получая сигналы от пирамидных клеток, они способны ограничивать их периферическое возбуждение и объединять несенсорную информацию, осуществляя, таким образом, регулирование обработки сигналов в дендритах их пирамидных клеток в зависимости от внешних обстоятельств.
• 5НТ3а-экспрессирующие вставочные нейроны — гетерогенная группа, к которой относят большую часть вставочных нейронов поверхностных корковых слоев. Получая информацию от корковых и таламических нейронов, они могут участвовать в процессах обучения, осуществляя влияние на корковые нейронные цепи. Он нейроглиеформных клеток (паутинообразных клеток)—одного из главных типов вставочных нейронов слоев II и III — веерообразно расходятся дендриты, обладающие уникальным свойством образования синапсов друг с другом и другими типами вставочных нейронов; этот факт указывает на их важнейшую роль в синхронизации корковых нейронных цепей. Другая морфологически гетерогенная группа вставочных нейронов, помимо ГАМК, экспрессирует вазоактивный интестинальный полипептид; другие типы вставочных нейронов этой группы экспрессируют также холецистокинин и другие пептидные рецепторы.
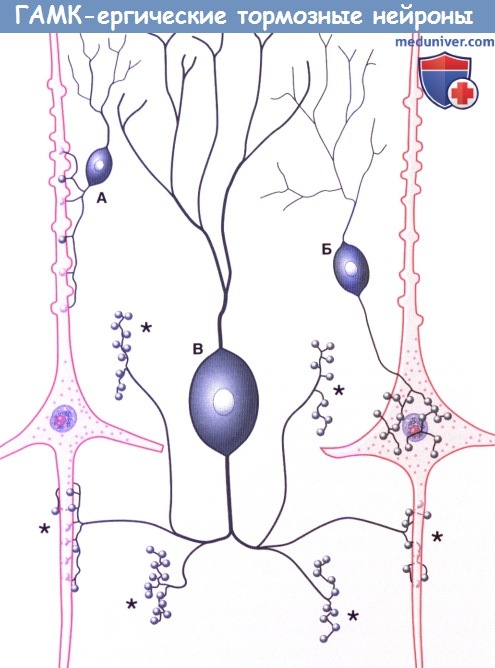
Три морфологических типа ГАМК-ергических тормозных нейронов:
А. Аксодендритическая клетка, образующая синапс с апикальным отделом пирамидного нейрона.
Б. Корзинчатая клетка, образующая аксоаксональные синапсы с пирамидными клетками.
В. Клетка-канделябр, формирующая аксоаксональные синапсы (*) с начальными сегментами аксонов двух пирамидных клеток, показанных здесь, и с начальными сегментами четырех других клеток, не показанных на рисунке.
(На основе DeFelipe, 1999, с разрешения автора.)
г) Афферентные волокна. Восходящие волокна к любому отделу коры могут иметь четыре различных источника (в первую очередь, корковые) и оканчиваются в разных отделах.
1. Длинные и короткие ассоциативные волокна от мелких и средних пирамидных клеток в слоях II и III из других корковых зон того же полушария.
2. Комиссуральные волокна от средних пирамидных клеток слоев II и III проходят через мозолистое тело от аналогичных или топографически идентичных участков противоположного полушария.
3. Таламо-корковые волокна от соответствующего специфического или ассоциативного ядра (например, волокна от вентрального заднего таламического ядра к соматосенсорной коре) и от дорсомедиального таламического ядра к предлобной (префронтальной) коре (описана ниже) оканчиваются в пластинке IV. Неспецифические таламо-корковые волокна от внутрипластинчатых ядер оканчиваются во всех пластинках.
4. Холинернические и аминергические волокна от базальных ядер, гипоталамуса и ствола мозга. Эти волокна обозначены на рисунке ниже зеленым цветом. Несмотря на то, что они распространены по всей коре, их возбуждение не сопровождается генерализованной или неспецифической реакцией. Анатомическая специфичность волокон (корковая, пластинчатая и клеточная) обусловливает возбуждение или торможение только определенных групп нейронов. Ядра их начала и нейромедиаторы:
• базальное ядро Мейнерта (базальные ядра мозга), ацетилхолин;
• серобугорно-сосцевидное ядро (задний отдел гипоталамуса), гистамин;
• черная субстанция, компактный слой (вентральный отдел покрышки среднего мозга), дофамин;
• ядра шва (средний мозг и ростральный отдел моста), серотонин;
• голубое пятно (ростральный отдел моста), норадреналин. Эти пять групп нейронов имеют особое значение в психиатрии и подробно рассмотрены в отдельной статье на сайте.
д) Эфферентные волокна. Единственные эфферентные волокна, покидающие кору больших полушарий, представлены возбуждающими аксонами пирамидных клеток. Одна часть аксонов пирамидных клеток соединяется с длинными и короткими ассоциативными волокнами, другая образует комиссуральные или проекционные пути. Ассоциативные и комиссуральные пути составляют большой объем белого вещества полушарий головного мозга.
• Примерами коротких ассоциативных путей (проходят между соседними зонами коры в составе поверхностного белого вещества в виде U-волокон) служат пути, направляющиеся в моторную кору от сенсорной коры и обратно. Примерами длинных ассоциативных путей служат волокна между префронтальной корой (лежащей кпереди от моторных зон коры) и ассоциативными сенсорными зонами. Источник этих волокон — пирамидные клетки, расположенные преимущественно в слоях II и III.
• Комиссуральные волокна мозга представлены исключительно аксонами пирамидных клеток, проходящих через мозолистое тело, переднюю и заднюю спайки (и другие мелкие спайки) к аналогичным участкам противоположного полушария (например, волокна от первичной корковой зоны направляются к соответствующей ассоциативной области другого полушария), а также к несоответствующим зонам [подобные комиссуральные связи отсутствуют между первичной зрительной корой, первичной соматосенсорной и моторной корой, иннервирующими дистальный отдел верхней конечности]. Волокна начинаются от пирамидных клеток, расположенных преимущественно в слоях II и III.
• Основная часть проекционных волокон от первичной сенсорной и моторной коры подходит к базальным ганглиям. К таламусу направляются волокна от всех отделов коры. К другим основным проводящим путям относят корково-мостовой (к ипсилатеральным ядрам моста), корково-ядерный (к двигательным и чувствительным ядрам черепных нервов моста и продолговатого мозга на противоположной стороне) и корково-спинномозговой. Источником этих волокон служат пирамидные клетки, расположенные преимущественно в слоях V и VI (отдающие волокна преимущественно к специфическим релейным ядрам таламуса).
Читайте также: