
Регуляция эмбрионального развития в кратком содержании
Обновлено: 05.07.2024
Сайт учителей биологии МБОУ Лицей № 2 г. Воронежа, РФ
Site biology teachers lyceum № 2 Voronezh city, Russian Federation
Регуляция эмбрионального развития
Как мы уже рассмотрели выше, материальные основы для регуляторных процессов, создающих из организма не бесформенную массу клеток, а хорошо согласованную систему органов и тканей, закладываются ещё в овогенезе.
В настоящее время большое значение в определении морфогенетических процессов развивающегося зародыша придают гомеозисным генам, регулирующим развитие анатомических особенностей сегментов тела. Первоначально открытые у дрозофилы, эти гены впоследствии были обнаружены практически у всех живых организмов. Их смысловая часть определяет структуру белка, получившего название гомеодомен . Учё ные доказали, что различные гомеодомены имеют специфическое сродство к определённым участкам ДНК. Активирующая транскрипцию функция гомеодоменов может быть модифицирована их взаимодействием с другими регуляторными белковыми молекулами.

Гомеодоменная регуляция анатомических особенностей. сегментов тела. Гены, кодирующие развитие органов на передне-задней оси тела, сходны у членистоногих и млекопитающих
Эмбриональная индукция. Наблюдения за оплодотворённой яйцеклеткой лягушки позволили проследить путь развития клеток, входящих в состав того или иного участка зародыша. Оказалось, что строго определённые клетки, занимающие соответствующее место в бластуле, дают начало строго определённым зачаткам органов. Удалось выяснить, какие группы клеток зародыша дают начало нервной трубке, хорде, мезодерме, кожному эпителию. А вот культивирование клеток зародыша вне организма ( in vitro ) не приводит к фор мированию типичных тканевых структур, которые могли бы образоваться из этих клеток. Чем же вызывается преобразование тех или иных клеток зародыша в конкретные ткани или органы?
Достижение эмбриональными закладками — тканями достаточной клеточной массы, зрелости, определяющей возможность синтеза регуляторных молекул, и компетентности для восприятия регуляторных стимулов обусловливает следующий этап эмбриональной регуляции путём индукции.
Явление эмбриональной индукции было открыто в 20-х гг. XX в. немецким эмбриологом Г. Шпеманом. На стадии ранней гаструлы он вырезал зачаток хорды вместе с участком зачатка мезодермы у одного зародыша амфибий и пересадил его другому зародышу под эктодерму, из которой должна была развиться кожа. В месте контакта пересаженного участка с эктодермой из зачатка кожного эпителия у второго зародыша возникла дополнительная нервная трубка, а из чужого зачатка — трансплантата — развивалась хорда и мезодерма. Все дополнительные структуры образуются помимо аналогичных нормально развивающихся образований зародыша. Такое влияние одного зачатка на другой, при котором первый — индуктор, определяет направление развития второго — компетентностного, получило название эмбриональной индукции .

Эмбриональная индукция. Схема опыта Г. Шпемана по пересадке зачатка хорды
Насколько важную роль играет эмбриональная индукция в развитии, показывает следующий опыт. Если на стадии ранней гаструлы полностью удалить зачаток хорды, то нервная трубка совсем не развивается. Эктодерма на спинной стороне зародыша, из которой в норме образуется нервная трубка, дифференцируется в кожный эпителий, и такой зародыш погибает.
При дальнейшем изучении развития зародышей оказалось, что зачаток хорды не только представляет собой индуктор нервной трубки, но и сам для своей дифференцировки нуждается в индуцирующем воздействии со стороны зачатка нервной системы. Во время эмбрионального развития имеет место не односторонняя индукция, а взаимодействие частей зародыша.
Современный этап изучения молекулярных механизмов эмбриональной индукции начался во второй половине 80-х гг. XX в., когда, благодаря развитию молекулярной биологии, учёным удалось связать индукционные процессы и дифференцировку клеток с активацией или подавлением работы определённых генов. Некоторые белки, синтезированные в результате экспрессии отдельных генов, оказались способными регулировать активность других генов, формируя в развитии целые каскады генетических программ, активизирующих или репрессирующих друг друга.
Иллюстрировать каскадный характер процессов эмбриональной индукции можно на примере развития глаза у позвоночных животных. Вырост второго мозгового пузыря — глазной бокал является и индуктором по отношению к прилегающей эктодерме, являющейся на этом этапе компетентной тканью. Результатом индуктивного воздействия является дифференцировка кожного эпителия в хрусталик, погружающийся внутрь тела. На следующем этапе уже формирующийся хрусталик выступает как индуктор по отношению к следующей порции эктодермального эпителия, направляя его развитие на образование роговицы.

Таким образом, эмбриональная индукция сопровождает весь процесс эмбриогенеза, причём практически каждый зачаток влияет на другой, определяя путь его развития, и сам подвергается индуцирующему воздействию. В самом скором времени к регуляции эмбрионального развития присоединяются формирующиеся нервная и эндокринная системы.
Эмбриональное (зародышевое) развитие охватывает процессы от первого деления зиготы до выхода из яйца или рождения и у большинства животных включает три основных этапа: дробление, гаструляцию и органогенез.
При дроблении дочерние клетки ( бластомеры ) не расходятся и не увеличиваются в размерах. С каждым следующим делением их размеры уменьшаются.

Яйцеклетки с небольшим запасом питательных веществ делятся полностью, т. е. происходит полное дробление. Если яйцеклетка содержит большое количество желтка, то наблюдается частичное дробление — делится только диск цитоплазмы с ядром, а сам желток остаётся без изменений (например, у птиц).
Бластула — это шарообразный зародыш, стенка которого ( бластодерма ) образована одним слоем клеток, а внутри — полость ( бластоцель ).
После дробления начинается гаструляция — часть клеток бластодермы перемещается внутрь зародыша. В результате этих перемещений образуется гаструла.
Гаструла — двухслойный зародыш, состоящий из двух зародышевых листков: наружного ( эктодермы ) и внутреннего ( энтодермы ).
У ланцетника образование гаструлы происходит в результате впячивания части бластодермы внутрь бластоцеля.
Внутренняя полость гаструлы называется первичной кишкой . Её связывает с внешней средой отверстие ( бластопор ), которое становится первичным ртом .

На стадии гаструлы (двух зародышевых листков) прекращается развитие двухслойных животных — губок и кишечнополостных.
У всех остальных животных развитие продолжается, и образуется третий зародышевый листок — мезодерма . Она формируется из энтодермы и всегда расположена между экто- и энтодермой в первичной полости тела.
Дальнейшая специализация клеток зародышевых листков обеспечивает формирование тканей и органов, т. е. гисто- и органогенез .
Из энтодермы образуется хорда — внутренний скелет в виде гибкого тяжа, расположенный на спинной стороне. Позже вместо хорды у позвоночных развивается позвоночник, и только у некоторых животных (например, у хрящевых рыб) её остатки сохраняются в течение всей жизни.
Из эктодермы, расположенной над самой хордой, выделяется нервная пластинка . Затем края пластинки поднимаются и смыкаются. Образуется нервная трубка — зачаток центральной нервной системы. Формируется нейрула .

Нервная трубка, хорда и кишечник создают осевой комплекс органов зародыша, который определяет двустороннюю симметрию тела.

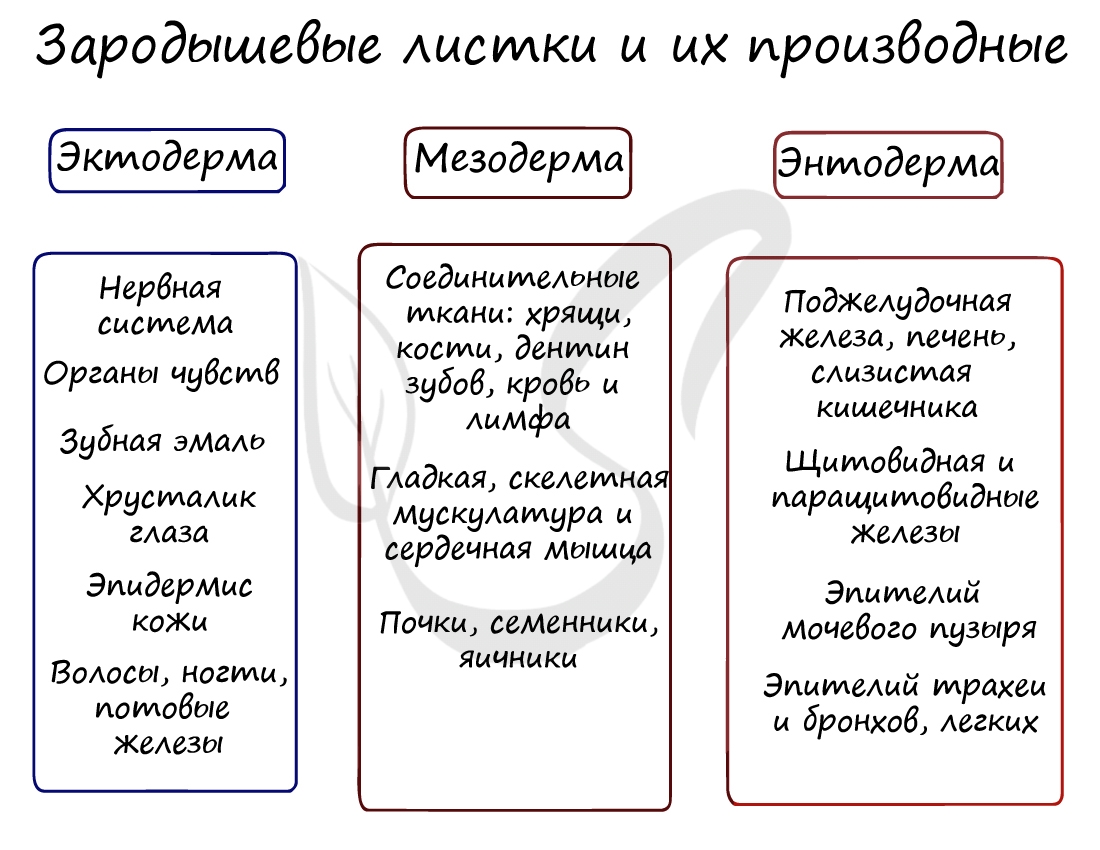
Из эктодермы у позвоночных животных образуется нервная система, органы чувств, покровный эпителий с его железами и производными структурами (волосы, перья, копыта, когти и т. п.).
Из энтодермы формируются органы пищеварительной и дыхательной системы: эпителий средней кишки, печень и поджелудочная железа, жабры, лёгкие, плавательный пузырь, а также щитовидная железа.
Из мезодермы формируются все виды мышечной и соединительной ткани (например, дерма кожи, тела позвонков), кровеносная система, органы выделения, половые железы.
Эмбриональное развитие животных - процесс, протекающий от момента образования зиготы до выхода из яйцевых оболочек или рождения.
Вопрос 2. Назовите стадии эмбрионального развития многоклеточных животных.
Основные этапы эмбрионального периода развития

Вопрос 3. Охарактеризуйте период дробления.
Дробление – процесс деления, в результате которого образуется бластула.
Характер дробления яйцеклеток зависит от количества желтка, находящегося в яйцеклетке. Желток, будучи инертным, не играет активной роли в дроблении, которое осуществляется ядром и цитоплазмой клетки, он проявляет местное замедляющее действие путем механического торможения этого процесса.
Различают следующие типы дробления: полное – голобластическое, когда вся цитоплазма зиготы подвергается дроблению и меробластическое или неполное, когда цитоплазма только в анимальном полюсе подвергается дроблению – такой тип дробления называют дискоидальным. По времени дробления различают равномерное и неравномерное. Дробление у животных, имеющих изолецитальные яйца идет по голобластическому типу.
Для анамний характерно дробление неполное, дискоидальное - у птиц и рептилий; полное, равномерное, асинхронное - у млекопитающих.
Вопрос 4. Чем дробление отличается от митотического деления клеток взрослых животных?
Можно выделить ряд отличий дробления от митоза (деления клеток):
1. Дробление характерно лишь для царства Животных. Процесс деления можно наблюдать у представителей всех царств живых организмов, существующих на Земле.
2. Деление – процесс, который идет перед оплодотворением. Процесс дробления запускается сразу после оплодотворения.
3. Дробление связано только с половым размножением. Деление – часть полового или собственно бесполое размножение.
Вопрос 5. Как образуется двуслойный зародыш?
Гаструляция – это процесс образования двухслойного зародыша называется. Для него характерно:
1) перемещение клеточных масс;
2) начало использования наследственной информации клеток зародыша;
3) появление первых признаков дифференцировки клеток;
4) возникновение первых тканей организма - зародышевых листков.
Вопрос 6. Какие зародышевые листки образуются в ходе эмбрионального развития?
В ходе эмбрионального развития образуется эктодерма, энтодерма и мезодерма – зародышевые листки.
ВОПРОСЫ И ЗАДАНИЯ ДЛЯ ОБСУЖДЕНИЯ
Вопрос 1. Что собой представляет и в чём выражается дифференцировка клеток в процессе эмбрионального развития?
Вопрос 2. Что такое эмбриональная индукция? Как можно доказать, что зачаток одного органа влияет на другой и определяет направление его развития?
Эмбриональная индукция — взаимодействие между частями развивающегося организма у многоклеточных, беспозвоночных и всех хордовых.
Явление было открыто в 1901 году при изучении образования зачатка хрусталика глаз у зародышей земноводных.
Вопрос 3. О чём свидетельствует гомология зародышевых листков?
Гомология зародышевых листков подавляющего большинства животных – одно из доказательств единства животного мира.
Регуляция эмбриогенеза осуществляется на всех уровнях биологической организации организма: надклеточном, клеточном и молекулярно-генетическом.
Надклеточный уровень. Большое значение в управлении ходом эмбриогенеза придается организационным центрам (организаторам). Впервые их роль была установлена в 1924 году немецким ученым Г. Шпеманом. Он проводил свои опыты на зародышах тритона. В норме у зародыша тритона из эктодермы на спинной стороне формируется нервная трубка. Однако если на стадии ранней гаструлы удалить верхнюю губу бластопора, то нервная трубка не сформируется. Если верхнюю губу бластопора пересадить под эктодерму брюшной стороны, то нервная трубка сформируется на брюшной стороне. Если добавить зародышу еще одну губу, то сформируется две нервные трубки.Из проведенных опытов следует, что верхняя губа бластопора направляет развитие эктодермы по пути формирования нервной трубки. Участок верхней губы бластопора Шпеман назвал организационным центром, или индуктором, а само явление получило название – эмбриональная индукция. Ткань, отвечающая на действие индуктора, – компетентная ткань. В последующем были установлены многочисленные примеры взаимовлияния зачатков в ходе эмбриогенеза. Причем деление зачатков на индукторы и компетентную ткань является относительным. Так, при закладке глаза вырост мозгового пузыря вызывает развитие из эктодермы зачатка хрусталика, а зачаток хрусталика - развитие зачатка роговицы.Исходя из учения Шпемана, ход эмбриогенеза можно представить как цепочку, состоящую из пар:
индуктор компетентная ткань (индуктор) компетентная ткань и т.д.
Клеточный уровень. В эмбриогенезе наблюдается пять типов клеточных реакций:
1. Пролиферация.
2. Клеточные перемещения.
3. Гибель клеток.
4. Избирательная сортировка.
5. Дифференцировка.
Пролиферация – размножение клеток митозом. Имеет место при формировании любого органа.
Клеточные перемещения – миграция отдельных клеток развивающегося организма. Например, перемещение нервных клеток ганглиозной пластинки к местам закладки рецепторного аппарата органов чувств.
Гибель клеток – запрограммированный процесс на завершающем этапе формообразования органа. Например, гибель клеток в межпальцевых промежутках кисти человека. Если она не произойдет, то ребенок родится со сросшимися пальцами (синдактилия).
Избирательная сортировка – выделение из смеси однотипных клеток и образование между ними прочных контактов.
Дифференцировка клеток – процесс образования специализированных типов клеток. Можно выделить три этапа на пути дифференцировки клеток:
1) тотипотентность (равнонаследственность) – путь развития клетки еще не определен. Это стадия зиготы и начало ее дробления (2-8 бластомеров). У гидромедузы клетки тотипотентны до стадии 32 бластомеров;
2) трансдетерминация – переопределение намеченного пути дифференцировки. Клетка теряет тотипотентность, но способна изменить направление намеченного пути развития (опыты Шпемана);
3) детерминация – клетка имеет строго определенный путь своего развития.
Таким образом, в ходе эмбриогенеза число возможных путей развития каждой клетки уменьшается в конечном счете до одного.
Молекулярно-генетический уровень. Ранние этапы эмбриогенеза (дробление) управляются веществами (РНК, белки), накопленными яйцеклеткой в ходе оогенеза. Они находятся в цитоплазме. Доказательством этого служат опыты английского ученого Д. Гердона, проведенные им в 1962-1972 гг. Он брал яйцеклетку лягушки, удалял из нее ядро и помещал туда ядро специализированной клетки эпителия кишечника. В последующем из такой клетки развивалась нормальная лягушка. Этим опытом было доказано:
1) все специализированные клетки имеют полный набор генов;
2) ранние стадии эмбриогенеза управляются не ядром, а цитоплазмой.
Для объяснения механизмов регуляции эмбриогенеза на молекулярно-генетическом уровне была предложена гипотеза дифференциальной активности генов: в ходе эмбриогенеза наблюдается последовательная смена активности генов, т.е. гены функционируют поочередно. Включение и выключение генов происходит за счет продуктов деятельности самих генов, т.е. путем саморегуляции.

Экспрессия отдельных генов регулируется на уровне транскрипции негистоновыми белками и гормонами. Различают пептидные гормоны (инсулин) и стероидные (эстрогены и андрогены). Молекулы пептидных гормонов из-за крупных размеров не могут проникнуть в клетку, и поэтому их эффект осуществляется через белки-рецепторы, локализованные в мембранах клеток-мишеней. Стероидные гормоны проникают через мембрану и связываются там с рецепторными белками, образуя комплекс: гормон+белок-рецептор. Затем этот комплекс связывается с негистоновыми белками, которые соединены с промоторными районами специфических генов. При этом промотор освобождается для действия РНК-полимеразы и начинается процесс транскрипции.
Доказательства справедливости гипотезы дифференциальной активности генов:
1) в ходе эмбриогенеза (онтогенеза) наблюдается смена локализации пуффов политенных хромосом у двукрылых насекомых. Пуфф - область интенсивного синтеза иРНК;
2) в онтогенезе человека имеет место смена нескольких видов гемоглобинов:
| Стадия | Гемоглобин |
| Эмбрион | Gover I Gover II Portland I |
| Плод | Гемоглобин F |
| Взрослый | Гемоглобин А Гемоглобин А2 |
Процесс дифференцировки сопровождается уменьшением числа активных генов. Например, у морского ежа из 40 тысяч генов функционируют:
- на стадии бластулы – 30 тысяч;
- на стадии гаструлы – 15-20 тысяч;
- у взрослой особи – 3-5 тысяч генов.
Регуляция эмбриогенеза осуществляется на всех уровнях биологической организации организма: надклеточном, клеточном и молекулярно-генетическом.
Надклеточный уровень. Большое значение в управлении ходом эмбриогенеза придается организационным центрам (организаторам). Впервые их роль была установлена в 1924 году немецким ученым Г. Шпеманом. Он проводил свои опыты на зародышах тритона. В норме у зародыша тритона из эктодермы на спинной стороне формируется нервная трубка. Однако если на стадии ранней гаструлы удалить верхнюю губу бластопора, то нервная трубка не сформируется. Если верхнюю губу бластопора пересадить под эктодерму брюшной стороны, то нервная трубка сформируется на брюшной стороне. Если добавить зародышу еще одну губу, то сформируется две нервные трубки.Из проведенных опытов следует, что верхняя губа бластопора направляет развитие эктодермы по пути формирования нервной трубки. Участок верхней губы бластопора Шпеман назвал организационным центром, или индуктором, а само явление получило название – эмбриональная индукция. Ткань, отвечающая на действие индуктора, – компетентная ткань. В последующем были установлены многочисленные примеры взаимовлияния зачатков в ходе эмбриогенеза. Причем деление зачатков на индукторы и компетентную ткань является относительным. Так, при закладке глаза вырост мозгового пузыря вызывает развитие из эктодермы зачатка хрусталика, а зачаток хрусталика - развитие зачатка роговицы.Исходя из учения Шпемана, ход эмбриогенеза можно представить как цепочку, состоящую из пар:
индуктор компетентная ткань (индуктор) компетентная ткань и т.д.
Клеточный уровень. В эмбриогенезе наблюдается пять типов клеточных реакций:
1. Пролиферация.
2. Клеточные перемещения.
3. Гибель клеток.
4. Избирательная сортировка.
5. Дифференцировка.
Пролиферация – размножение клеток митозом. Имеет место при формировании любого органа.
Клеточные перемещения – миграция отдельных клеток развивающегося организма. Например, перемещение нервных клеток ганглиозной пластинки к местам закладки рецепторного аппарата органов чувств.
Гибель клеток – запрограммированный процесс на завершающем этапе формообразования органа. Например, гибель клеток в межпальцевых промежутках кисти человека. Если она не произойдет, то ребенок родится со сросшимися пальцами (синдактилия).
Избирательная сортировка – выделение из смеси однотипных клеток и образование между ними прочных контактов.
Дифференцировка клеток – процесс образования специализированных типов клеток. Можно выделить три этапа на пути дифференцировки клеток:
1) тотипотентность (равнонаследственность) – путь развития клетки еще не определен. Это стадия зиготы и начало ее дробления (2-8 бластомеров). У гидромедузы клетки тотипотентны до стадии 32 бластомеров;
2) трансдетерминация – переопределение намеченного пути дифференцировки. Клетка теряет тотипотентность, но способна изменить направление намеченного пути развития (опыты Шпемана);
3) детерминация – клетка имеет строго определенный путь своего развития.
Таким образом, в ходе эмбриогенеза число возможных путей развития каждой клетки уменьшается в конечном счете до одного.
Молекулярно-генетический уровень. Ранние этапы эмбриогенеза (дробление) управляются веществами (РНК, белки), накопленными яйцеклеткой в ходе оогенеза. Они находятся в цитоплазме. Доказательством этого служат опыты английского ученого Д. Гердона, проведенные им в 1962-1972 гг. Он брал яйцеклетку лягушки, удалял из нее ядро и помещал туда ядро специализированной клетки эпителия кишечника. В последующем из такой клетки развивалась нормальная лягушка. Этим опытом было доказано:
1) все специализированные клетки имеют полный набор генов;
2) ранние стадии эмбриогенеза управляются не ядром, а цитоплазмой.
Для объяснения механизмов регуляции эмбриогенеза на молекулярно-генетическом уровне была предложена гипотеза дифференциальной активности генов: в ходе эмбриогенеза наблюдается последовательная смена активности генов, т.е. гены функционируют поочередно. Включение и выключение генов происходит за счет продуктов деятельности самих генов, т.е. путем саморегуляции.
Экспрессия отдельных генов регулируется на уровне транскрипции негистоновыми белками и гормонами. Различают пептидные гормоны (инсулин) и стероидные (эстрогены и андрогены). Молекулы пептидных гормонов из-за крупных размеров не могут проникнуть в клетку, и поэтому их эффект осуществляется через белки-рецепторы, локализованные в мембранах клеток-мишеней. Стероидные гормоны проникают через мембрану и связываются там с рецепторными белками, образуя комплекс: гормон+белок-рецептор. Затем этот комплекс связывается с негистоновыми белками, которые соединены с промоторными районами специфических генов. При этом промотор освобождается для действия РНК-полимеразы и начинается процесс транскрипции.
Доказательства справедливости гипотезы дифференциальной активности генов:
1) в ходе эмбриогенеза (онтогенеза) наблюдается смена локализации пуффов политенных хромосом у двукрылых насекомых. Пуфф - область интенсивного синтеза иРНК;
2) в онтогенезе человека имеет место смена нескольких видов гемоглобинов:
| Стадия | Гемоглобин |
| Эмбрион | Gover I Gover II Portland I |
| Плод | Гемоглобин F |
| Взрослый | Гемоглобин А Гемоглобин А2 |
Процесс дифференцировки сопровождается уменьшением числа активных генов. Например, у морского ежа из 40 тысяч генов функционируют:
От момента образования зиготы и до выхода зародыша из яйцевых оболочек длится эмбриональный период развития.
Дробление зиготы
После того, как произошло оплодотворение - слияние сперматозоида и яйцеклетки, образовавшаяся зигота начинает интенсивно делиться. Ее множественные митотические деления называют дроблением.
Важная особенность дробления в том, что не происходит увеличение в размере зародыша: клетки дробятся (делятся) настолько быстро, что не успевают накопить цитоплазматическую массу. Дробление зиготы человека является полным неравномерным асинхронным.
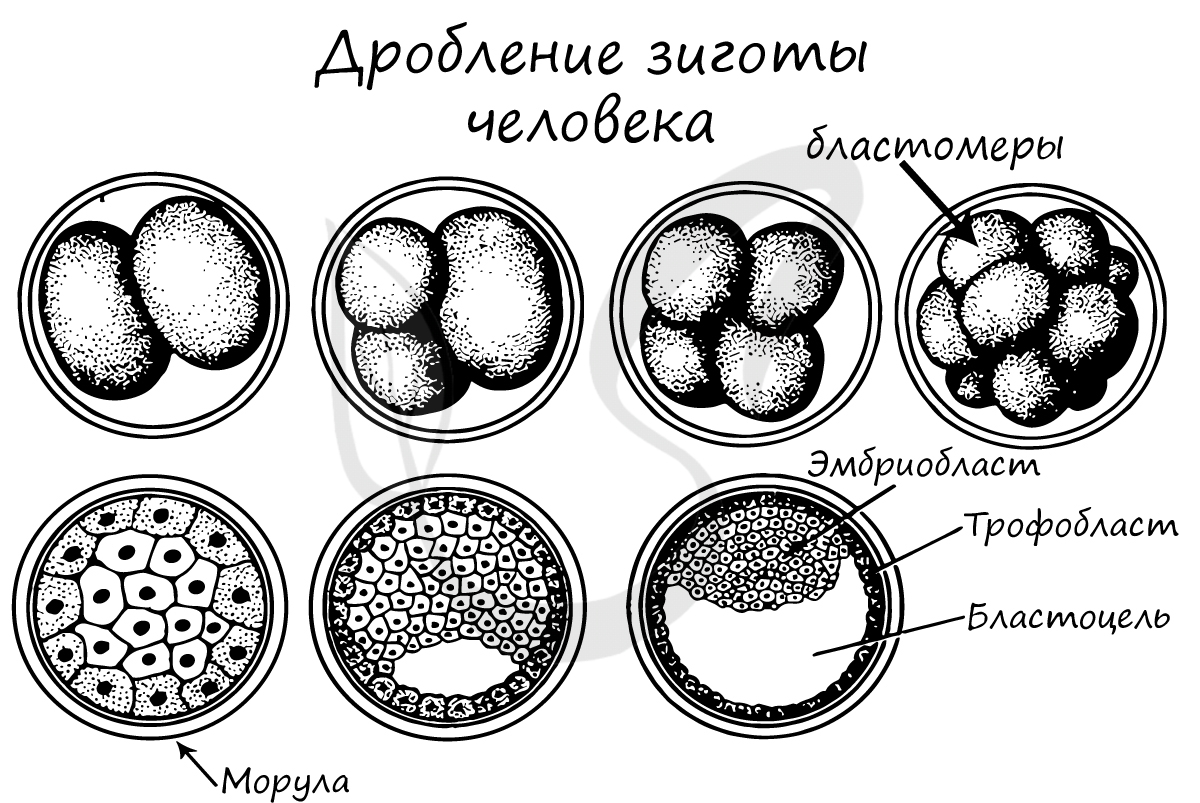
В результате дробления образуется морула. Морула (лат. morum - ягода тутового дерева) - клетка на стадии этапа дробления, когда зародыш представляет собой компактную совокупность клеток (без полости внутри).
Бластуляция
Бластуляция - заключительный период дробления, в который зародыш называется бластулой.
После очередных этапов многократного деления образуется однослойный зародыш с полостью внутри - бластула (греч. blastos — зачаток).
Стенки бластулы состоят из бластомеров, которые окружают центральную полость - бластоцель (греч. koilos — полый). Соединяясь друг с другом, бластомеры образуют бластодерму из одного слоя клеток.

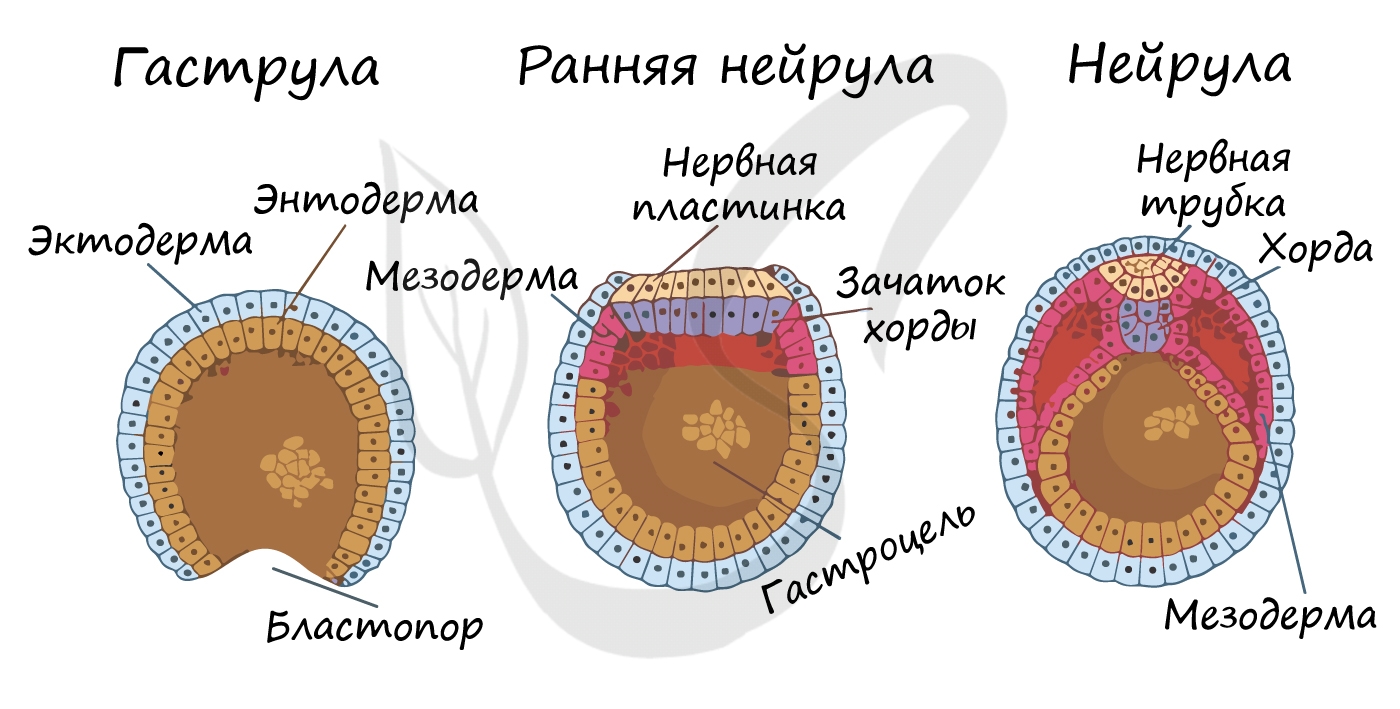
Гаструляция (греч. gaster — желудок, чрево)
Гаструляцией называют стадию эмбрионального развития, в ходе которой клетки, возникшие в результате дробления зиготы, формируют три зародышевых листка: эктодерму, мезодерму и энтодерму.
Стенка бластулы начинается впячиваться внутрь - происходит инвагинация стенки. По итогу такого впячивания зародыш становится двухслойным. Двухслойный зародыш называется - гаструла. Полость гаструлы называется гастроцель (полость первичной кишки), а отверстие, соединяющее гастроцель и внешнюю среду - первичный рот (бластопор).

У первичноротых животных на месте первичного рта (бластопора) образуется ротовое отверстие. К первичноротым относятся: кишечнополостные, плоские, круглые и кольчатые черви, моллюски, членистоногие.
У вторичноротых на месте бластопора формируется анальное отверстие, а ротовое отверстие образуется на противоположном полюсе. К вторичноротым относят хордовых и иглокожих (морских звезд, морских ежей).

При впячивании части бластулы (инвагинации) клетки бластодермы мигрируют внутрь и становятся энтодермой (греч. entós — внутренний). Оставшаяся часть бластодермы снаружи называется эктодермой (греч. ἔκτος - наружный).
Между энто- и эктодермой из группы клеток формируется третий зародышевый листок - мезодерма (греч. μέσος — средний).

Нейрула
Эта стадия следует за гаструлой. Ранняя нейрула представляет собой трехслойный зародыш, состоящий из энто-, экто- и мезодермы. На этапе нейрулы происходит закладка отдельных органов.
Важно отметить, что на стадии нейрулы происходит процесс нейруляции - закладывание нервной трубки. Нервная пластинка, образовавшаяся на ранних этапах, прогибается внутрь, при этом ее края сближаются и, замыкаясь, формируют нервную трубку.

Итак, как уже было сказано, на стадии нейрулы закладываются отдельные органы. Эктодерма образует покровный эпителий и нервную пластинку, мезодерма (из которой в дальнейшем появятся все соединительные ткани), энтодерма - окружает полость первичной кишки (гастроцель), образуя кишечник. От энтодермы отшнуровывается хорда.
Все три зародышевых листка требуют нашего особого внимания, а также понимания того, какие органы и структуры из них образуются.
Эктодерма (греч. ἔκτος - наружный) - наружный зародышевый листок, образует головной и спинной мозг, органы чувств, периферические нервы, эпителий кожи, эмаль зубов, эпителий ротовой полости, эпителий промежуточного и анального отделов прямой кишки, гипофиз, гипоталамус.
Мезодерма (греч. μέσος — средний) - средний зародышевый листок, образует соединительные ткани: кровеносную и лимфатическую системы, костную и хрящевую ткань, мышечные ткани, дентин и цемент зубов, а также выделительную (почки) и половую системы (семенники, яичники).
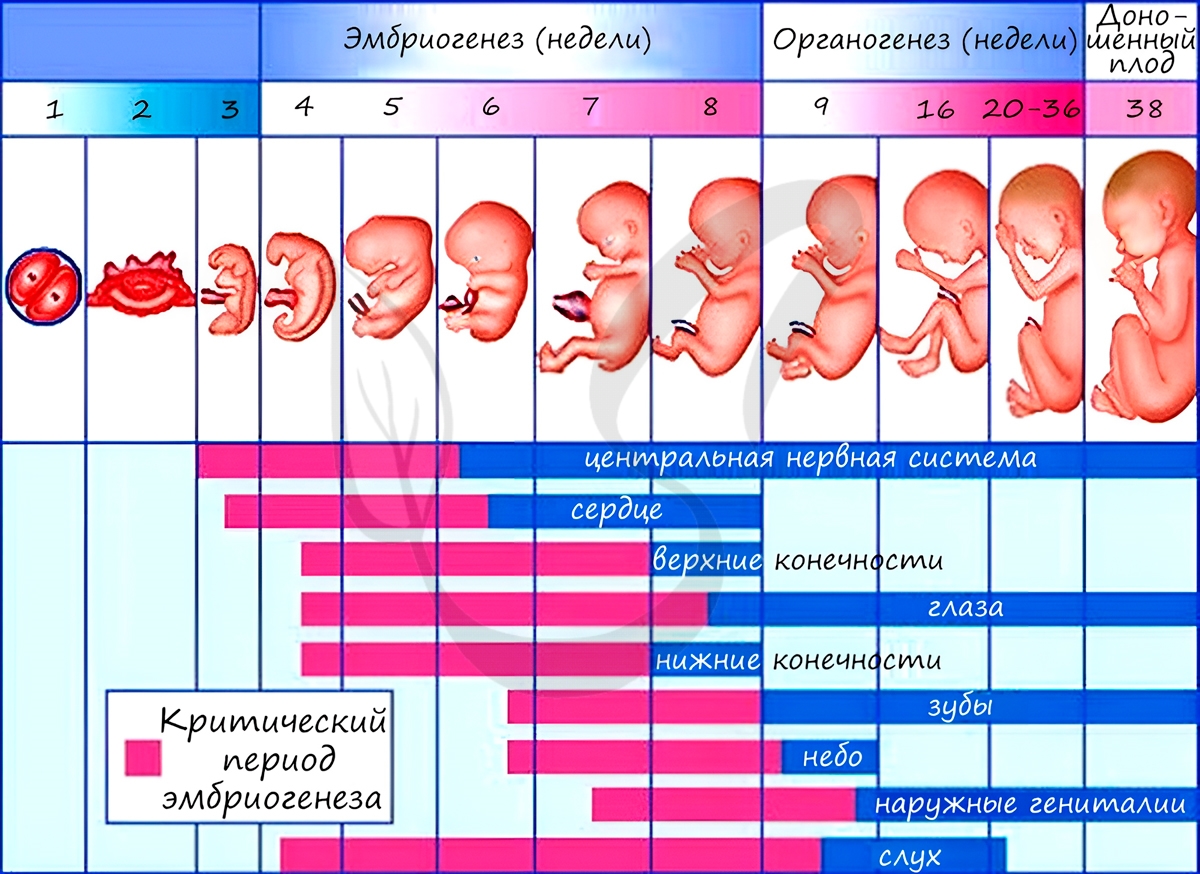
Из зародышевых листков образуются ткани, органы и системы органов. Такой процесс называется органогенезом. В период закладки органов важное значение имеет воздержание матери от вредных привычек (алкоголь, курение), которые могут нарушить процесс дифференцировки клеток и привести к тяжелейшим аномалиям, уродствам плода.
Некоторые лекарства также могут оказывать на плод тератогенный эффект (греч. τέρας — чудовище, урод), приводя к развитию уродств. Периоды закладки органов и система органов вследствие их большой важности носят название критических периодов эмбриогенеза.
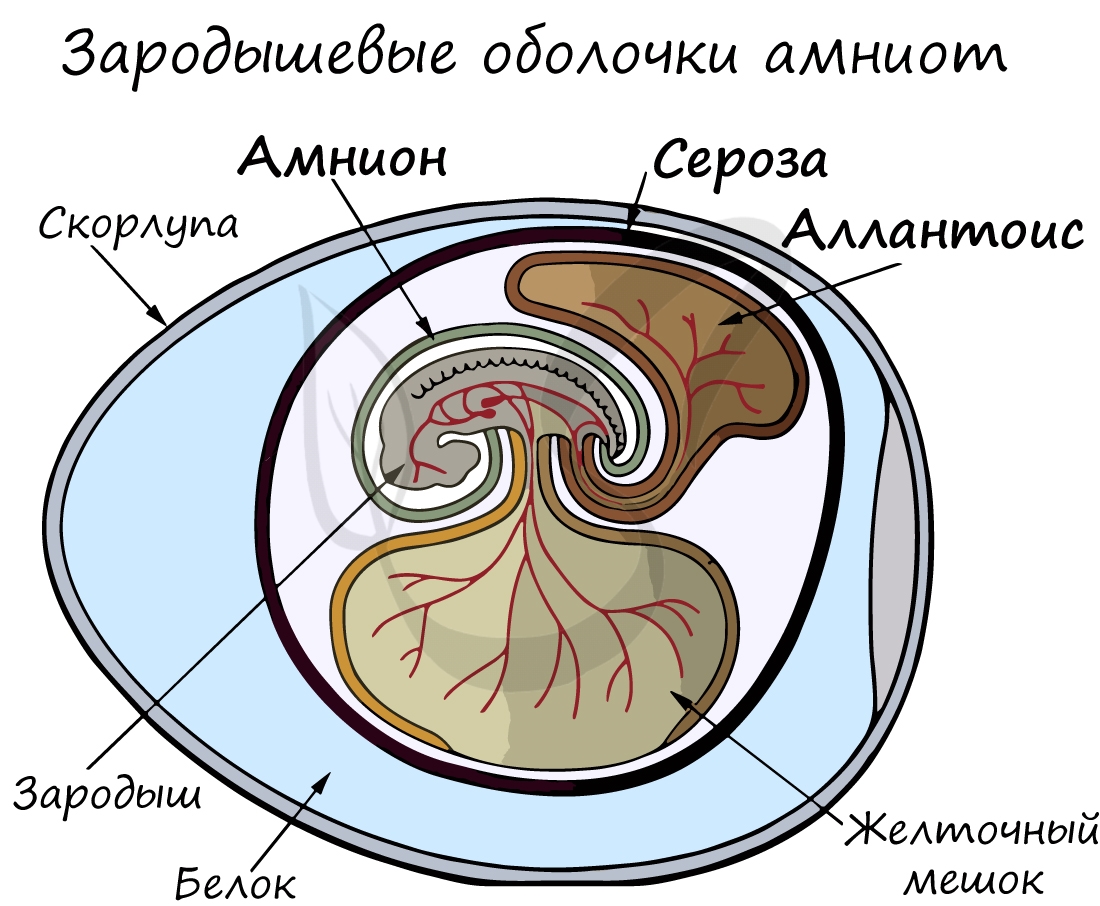
Анамнии и амниоты
Анамнии, или низшие позвоночные - группа животных, не имеющая зародышевых оболочек (зародышевого органа - аллантоиса и амниона). Анамнии проводят большую часть жизни в воде, без которой невозможно их размножение.
К анамниям относятся рыбы, земноводные.

Амниоты - группа высших позвоночных, характеризующаяся наличием зародышевых оболочек. К амниотам относятся пресмыкающиеся, птицы и млекопитающие.
Зародышевый орган, аллантоис, является органом дыхания и выделения.
За счет особых оболочек, развивающихся в ходе эмбрионального развития, амниона и серозы, у амниот формируется амниотическая полость. В ней находится зародыш, окруженный околоплодными водами. Благодаря такому гениальному устройству, амниотам для размножения и развития более не нужно постоянное нахождение в водоеме, они "обрели независимость" от него.
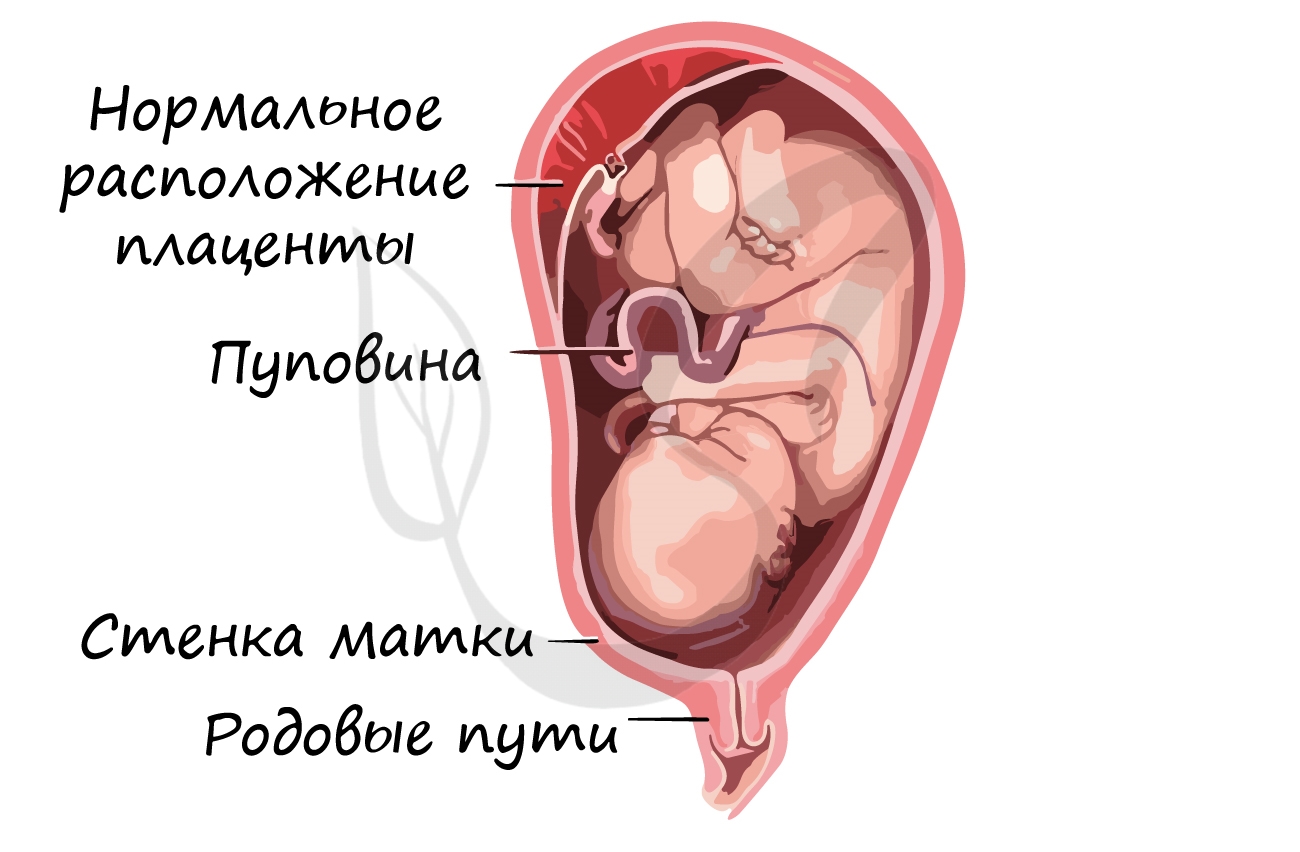
Развитие плода происходит в мышечном органе - матке, которая, сокращаясь во время родов, стимулирует изгнание плода через родовые пути. Питание осуществляется через плаценту - "детское место" - орган, который с одной стороны омывается кровью матери, а с другой - кровью плода. Через плаценту происходит транспорт питательных веществ и газообмен.
Соединяет плаценту и плод особый орган - пуповина, внутри которой проходят артерии, вены.
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Читайте также: