
Опишите участие гиппокампа в процессах консолидации кратко
Обновлено: 02.07.2024
Гиппокамп находится в височной доле мозга. Судя по некоторым данным, гиппокамп и медиальная часть височной доли, т.е. часть, расположенная ближе к средней плоскости тела, играют роль в процессезакрепления, или консолидации, следов памяти. Под этим подразумеваются те изменения, физические и психологические, которые должны произойти в мозгу, для того чтобы полученная им информация могла перейти в постоянную память. Даже после того, как информация уже поступила в долговременную память, некоторые ее части все еще могут подвергаться преобразованию или даже забываться, и только после этого реорганизованный материал отправляется на постоянное хранение.
По-видимому, гиппокамп и медиальная височная область участвуют в формировании и организации следов памяти, а не служат местами постоянного хранения информации. Н.М., у которого эта область мозга была разрушена, хорошо помнил события, происшедшие более чем за 3 года до операции, и это показывает, что височная область не является местом длительного хранения следов. Однако она играет роль в их формировании, о чем свидетельствует потеря у Н.М. памяти на многие события, происходившие в последние три года до операции. (Конечно, возможно и то, что процессу консолидации следов памяти у Н.М. мешала эпилепсия.)
Подобные данные получены и при исследовании больных после электрошоковой терапии (ЭШТ). Известно, что электрошок оказывает особенно разрушительное действие на гиппокамп. После электрошока больные, как правило, страдают частичной амнезией на события, происходившие в течение нескольких предшествовавших лечению лет. Память о более давних событиях сохраняется полностью.
Лэрри Сквайр (Sguire, 1984) высказал предположение, что в процессе усвоения каких-либо знаний височная область устанавливает связь с местами хранения следов памяти в других частях мозга, прежде всего в коре. Потребность в таких взаимодействиях может сохраняться довольно долго - в течение нескольких лет, пока идет процесс реорганизации материала памяти. По мнению Сквайра, эта реорганизация связана с физической перестройкой нервных сетей. В какой-то момент, когда реорганизация и перестройка закончены и информация постоянно хранится в коре, участие височной области в ее закреплении и извлечении становится ненужной.
Помимо фаз кратковременной и долговременной памяти, характерных для процесса запоминания, у человека существует, оказывается, два разных способа усвоения и запоминания информации в зависимости от того, что именно нужно усвоить.
Гиппокамп находится в височной доле мозга. Судя по некоторым данным, гиппокамп и медиальная часть височной доли, т.е. часть, расположенная ближе к средней плоскости тела, играют роль в процессезакрепления, или консолидации, следов памяти. Под этим подразумеваются те изменения, физические и психологические, которые должны произойти в мозгу, для того чтобы полученная им информация могла перейти в постоянную память. Даже после того, как информация уже поступила в долговременную память, некоторые ее части все еще могут подвергаться преобразованию или даже забываться, и только после этого реорганизованный материал отправляется на постоянное хранение.

По-видимому, гиппокамп и медиальная височная область участвуют в формировании и организации следов памяти, а не служат местами постоянного хранения информации. Н.М., у которого эта область мозга была разрушена, хорошо помнил события, происшедшие более чем за 3 года до операции, и это показывает, что височная область не является местом длительного хранения следов. Однако она играет роль в их формировании, о чем свидетельствует потеря у Н.М. памяти на многие события, происходившие в последние три года до операции. (Конечно, возможно и то, что процессу консолидации следов памяти у Н.М. мешала эпилепсия.)
Подобные данные получены и при исследовании больных после электрошоковой терапии (ЭШТ). Известно, что электрошок оказывает особенно разрушительное действие на гиппокамп. После электрошока больные, как правило, страдают частичной амнезией на события, происходившие в течение нескольких предшествовавших лечению лет. Память о более давних событиях сохраняется полностью.
Лэрри Сквайр (Sguire, 1984) высказал предположение, что в процессе усвоения каких-либо знаний височная область устанавливает связь с местами хранения следов памяти в других частях мозга, прежде всего в коре. Потребность в таких взаимодействиях может сохраняться довольно долго - в течение нескольких лет, пока идет процесс реорганизации материала памяти. По мнению Сквайра, эта реорганизация связана с физической перестройкой нервных сетей. В какой-то момент, когда реорганизация и перестройка закончены и информация постоянно хранится в коре, участие височной области в ее закреплении и извлечении становится ненужной.
Помимо фаз кратковременной и долговременной памяти, характерных для процесса запоминания, у человека существует, оказывается, два разных способа усвоения и запоминания информации в зависимости от того, что именно нужно усвоить.
Лимбическая система: парагиппокампальная извилина и гиппокампальный комплекс
На рисунке ниже схематично представлены основные подкорковые структуры лимбической системы.
Корковая и подкорковая части лимбической системы, медиальная поверхность.
ДМЯ — дорсальное медиальное ядро таламуса. Трехмерная компьютерная реконструкция мозга умершего человека, на которой показано соотношение структур лимбической системы с желудочками мозга.
Основные клетки основания и гиппокампа — пирамидальные клетки; основные клетки зубчатой извилины — гранулярные клетки. Дендриты гранулярных и пирамидальных клеток имеют шипики. В гиппокампальном комплексе также имеется множество вставочных тормозных ГАМК-ергических нейронов.
1. Афферентные связи. Основной источник афферентных волокон, идущих к гиппокампальному комплексу,— перфорантный путь, который проецируется от энторинальной коры на дендриты гранулярных клеток зубчатой извилины. От основания гиппокампа начинается второй афферентный путь, альвеолярным (от alveus), который идет к пучку волокон на вентральной поверхности гиппокампа, лотку гиппокампа (alveus).
Аксоны гранулярных клеток называют мшистыми волокнами; они образуют синапсы с пирамидальными нейронами сектора САЗ. Аксоны нейронов САЗ следуют к бахромке; перед этим они отдают коллатерали Шаффера, которые делают обратный путь от САЗ до СА1. Нейроны СА1 отдают волокна к энторинальной коре.
Слуховая информация поступает в гиппокамп от ассоциативной коры верхней и средней височных извилин. Надкраевая извилина (поле 40) передает закодированную информацию о расположении частей тела (схема тела) и внешнем (зрительном) пространстве. От нижней поверхности затылочно-височной области информация о форме и цвете объектов (а также лицах) поступает на участок коры, который называют периринальной, или трансринальной, корой. Этот участок коры расположен сразу латеральнее от энторинальной коры. Отсюда эти нервные импульсы поступают в гиппокамп. Возвратные волокна, идущие от энторинальной коры к периринальной, связаны с префронтальной корой и корой височного полюса.
Помимо отдельных афферентных связей, перечисленных выше, гиппокамп получает обширные связи от нескольких других источников, которые поступают к нему преимущественно по своду.
• Обширные холинергические волокна, которые имеют особое значение для функционирования памяти, идут от ядра перегородки.
• Норадренергическую иннервацию обеспечивает голубое пятно.
• Серотонинергические волокна идут от ядра шва среднего мозга. Связь между истощением запасов серотонина и большим депрессивным расстройством описана в отдельной статье на сайте.
• Дофаминергические волокна идут от вентральной покрышечной области среднего мозга. Связь между дофамином и шизофренией рассмотрена в последующей отдельной статье на сайте.
2. Эфферентные связи. Наиболее обширные эфферентные связи гиппокампальная формация имеет с ассоциативными участками коры больших полушарий, которые направляются сюда через эн-ториальную кору. Вторая группа волокон представлена сводом. Свод — прямое продолжение бахромки, в которую входят аксоны от основания и собственно гиппокампа. Ножка свода делает изгиб под мозолистым телом, где она соединяется со второй своей половиной и формирует тело. Спайка свода соединяет правый и левый гиппокампы. Спереди тело разделяется на столбы свода. Огибая переднюю комиссуру, каждый свод разделяется, отдавая прекомиссуральные волокна к области перегородки и посткомиссуральные волокна к гипоталамусу, сосцевидному телу и медиальному пучку переднего мозга. От сосцевидного тела волокна направляются к переднему ядру таламуса, от которого, в свою очередь, отходят проекции к поясной коре. Так завершается круг Папеца, направляющийся от поясной коры к гиппокампу, а затем возвращающийся обратно к поясной коре через свод, сосцевидное тело и переднее ядро таламуса.
3. Гиппокампальный комплекс и память. Факты того, что г иппокампальный комплекс выполняет мнемонические (связанные с памятью) функции, достаточно подробно обсуждают в литературе по психологии. Ниже приведены некоторые ключевые моменты.
4. Терминология:
• Краткосрочная память: задержка в памяти одной или нескольких единиц информации в течение короткого промежутка времени (например, новый телефонный номер во время нажатия на кнопки).
• Долгосрочная (длительная) память: хранящаяся в головном мозге информация, к которой можно обратиться в нужный момент. Существует два вида долгосрочной памяти: эксплицитная и имплицитная.
• Эксплицитная память касается любых фактов или событий, которые можно произвольно эксплицитно озвучить (декларировать). Иначе эксплицитную память называют декларативной. Также используют термин эпизодическая память, понимая под ней собрание автобиографических воспоминаний о собственном опыте. Другой термин — семантическая память — был предложен для использования в контексте воспоминания смысла устных и письменных слов, однако в настоящее время к семантической памяти относят также знание отдельных фактов и понятий.
• Имплицитная память касается выполнения выученных двигательных актов, например езды на велосипеде или сбора пазла. Часто также используют термин процедурная память.
• Рабочая память: быстрое извлечение из долгосрочной памяти нескольких единиц информации, необходимых для выполнения какой-либо текущей задачи. Например, при управлении автомобилем вдоль знакомого маршрута мы принимаем нужные решения, основываясь на предшествующем опыте.
• Консолидация: процесс перевода новой информации в долгосрочную память. Новая фактическая информация передается на ассоциативные участки гиппокампального комплекса для кодирования. После длительной обработки эта же информация в закодированном виде поступает обратно на ассоциативные участки коры. После этого (за исключением автобиографических воспоминаний) ее использование более не зависит от гиппокампального комплекса.
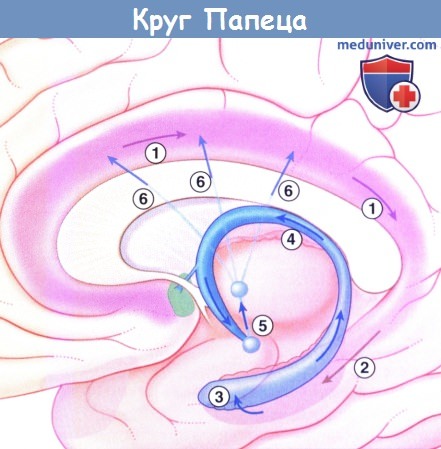
Круг Папеца.
1. Нейроны, проецирующиеся назад к поясной коре.
2. Волокна к энторинальной коре.
3. Волокна к гиппокампу.
4. Свод.
5. Сосцевидно-таламический путь.
6. Волокна от переднего ядра таламуса к поясной коре.
5. Клинические и экспериментальные наблюдения. При двустороннем повреждении или удалении передних отделов гиппокампального комплекса у пациента развивается антероградная амнезия, при которой больной не может вспомнить о событиях, произошедших буквально несколько минут назад. Когда пациента просят назвать какой-то общеизвестный предмет, он делает это без каких-либо затруднений, поскольку долгосрочные воспоминания сохранены. Однако если этот же предмет показать ему несколько минут спустя, пациент не сможет вспомнить, что уже видел его буквально только что. Нарушена эксплицит-ная/декларативная память.
Процедурная (практическая) память сохранена. Если пациента попросить собрать пазл, он сделает это без затруднений. Если затем попросить его сделать это на следующий день, он справится быстрее, но при этом не будет помнить о том, что уже собирал его вчера. Для работы процедурной памяти участие гиппокампа не требуется. Как уже было отмечено, базальные ганглии представляют собой хранилище рутинных двигательных программ, а мозжечок отвечает за двигательную адаптацию к измененным условиям среды.
Долговременная потенциация (ДВП) особенно выражена в зубчатой извилине и гиппокампе. Этот феномен необходим для сохранения (консолидации) отдельных воспоминаний. В экспериментальных условиях ДВП наблюдают в первую очередь в связях между перфорантным путем с гранулярными клетками зубчатой извилины, а также между коллатералями Шаффера и СА1. При сильной быстрой (миллисекунды) стимуляции перфорантного пути или коллатералей Шаффера в иннервируемых ими клетках наблюдают длительное (часы) повышение чувствительности к новым стимулам. При обсуждении болевой сенситизации, ДВП запускает в конечных нейронах каскад биохимических реакций, следующий за активацией глутаматных рецепторов. При повторной стимуляции циклический аденозин-3‘,5‘-монофосфат (цАМФ) повышает скорость активации протеинкиназ, участвующих в фосфорилировании белков, которые регулируют транскрипцию. В результате происходит повышение продукции белков (в том числе ферментов), которые требуются для синтеза медиаторов, а также других белков, необходимых для построения дополнительных каналов и цитоскелета синапсов.
Холинергическая активность гиппокампа имеет большое значение для обучения. Во время экспериментов на добровольцах назначение препарата скополамина, нарушающего холинергическую передачу, приводило к серьезным нарушениям при запоминании имен и чисел. В то же время назначение ингибитора холинэстеразы (физостигмин) приводило к улучшению запоминания тех же самых чисел и имен. В клинике выраженное снижение холинергической активности гиппокампа наблюдают у пациентов с болезнью Альцгеймера — заболеванием, основным симптомом которого служит нарушение памяти.
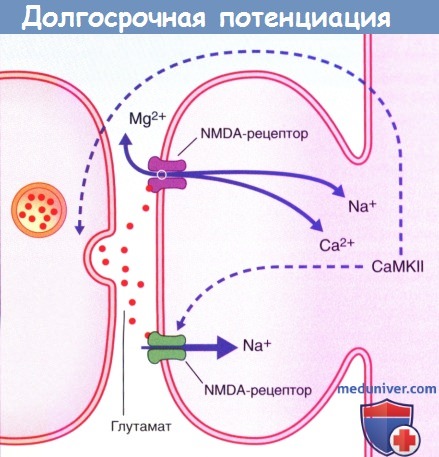
Долгосрочная потенциация.
CaMKII — Са 2+ /кальмодулин-зависимая протеинкиназа II;
АМРА receptor—АМРА-рецептор (рецептор а-амино-3-гидрокси-5-метил-4-изоксазолпропионовой кислоты.
До настоящего времени не установлено, какое значение в работе памяти имеет свод мозга. Косвенные данные можно получить из наблюдений за пациентами с диэнцефалической амнезией, при которой антероградная амнезия развивается вследствие двустороннего повреждения промежуточного мозга. Такое повреждение может сопровождаться нарушением волокон круга Папеца, который соединяет свод с поясной извилиной (через сосцевидное тело и переднее ядро таламуса). Сильнее всего страдает реляционная память (возможность вспомнить звуки или ощущения, сопровождавшие определенное событие, например звук водопада или чувство брызг воды на лице).
- Функции переднего и заднего гиппокампа. Длина гиппокампа составляет около 8 см. Существуют доказательства того, что в переднезаднем направлении гиппокамп разделен на функциональные области, одна из которых обрабатывает новую информацию, а вторая — уже известную информацию. Например, во время чтения наиболее активна передняя часть гиппокампа слева; при повторном же представлении одного и того же текста более активным становится задняя часть гиппокампа, вероятно, вследствие того, что именно она отвечает за перевод новой информации в долгосрочную память.
- Долгосрочная память и медиальная височная доля. Автобиографические воспоминания чаще всего являются зрительными. Мы способны представлять себе сцены из нашего прошлого, иногда даже из детства. Согласно клиническим данным, повреждение медиальной височной доли может приводить к значительному выпадению эгоцентрических (личных) воспоминаний, в то время как аллоцентрические (неличные) воспоминания, например о каком-либо месте или предмете, остаются сохранены.
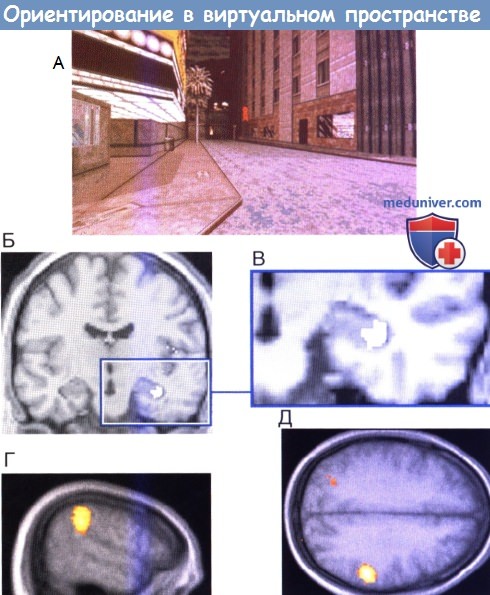
Ориентирование в виртуальном пространстве.
(А) Сцена городского города. Испытуемый, используя клавиши, должен идти по улицам, избегая препятствия. Во время исследования проводят ПЭТ головного мозга.
(Б), (В) Повышается активность правого гиппокампа и (Г), (Д) правой надкраевой извилины.
Ориентация МР-изображений (справа, слева) изменена для облегчения понимания.

Действительно, способность фиксировать, сохранять и использовать информацию, полученную в ходе пережитого опыта, представляет собой одно из важнейших эволюционных приобретений нашего вида. Совокупность наших воспоминаний, по сути, и определяет то, кто мы есть. Мы знаем, как нас зовут, какого мы пола, сколько нам лет, где мы живем, как выглядим мы и наши родители. То, как мы реагируем на различные жизненные ситуации, зависит от того, как мы действовали в аналогичных ситуациях в прошлом. Все это утратится, потеряй мы долговременную память.
Как некоторые, наверное, уже подметили, в приведенном в начале статьи примере, произошла утрата не всей долговременной памяти, а только одной ее составляющей – эксплицитной или, как ее еще называют, декларативной. Память не представляет из себя единый процесс, происходящий в каких-то одних структурах мозга. Память – это скорее совокупность процессов, каждый из которых приспособлен для сохранения какого-то определенного вида информации на какой-то определенный промежуток времени. Иерархическая схема различных типов памяти представлена на рис. 1.

Рис. 1. Схема типов памяти
Однако для нормальной жизни нам необходимо не только помнить, что когда-либо происходило и что какое-нибудь понятие означает. Мы должны помнить, и как нам взаимодействовать с объектами, которые мы знаем. Как кататься на велосипеде? Как разговаривать с начальником? Как открыть этот надоевший шкаф? Да, прочитав эти вопросы сейчас и сконцентрировав свое внимание на них, у нас, скорее всего, возникнут в сознании некие образные воспоминания о конкретных событиях. Но если перед нами встанут эти проблемы в обычной рутинной жизни, мы вряд ли будем вспоминать прошлое, а просто сделаем все, как надо. Если нам понадобится открыть шкаф, мы не будем проговаривать последовательность действий про себя и не будем вспоминать, как открывали его в самый первый раз после покупки. Мы просто возьмемся за ручку и потянем в нужную сторону. Эту память, сохраняющую категорию информации, неосознаваемой нами в течение обычных процессов мышления, но критически важной для адаптации нас к внешней среде и корректирующей наши действия, называют имплицитной .
Перечисленные выше процессы крайне разнородны по своим нейрофизиологическим механизмам и объединяются в систему имплицитной памяти исходя скорее из их функциональной схожести, а не из-за единого биологического субстрата. Тем не менее, можно выделить некие основные структуры головного мозга, необходимые соответственно для имплицитной и для эксплицитной памяти. Эти структуры указаны на рис. 2. В механизмы имплицитной памяти вовлечены неокортекс, стриатум, миндалина, мозжечок, а в простейших случаях – проходящие через спинной мозг рефлекторные пути сами по себе. Для функционирования эксплицитной памяти необходимы медиальные височные доли, гиппокамп, а также различные отделы коры.

Principles of Neural Science, Fifth Edition, 2012
а. Структуры, необходимые для функционирования различных типов эксплицитной памяти. Striatum – полосатое тело, Neocortex – неокортекс, Amygdala – миндалевидное тело, Cerebellum – мозжечок, Reflex pathways – рефлекторные пути. Структуры, необходимые для функционирования эксплицитной памяти. Medial temporal lobe – медиальные височные доли, Hippocampus – гиппокамп.
Два вида эксплицитной памяти – семантическая и эпизодическая – в большей степени сходны между собой нейрофизиологически, однако и между ними имеются различия. В настоящий момент семантическая память исследована в меньшем объеме, и дискуссия о конкретных структурах и процессах, ответственных за ее функционирование, все еще ведется. Хотя довольно больший объем исследований свидетельствует о том, что для формирования семантических воспоминаний так же, как и для эпизодических, необходима активность гиппокампа и медиальной височной доли, ряд ученых сомневается в данной концепции. В настоящий момент наиболее изучены механизмы именно долговременной эпизодической памяти, поэтому далее речь пойдет именно о них.
Эпизодическая память не представляет собой какой-то единственный непрерывно протекающий процесс. Она состоит из совокупности сменяющих друг друга процессов, обеспечивающих накопление, эффективное хранение и использование имеющихся воспоминаний. Схематично это показано на рис. 3.

Рис.3. Схема процессов эксплицитной эпизодической памяти
Значение гиппокампа в процессе запоминания новой информации было наиболее ярко продемонстрировано американскими исследователями Scolville и Milner, описавшими в 1957-м году клинический случай пациента H.M. Этот пациент страдал от тяжелейшей эпилепсии, в связи с чем было принято решение тотально удалить оба гиппокампа. В результате этой операции у H.M. была полностью потеряна способность фиксировать новые воспоминания при сохранении до определенной степени возможности воспроизводить старые, то есть, развилась изолированная антероградная амнезия. Интересно, что, хотя H.M. больше не мог запоминать какие-то новые факты или эпизоды из текущей жизни, его имплицитная память осталась интактной. В ходе множества экспериментально-психологических и когнитивных тестов было показано, что H.M. способен к определенного рода обучению и освоению новых процедурных навыков, хотя никогда и не сможет запомнить, когда и как он им обучался.
При этом, чтобы вызвать активность энграммных нейронов, то есть, вызвать воспоминание, не нужно полностью повторить активность, идущую по антероградным путям, иначе мы бы смогли вспоминать что-либо, только увидев абсолютно идентичную картину. То есть, мы бы вспоминали о своем предыдущем дне рождения, только если наши друзья в этот год поздравляли бы нас абсолютно точно так же. Для активации энграммных нейронов гиппокампа и последующей активации сенсорной и ассоциативной коры достаточно лишь частичного совпадения стимулов. Даже запах свечей сам по себе, увиденный торт в магазине, услышанная краем уха по радио поздравительная песня может вызвать у нас развернутое воспоминание о том самом прошлом дне рождения и наших друзьях. И, конечно, вспоминать что-то можно и в полное отсутствие каких-либо ассоциативно связанных стимулов, лишь по собственному желанию. Это возможно благодаря наличию связей между гиппокампом и лобными отделами коры, отвечающими за произвольные действия и за сознательный контроль физической и психической деятельности вообще.
Кроме того, с течением времени, если воспоминание не теряет своей актуальности и сохраняется надолго, постепенно усиливаются связи между самими сенсорными и ассоциативными отделами коры, а связи с гиппокампом, наоборот, ослабевают. В результате этого для воспроизведения полноценного эпизодического воспоминания не нужна будет активация энграммных клеток гиппокампа, а достаточно будет возбудить лишь один участок коры, чтобы активировались все связанные с ним в рамках данного конкретного воспоминания (рис.4.).

Principles of Cognitive Neuroscience, 2nd ed. 2013.
Рис.4. Кодирование и воспроизведение воспоминаний. А. Кодирование; В. Воспроизведение неконсолидированного воспоминания; кодирование консолидированного воспоминания.
На данном этапе заканчивается консолидация не самых нужных для нас воспоминаний. Действительно, мы еще можем вспомнить, если постараемся, внешний вид ничем не примечательных людей, с которыми мы ехали полчаса назад в метро, но на следующий день у нас это уже, скорее всего, не получится. Однако, если те же соседи в метро были чем-то крайне примечательны, и мы целенаправленно и долго на них смотрели, то воспоминание о них, скорее всего, сохранится у нас на гораздо больший срок, что будет возможно благодаря включившимся механизмам поздней потенциации синаптических связей. Более сильный или часто повторяющийся стимул приводит к большему и более частому возбуждению пресинаптической мембраны, что приводит к большему выделению глутамата, большей активации NMDA-рецепторов и, следовательно, большей концентрации кальция внутри нейрона. Если кальция в этом нейроне достаточно много, то помимо связывания с кальмодулином, он также активирует аденилатциклазу, что, спустя несколько этапов, приведет к активации CREB (cAMP-responsive element binding protein). Фосфорилированный CREB затем связывается с CRE-последовательностями определенных генов и активирует их транскрипцию. Это приводит к долговременным клеточным изменениям.

Рис. 5. Биохимические механизмы ранней и поздней потенциации.
Подытожив, хочется отметить, что в данной статье крупными мазками были описаны лишь основные механизмы, связанные с функционированием лишь одного вида памяти – эксплицитной эпизодической. Память представляет из себя невероятно сложный и обширный предмет исследования, разбираться в котором мы только начинаем. Однако в настоящий момент огромное количество исследовательских групп по всему миру углубляет наши знания о памяти. И, скорее всего, в самое ближайшее время именно в этой области стоит ждать громких открытий, детально объясняющих, как же все-таки на самом деле работает мозг, наш самый таинственный орган.

Одной из функций гиппокампа, выявленной в результате электрофизиологической стимуляции и наблюдения за поведением животных (противоречивые данные) и человека (более ранние наблюдения) при его повреждении, является отношение к процессам, связанным с памятью.
Все исследователи сходятся в том, что гиппокамп играет крайне важную роль в когнитивных процессах. Одна из гипотез говорит об участии гиппокампа в организации пространственной памяти или, по крайней мере, способствует реализации этого феномена. По мнению Squire et.al. (1989) поля СА1 и СА3 гиппокампа особенно вовлечены в получении и трансмиссии хорошо интегрированной информации как от, так и до неокортекса. Здесь очевидна и роль пластичности нейронов, а также NMDA-рецепторов при долговременной потенциации нейронов, в частности, расположенных в поле СА1. Ацетилхолин является еще одним медиатором обеспечивающим процессы эпизодической памяти.
Для обеспечение консолидации следов в памяти необходимо миграция возбуждения от этих аллокортикальных структур к неокортексу фронтальной, поясной и темпоральной областям. Эти нейроннальные связи с их обширным распространением в коре снижают зависимость процессов связанных с памятью только от одного гиппокампа.
Читайте также: