
Принцип общего конечного пути кратко
Обновлено: 19.05.2024
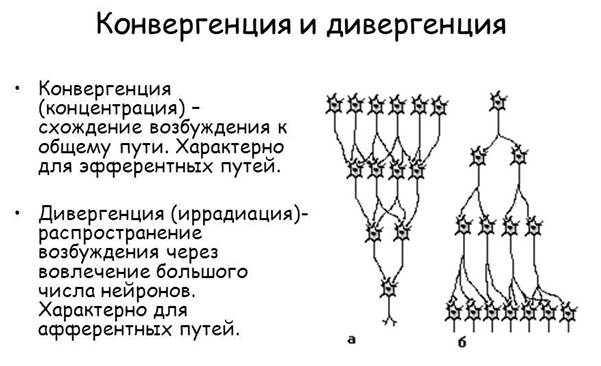
Принцип конвергенции возбуждения (или принцип общего конечного пути, воронка Шеррингтона). Конвергенция нервных импульсов означает схождение к одному нейрону двух или нескольких различных возбуждений одновременно.
Это явление было открыто Ч. Шеррингтоном. Он показал, что одно и то же движение, например рефлекторное сгибание конечности в коленном суставе, можно вызвать путем раздражения различных рефлексогенных зон. В связи с этим им было введено понятие "общего конечного пути", или "принципа воронки", согласно которому потоки импульсов от различных нейронов могут сходиться на одном и том же нейроне (в данном случае - на альфа-мотонейронах спинного мозга). В частности, Ч. Шеррингтон обнаружил схождение к одним и тем же промежуточным или эфферентным нейронам различных афферентов от разных участков общего рецептивного поля (в спинном и продолговатом мозге) или даже от разных рецептивных полей (в высших отделах головного мозга). В настоящее время показано, что конвергенция возбуждения, так же как и дивергенция возбуждения, - очень распространенное явление в ЦНС.
Основой для конвергенции (как и для иррадиации) является определенная морфологическая и функциональная структура различных отделов мозга. Очевидно, что часть конвергентных путей является врожденными, а другая часть (главным образом в коре большого мозга) - приобретенная в результате обучения в процессе онтогенеза. Формирование новых конвергентных отношений для нейронов коры большого мозга в процессе онтогенеза во многом связаны с формированием в коре доминантных очагов возбуждения, которые способны "притягивать" к себе возбуждение от других нейронов.
30. Центры вегетативной нервной системы
Мезенцефалический – волокна входят в состав глазодвигательного нерва (парасимпатический)
Бульбарный – волокна в составе лицевого, языкоглоточного и блуждающего нервов (парасимпатический)
Тораколюмбальный – ядра боговых рогов с 8 шейного по 3 поясничный сегменты (симпатический)
Сакральный – во 2-4-м сегментах крестцового отдела спинного мозга (парасимпатический)
31. Отделы вегетативной нервной системы
Симпатический отдел. Тела первых нейронов симпатического отдела ВНС расположены преимущественно в задних ядрах гипоталамуса, среднем и продолговатом мозге и в передних рогах спинного мозга, начиная с 1-го грудного и кончая 3, 4-м сегментом поясничного ее отдела.
Парасимпатический отдел. Центральные нейроны парасимпатического отдела вегетативной нервной системы расположены преимущественно в передних отделах гипоталамуса, среднем и продолговатом мозге, во 2-4-м сегментах крестцового отдела спинного мозга.
Симпатическая нервная система активируется при стрессовых реакциях. Для неё характерно генерализованное влияние, при этом симпатические волокна иннервируют подавляющее большинство органов.
Известно, что парасимпатическая стимуляция одних органов оказывает тормозное действие, а других — возбуждающее действие. В большинстве случаев действие парасимпатической и симпатической систем противоположно.
Преобладание на конечных путях той или иной рефлекторной реакции обусловлено ее значением для жизнедеятельности организма в данный момент.
В таком отборе важную роль играет наличие в центральной нервной системе доминанты. Она обеспечивает протекание главной реакции. Например, ритмический шагательный рефлекс и одиночный, непрерывный рефлекс сгибания при болевом раздражении являются антагонистическими. Однако спортсмен, внезапно получивший травму, может продолжать бег к финишу, т. е. осуществлять ритмический рефлекс и подавлять болевые раздражения, которые, поступая к мотонейронам сгибательных мышц, препятствуют попеременному сгибанию и разгибанию ноги.

1. Иррадиация и концентрация нервных процессов:
2. Торможение в ЦНС:

3. Принцип общего конечного пути:

4. Принцип проторения пути


Преобладание на конечных путях той или иной рефлекторной реакции обусловлено ее значением для жизнедеятельности организма в данный момент.
В таком отборе важную роль играет наличие в центральной нервной системе доминанты. Она обеспечивает протекание главной реакции. Например, ритмический шагательный рефлекс и одиночный, непрерывный рефлекс сгибания при болевом раздражении являются антагонистическими. Однако спортсмен, внезапно получивший травму, может продолжать бег к финишу, т. е. осуществлять ритмический рефлекс и подавлять болевые раздражения, которые, поступая к мотонейронам сгибательных мышц, препятствуют попеременному сгибанию и разгибанию ноги.
1. Иррадиация и концентрация нервных процессов:
2. Торможение в ЦНС:
3. Принцип общего конечного пути:
4. Принцип проторения пути
Суть этого механизма была раскрыта английским физиологом Ч. Шеррингтоном, который сформулировал принцип общего конечного пути. По Ч. Шеррингтону, количественное преобладание чувствительных и других приходящих волокон над двигательными создает неизбежное столкновение импульсов в общем конечном пути, которым являются группа мотонейронов и иннервируемые ими мышцы. В результате этого столкновения достигается торможение всех возможных степеней свободы двигательного аппарата, кроме одной, в направлении которой протекает рефлекторная реакция, вызванная максимальной стимуляцией одного из афферентных входов.
Рассмотрим случай с одновременным раздражением рецептивных полей чесательного и сгибательного рефлексов, которые реализуются идентичными группами мышц. Импульсы, идущие от этих рецептивных полей, приходят к одной и той же группе мотонейронов, и здесь в узком месте воронки за счет интеграции синаптических влияний осуществляется выбор в пользу сгибательного рефлекса, вызванного более сильным болевым раздражением. Принцип общего конечного пути, как один из принципов координации, действителен не только для спинного мозга, он применим для любого этажа ЦНС, в том числе для моторной коры.
Вопрос 56
Вопрос 57
Какое значение ретикулярной формации в восприятии информации?
Человек познает мир с помощью информации (сигналов), которую получает, перерабатывает и с помощью которой принимает решения и формирует поведение. Восприятие информации связано с ретикулярной формацией.
Ретикулярная формация и кора больших полушарий головного мозга тесно связаны между собой. Между ними действует связь: кора-ретикулярная формация-кора.
Вопрос 58
Вопрос 59

Компенсаторные реакции возникают в ответ на нарушение функций, структур, обменных процессов и являются реакцией целостного организма. Они направлены, прежде всего, на восстановление гармоничных, координированных взаимоотношений органов и систем
в интересах целостного организма; поддерживают и сохраняют равновесие организма со средой. В основе сложного механизма компенсации лежит перестройка функций организма, регулируемая центральной нервной системой (ЦНС).
Чем тяжелее дефект, тем большее количество систем организма включается в процесс компенсации. Наиболее сложные функциональные перестройки наблюдаются при нарушениях ЦНС, в том числе и анализаторов. Таким образом, степень сложности механизмов компенсаторных явлений находится в зависимости от тяжести дефекта.
Автоматизм включения компенсаторных функций не определяет сразу механизмы компенсации; так, при сложных нарушениях деятельности организма они формируются постепенно. Постепенность развития компенсаторных процессов проявляется в том, что они имеют определенные стадии становления, которые характеризуются особым составом и структурой динамических систем нервных связей и своеобразием протекания процессов возбуждения и торможения.
Материальным субстратом компенсаторных перестроек является центральная нервная система. Формирование механизмов компенсации подчинено законам высшей нервной деятельности. Современная теория компенсации рассматривает компенсаторные явления в свете рефлекторной теории И.П. Павлова. Эта теория, базирующаяся на трех основных принципах: причинность (детерминизм), единство анализа и синтеза и структурность.
Применительно к учению о компенсации нарушенных или утраченных функций принципы рефлекторной деятельности означают следующее:
Принцип причинности. Любой дефект неизбежно вызывает ответную реакцию организма, причем сила и характер этой реакции зависят не только от степени нарушения той или иной функции или органа, но и от состояния организма и тех условий, которые его окружают. Эта реакция имеет своим механизмом замыкание новых временных связей в коре больших полушарий головного мозга.
Так, известен случай, когда после четырехкратной операции по поводу опухоли мозга у 12-летнего ребенка была удалена большая часть левого полушария мозжечка. Сразу после каждой операции у ребенка возникали нарушения двигательной сферы, речи и других функций мозга. Однако довольно быстро эти нарушения компенсировались. Компенсаторные возможности мозга с возрастом уменьшаются, это обусловлено ослаблением лабильности в формировании новых функциональных связей.
Принцип единства анализа и синтеза. В процессе анализа и синтеза, внешних воздействии у человека образуется весьма сложная, по строению функциональная система анализаторов. Полное или частичное нарушение функций какого-либо анализатора приводит к определенным нарушениям этой системы, что отражается в первую очередь на аналитической деятельности. Включение компенсаторных функций приводит к перестройке сохранных анализаторов, благодаря чему способность к аналитико-синтетической деятельности сохраняется, хотя диапазон, уровень, степень и путь анализа суживаются.
В результате взамен утраченного способа образования временных нервных связей в нервной системе проторяются новые, обходные пути, формируются новые условно-рефлекторные нервные связи, восстанавливающие нарушившееся равновесие во взаимоотношениях организма и среды.
Таким образом, физиологический механизм компенсации основывается на нормальном функционировании сохранных систем. При этом включение механизмов компенсации происходит, безусловно-рефлекторным путем, автоматически, а дальнейшее развитие компенсаторных приспособлений есть деятельность условно рефлекторная.
Свойства ЦНС, обеспечивающие механизмы компенсации:
Полифункциональность и полисенсорность каждого из элементов нервной системы. Основная функция нервной системы заключается в сборе, переработке, хранении, воспроизведении и передаче информации с целью организации интеллектуальной, поведенческой деятельности, регуляции функционирования органов, систем органов и обеспечения их взаимодействия. Многие из перечисленных функций реализуются уже на нейронном уровне. Нейроны обладают способностью выполнять все информационные функции нервной системы: восприятие, обработку, хранение, многократное воспроизведение и передачу информации. В этом и заключается основной принцип функционирования нервной системы – принцип полифункциональности.
Полифункциональность присуща большинству структур ЦНС. Например, сенсомоторная кора способна воспринимать сигналы кожной, зрительной, слуховой и других видов рецепции. В ответ на эти сигналы в сенсомоторной коре формируются реакции, которые обычно возникают при нормальной деятельности коркового конца зрительного, слухового или других анализаторов. Следовательно, благодаря полифункциональности одна и та же функция может быть выполнена разными структурами мозга. Этот принципиальный момент свидетельствует о практически безграничных возможностях компенсации функции в ЦНС.
Свойства полифункциональности нервных центров тесно связаны со свойством полисенсорности нейронов. Полисенсорность – это способность одного нейрона реагировать на сигналы разных афферентных систем. Нейрофизиологи выделяют нейроны моносенсорные, реагирующие только на один вид сигналов, бисенсорные – реагирующие на два разных сигнала, например, некоторые нейроны зрительной коры могут реагировать на зрительные и слуховые раздражения. Наконец, в коре мозга имеются нейроны, которые реагируют на три и более вида сигналов. Эти нейроны называются полисенсорными.
Относительная специализация нейронов отдельных областей мозга и локализация функций в коре. Нейроны отдельных областей мозга способны реагировать только на одну характеристику сенсорного раздражения, например, на определенную частоту звука или только на один цвет. Такие нейроны называются мономодальными (моносенсорными). Они обладают высокой избирательностью и высокой чувствительностью к определенным видам раздражений, т.е. являются специализированными. Локализуются специализированные нейроны в зонах первичных проекций анализаторов. Такими зонами являются первичные области зрительной, слуховой, кожной и других зон коры.
Локализация функций в коре определяется, прежде всего, моносенсорными нейронами, имеющими наименьшие пороги чувствительности на свои адекватные раздражения. Однако рядом с этими нейронами всегда имеются полисенсорные нейроны, которые обеспечивают взаимодействие локальной структуры с другими структурами мозга, а тем самым – возможность образования временной связи, компенсацию нарушений функций своей структуры и структур, с нею связанных. Полимодальные нейроны обеспечивают внутрисистемную компенсацию нарушенных функций.
Параллельная (одновременная) обработка разно сенсорной информации. В коре мозга нет такой зоны, которая была бы связана с реализацией только одной функции. В разных отделах мозга имеется разное количество полисенсорных и полимодальных нейронов. Наибольшее количество таких нейронов находится в ассоциативных и во вторичных, третичных зонах коркового конца анализаторов. Значительная часть нейронов моторной коры (около 40 %) также является полисенсорной, они реагируют на раздражения кожи, на звук, свет.
Число полисенсорных нейронов в структурах мозга меняется в зависимости от функционального состояния нервной системы и от выполняемой в данный момент времени задачи. Так, в период обучения с участием зрительного и моторного анализаторов число полисенсорных нейронов в этих зонах коры возрастает. Следовательно, направленное обучение создает условия увеличения полисенсорных нейронов и, тем самым, компенсаторные возможности нервной системы возрастают.
Важно также, что некоторые нейроны коры мозга в результате обучения способны становиться полисенсорными, т.е. если до применения сочетания условного и безусловного стимулов нейрон реагировал только на безусловный стимул, то после ряда сочетаний этот нейрон становится способным реагировать и на условный стимул.
Полимодальность и полисенсорность позволяют нейрону одновременно воспринимать раздражения от разных анализаторов или, если от одного анализатора, то воспринимать одновременно сигналы с разными его характеристиками.
Структурная избыточность и функциональная надежность. Полифункциональность и полисенсорность связаны с другим свойством функционирования мозга – его надежностью. Надежность также обеспечивается такими механизмами, как избыточность, модульность, кооперативность.
Избыточность достигается разными способами. Наиболее распространенным является резервирование элементов. У человека в коре постоянно активны только доли процента нейронов, но их достаточно для поддержания тонуса коры, необходимого для реализации ее деятельности. При нарушении функционирования коры количество фоновоактивных нейронов в ней значительно увеличивается. Избыточность элементов в ЦНС обеспечивает сохранение функций ее структур даже при повреждении значительной их части.
Например, удаление значительной части зрительной коры не приводит к нарушениям зрения. Одно полушарное повреждение структур лимбической системы не вызывает специфических для нее клинических симптомов. Доказательством того, что нервная система имеет большие резервы, являются следующие примеры. Глазодвигательный нерв нормально реализует свои функции регуляции движений глазного яблока при сохранности в его ядре всего 45 % нейронов. Отводящий нерв нормально иннервирует свою мышцу при сохранности 38 % нейронов его ядра, а лицевой нерв выполняет свои функции всего при 10 %-ной сохранности числа нейронов, расположенных в ядре этого нерва.
Высокая надежность в нервной системе обусловлена также множеством связей ее структур, большим количеством синапсов на нейронах. Так, нейроны мозжечка имеют на своем теле и дендритах до 60 тыс. синапсов, пирамидные нейроны двигательной коры – до 10 тыс., альфа-мотонейроны спинного мозга – до 6 тыс. синапсов.
Резервирование проявляется множеством путей реализации сигнала; так, дублирующийся двигательный сигнал, идущий из коры к мотонейронам спинного мозга, может достигнуть их не только от пирамидных нейронов 4 поля коры, но и от добавочной моторной зоны, из других проекционных полей, из базальных ганглиев, красного ядра, ретикулярной формации и других структур. Следовательно, повреждение моторной коры не должно приводить к полному выпадению двигательной информации к мотонейронам спинного мозга.
Следовательно, помимо резервирования, надежность нервной системы достигается дублированием, что позволяет оперативно вводить, по мере надобности, дополнительные элементы, чтобы реализовать ту или иную функцию. Примером такого дублирования может служить многоканальная передача информации, например, в зрительном анализаторе.
Модульность – это принцип структурно-функциональной организации коры мозга, который заключается в том, что в одном нейронном модуле осуществляется локальная переработка информации от рецепторов одной модальности. Между дендритами этого пучка имеют место не только синаптические связи, но и электротонические контакты. Последние обеспечивают синхронность работы нейронов микромодуля, что повышает надежность передачи информации.
В зрительной коре имеет место чередование колонок, нейроны которых реагируют на зрительные стимулы либо только правого, либо только левого глаза. Следовательно, в зрительной коре обоих полушарий мозга имеются глазодоминантные колонки, т.е. колонки, реагирующие на стимуляцию одного глаза (А.Г. Литвак, 2017).
В слуховой коре выделяются колонки, способные дифференцировать сигналы, идущие от обоих ушей, и колонки, не способные к такой дифференциации.
В сенсомоторной коре рядом расположенные колонки выполняют разнонаправленные реакции: например, одни из них возбуждают мотонейроны спинного мозга, другие – тормозят их.
Модульный принцип структурно-функциональной организации работы мозга является проявлением кооперативного характера функционирования нейронов мозга.
Кооперативность создает возможность относительной взаимозаменяемости нейронов, и, тем самым, повышает надежность нервной деятельности. В результате функционирование системы становится мало зависящим от состояния отдельной нервной клетки.
Кооперативность дает возможность структуре выполнять функции, не присущие отдельным ее элементам. Так, отдельный нейрон мозга не способен к обучению, но, находясь в сети нейронов, он приобретает такую способность.
Способность к саморегуляции и самоорганизации. Саморегуляция – свойство структур нервной системы автоматически устанавливать и поддерживать на определенном уровне свое функционирование. Основным механизмом саморегуляции является механизм обратной связи. Обратная связь упорядочивает, суживает множество вариантов прохождения сигнала, создавая тормозное окружение пути возбуждения из неактивных нейронов.
Тесно связан с саморегуляцией нервной системы механизм ее самоорганизации. Самоорганизующиеся системы вообще имеют ряд особенностей, которые присущи и ЦНС: множество входов и выходов; высокий уровень сложности взаимодействия своих элементов; большое количество функционирующих элементов и т.д. Благодаря принципу самоорганизации компенсация функций в нервной системе обеспечивается формированием новых связей на основе включения в активность потенциальных синапсов, использованием накопленного опыта данного индивида.
Развитие нервной системы в онтогенезе приводит к непрерывному усложнению взаимодействия ее систем. Чем больше форм, видов, число условных рефлексов, организуемых в онтогенезе, тем больше связей устанавливается между структурами нервной системы.
Увеличение количества функциональных связей между структурами нервной системы имеет решающее значение, так как в этом случае возрастает число вариантов прохождения сигналов, значительно расширяются возможности компенсации нарушенных функции.
В функции самоорганизации немаловажно то, что нервная система, помимо возможности большого выбора путей для достижения цели, способна избирательно усиливать или ослаблять сигналы.
Так, при усилении сигнала, обеспечивается надежная передача информации при частичной морфологической сохранности структуры, а при ослаблении сигнала – появляется возможность снизить помеху, идущую от других источников. Так как нервная система способна к избирательной фильтрации нужного сигнала, то это позволяет ей, выделив нужный, но слабый сигнал, во-первых, прямо усилить его, а во-вторых, дать ему преимущество при прохождении к воспринимающей структуре за счет снижения силы ненужных, мешающих сигналов.
Принципы иерархичности, иррадиации и концентрация активности. Структурная локализация функций предполагает, что мозг имеет детерминированные пути, системы, реализующие проведение сигнала, организацию той или иной реакции и т.д. Однако помимо жестко детерминированных связей в мозгу реализуются функциональные связи, развивающиеся в онтогенезе. Чем более упрочены, закреплены связи между структурами мозга в процессе индивидуального развития, тем труднее использование компенсаторных возможностей при патологиях.
На основе принципа структурности реализуется механизм иерархичности. Он заключается не столько в соподчинении, сколько в организации компенсаторных процессов. Каждая вышележащая структура участвует в реализации функций нижележащей, но делает это тогда, когда нижележащая структура затрудняется в выполнении своих функций.
Структуры мозга при обучении, при дисфункции одной из них не локализуют возбуждение в своих границах, а позволяют ему широко распространяться по мозгу – принцип иррадиации.
Иррадиация состояния активности распространяется в другие структуры мозга как по прямым связям, так и по опосредованным путям. Возникновение иррадиации при гипофункции структуры, участвующей в реализации того или иного процесса, позволяет найти пути компенсации гипофункции и реализовать нужную реакцию.
Нахождение нового пути закрепляется по рефлекторному принципу и заканчивается концентрацией активности в определенных структурах, заинтересованных в выполнении реакции.
Принцип общего конечного пути. С концентрацией активности в определенных структурах мозга тесно связаны конвергентность и принцип общего конечного пути. Этот принцип реализуется на отдельном нейроне и на системном уровне. В первом случае информация в нейроне собирается на дендритах, соме нейрона, а передается преимущественно через аксон в нейроны других структур мозга. Через синапсы дендритов информация передается только на соседние нейроны.
Наличие общего конечного пути позволяет нервной системе иметь разные варианты достижения нужного эффекта через разные структуры, имеющие выход на один и тот же конечный путь.
Трудности компенсаций, отмечаемые в более старших возрастах, обусловлены не тем, что резервы мозга исчерпаны, а тем, что сформировано большое количество оптимальных путей реализации функций, которые хотя и задействуются в случае патологии, но из-за нее же и не могут быть реализованы. Чаще при патологии требуется формирование новых путей реализации той или иной функции.
Пластичность нервных центров и отдельных нейронов. В основе формирования новых путей, новых функций структуры мозга лежит принцип пластичности. Пластичность позволяет нервной системе под воздействием различных стимулов осуществлять реорганизацию связей для целей сохранения основной функции или для реализации новой функции.
Пластичность позволяет нервным центрам реализовать функции, которые ранее им не были присущи, но благодаря имеющимся и потенциальным связям эти центры становятся способными участвовать в компенсации нарушенных в других структурах функций. Полифункциональные структуры обладают большими возможностями пластичности. В связи с этим неспецифические системы мозга, ассоциативные структуры, вторичные зоны проекций анализаторов, как имеющие значительное число полифункциональных элементов, более способны к пластичности, чем зоны первичных проекций анализаторов. Четким примером пластичности нервных центров является классический опыт П.К. Анохина с изменением связей центров диафрагмального и плечевого нервов.
В этом опыте были перерезаны диафрагмальный и плечевой нервы и центральный конец диафрагмального нерва был присоединен к периферическому концу плечевого, и, наоборот, центральный конец плечевого нерва к периферическому диафрагмального. По истечении некоторого времени после операции у животного восстанавливались правильная регуляция дыхания и правильная последовательность произвольных движений. Следовательно, нервные центры перестроили свою функцию таким образом, как этого требовала периферическая мышечная система, с которой была установлена новая связь. На ранних этапах онтогенеза перестройки такого типа более совершенны и динамичны.
Рефлекторный принцип функционирования. Наиболее существенную роль в компенсации дисфункций структур мозга играет рефлекторный принцип его функционирования. Каждая новая рефлекторная связь между структурами мозга является новым его состоянием, позволяющим реализовывать требуемую в данный момент функцию.
В настоящее время взаимодействие полушарий головного мозга понимается как взаимодополняющее, взаимно компенсирующее в реализации различных функций центральной нервной системы. Несмотря на то, что каждое полушарие выполняет ряд специфичных для него функций, нужно иметь в виду, что любая функция мозга, выполняемая левым полушарием, может быть выполнена и правым полушарием. Речь идет только о том, насколько успешно, быстро, надежно, полно формируются рефлексы.
Читайте также: