
Опишите кратко процесс пресинаптического торможения
Обновлено: 07.07.2024
Пресинаптическое торможение
Другой вид торможения — пресинаптическое торможение — открыт в центральной нервной системе в сравнительно недавнее время и потому изучен в меньшей степени, чем постсинаптическое торможение.
Пресинаптическое торможение , как это следует уже из самого названия локализуется в пресинаптических элементах, а именно в тончайших разветвлениях аксонов перед их переходом в нервное окончание.
На этих разветвлениях — пресинаптических миналях — располагаются окончания других нервных клеток, образующие здесь особые тормозные синапсы ( рис. 181 ).
Медиаторы, выделяющиеся в этих тормозных синапсах, деполяризуют в мембрану терминалей и приводят их в стояние, подобное катодической депрессии Вериго . Это обусловливаетчастичную или полную блокаду проведения нервных импульсов к нервным окончаниям.
Пресинаптическое торможение широко распространено в центральной нервной системе.
Рис. 181.Расположение тормозных синапсов на пресинаптических разветвлениях аксонов (схема). Н — нейрон, возбуждаемый афферентными импульсами, приходящими по волокну 1; Т — нейрон, образующий тормозные синапсы на пресинаптических разветвлениях волокна 1; 2 —афферентные волокна, вызывающие активность тормозного нейрона Т.
Тормозной постсинаптический потенциал. Пресинаптическое торможение
а) Электрические явления во время торможения нейрона. Влияние тормозных синапсов на постсинаптическую мембрану. Тормозной постсинаптический потенциал. Тормозные синапсы открывают в основном хлорные каналы, что позволяет ионам хлора легко проходить через мембрану. Чтобы понять, как тормозные синапсы тормозят постсинаптический нейрон, нужно вспомнить, что мы знаем о потенциале Нернста для ионов Сl-. Мы рассчитали, что он равен примерно -70 мВ. Этот потенциал отрицательнее, чем мембранный потенциал покоя нейрона, равный -65 мВ. Следовательно, открытие хлорных каналов будет способствовать движению отрицательно заряженных ионов Сl- из внеклеточной жидкости внутрь. Это сдвигает мембранный потенциал в направлении более отрицательных значений по сравнению с покоем приблизительно до уровня -70 мВ.
Открытие калиевых каналов позволяет положительно заряженным ионам К+ двигаться наружу, что приводит к большей отрицательности внутри клетки, чем в покое. Таким образом, оба события (вход ионов Сl- в клетку и выход ионов К+ из нее) увеличивают степень внутриклеточной отрицательности. Этот процесс называют гиперполяризацией. Увеличение отрицательности мембранного потенциала по сравнению с его внутриклеточным уровнем в покое тормозит нейрон, поэтому выход значений отрицательности за пределы исходного мембранного потенциала покоя называют тормозным постсинаптическим потенциалом (ТПСП).
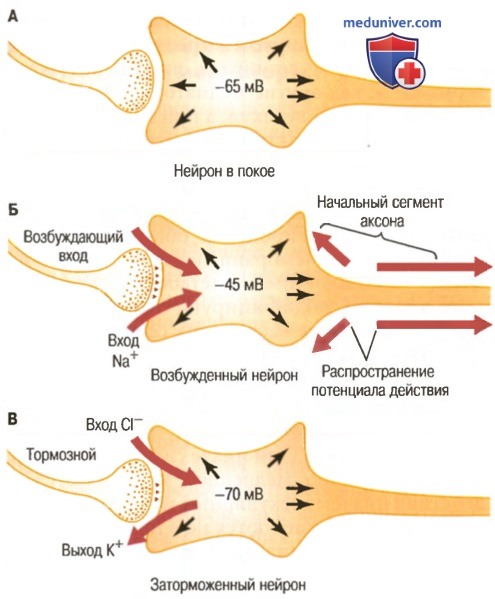
Три состояния нейрона.
А. Нейрон в покое с нормальным внутриклеточным потенциалом -65 мВ.
Б. Нейрон в состоянии возбуждения с менее отрицательным внутриклеточным потенциалом (-45 мВ), связанным с входом натрия.
В. Нейрон в заторможенном состоянии с более отрицательным внутриклеточным потенциалом (-70 мВ), что может быть связано с выходом ионов К+, входом ионов Cl- или с тем и другим
На рисунке В выше показано влияние активации тормозных синапсов, позволяющих ионам Сl-входить в клетку и/или ионам К+ выходить из нее, на мембранный потенциал. При этом он сдвигается от значения -65 мВ до более отрицательного значения -70 мВ. Этот мембранный потенциал на 5 мВ отрицательнее, чем в покое, и, следовательно, проведение нервного сигнала через синапс тормозит ТПСП, равный -5 мВ.
б) Пресинаптическое торможение. Кроме постсинаптического торможения, вызываемого тормозными синапсами, действующими на мембране нейрона, часто происходит торможение другого типа, которое развивается на пресинаптических терминалях, прежде чем сигнал достигает синапса. Этот тип торможения, называемый пресинаптическим торможением, осуществляется следующим путем.
Причиной пресинаптического торможения является выделение тормозного медиатора на наружную поверхность пресинаптических нервных волокон, прежде чем их собственные окончания достигнут поверхности постсинаптического нейрона. В большинстве случаев тормозным медиатором является ГАМК. При этом развивается специфический эффект, связанный с открытием анионных каналов, что позволяет большому числу ионов СГ диффундировать в терминальное волокно. Отрицательные заряды этих ионов тормозят синаптическое проведение, нейтрализуя большую часть возбуждающего эффекта положительно заряженных ионов Na+, которые также входят в терминальные волокна, когда здесь возникает потенциал действия.
Пресинаптическое торможение происходит во многих сенсорных путях нервной системы. Смежные сенсорные волокна часто взаимно тормозят друг друга, что сводит к минимуму боковое распространение и смешивание сигналов в чувствительных трактах.
Подробнее мы обсудили важность этого феномена в отдельных статьях на сайте - просим вас пользоваться формой поиска выше.
Видео физиология торможения в ЦНС - профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Пресинаптическое торможение первоначально было выявлено также в спинном мозге в опыте с регистрацией активности мотонейронов моносинаптической рефлекторной дуги|дуги при раздражении антагонистических мышечных нервов(Фрэнк, Фоуртес). Так, известно, что раздражение первичных афферентов мышечных веретён сопровождается возбуждением гомонимных α-мотонейронов(α-мотонейронов этой же мышцы). Однако опережающее раздражение афферентов сухожильных рецепторов мышц-антагонистов предотвращает возбуждение активируемыхα-мотонейронов . Интересно,что мембранный потенциал и возбудимость исследуемыхα-мотонейронов не изменялись либо регистрировался низкоамплитудный ВПСП,недостаточный для возникновения ПД.Посколькув опыте исследовались мотонейроны в составе моносинаптической рефлекторной дуги|дуги, было очевидно, что они не возбуждаются вследствие процессов, происходящих в пресинаптическом окончании, что определяет название этого вида торможения.
Механизм пресинаптического торможения.Электрофизиологическое изучение процессов на уровне пресинаптических окончаний в выше описанном опыте показало, что здесь регистрируется выраженная и продолжительная деполяризация,которая ведёт к развитию торможения. В очаге деполяризации нарушается процесс распространения возбуждения,следовательно,поступающие импульсы, не имея возможности пройти зону деполяризации в обычном количестве и с обычной амплитудой, не обеспечивают выделение медиатора в синаптическую щель в достаточном количестве, поскольку мало|мало ионов Са 2+ входит в нервное окончание – пост синаптический нейрон не возбуждается, его функциональное состояние, естественно, остаётся неизменным. Деполяризацию пресинаптической терминали вызывают специальные тормозные вставочные клетки, аксоны которыхобразуют синапсы на пресинаптическихокончаниях аксона-мишени.Торможение(деполяризация) после одного афферентного залпа продолжается 300–400 мс, медиаторо мявляется ГАМК, которая действует наГАМК1-рецепторы.
Деполяризацияявляется следствием повышенияпроницаемости для Cl–,в результате чего он выходит из клетки.Этот факт свидетельствует о том, что всоставе мембран пресинаптическихтерминалей имеется хлорный насос,обеспечивающий транспорт Cl–внутрьклетки вопреки электрическому градиенту.Под действием ГАМК тормозных нейронови последующего повышения проницаемостимембраны для Cl–ионыCl– начинаютвыходить наружу согласно электрическомуградиенту. Это приводит к деполяризациипресинаптических терминалей и ухудшениюих способности проводить импульсы.
Полагаюттакже, что деполяризация пресинаптическихтерминалей может возникнуть принакоплении К+ вмежклеточной жидкости в результатеповышенной активности нервных окончанийи соседних нервных клеток. В этом случаетакже ухудшается проводимостьпресинаптических терминалей из-заустойчивого снижения мембранногопотенциала в связи с уменьшениемконцентрационного градиента для К+.Роль ГАМК2-рецепторовна пресинаптических окончаниях изученанедостаточно.
Разновидностипресинаптического торможениятакжеизучены недостаточно. По-видимому,имеются те же варианты, что и дляпостсинаптического торможения. Вчастности, на рис. 4.12 представленопараллельное и латеральное пресинаптическоеторможение. Однако возвратноепресинаптическое торможение на уровнеспинного мозга (по типу возвратногопостсинаптического торможения) умлекопитающих обнаружить не удалось,хотя у лягушек оно выявлено.
Вреальной действительности взаимоотношениявозбуждающих и тормозных нейроновзначительно сложнее, чем представленона рис. 4.12, тем не менее всё|все вариантыпре- и постсинаптического торможенияможно объединить в две группы: 1) когдаблокируется собственный путь самимраспространяющимся возбуждением спомощью вставочных тормозных клеток(параллельное и возвратное торможение)и 2) когда блокируются другие нервныеэлементы под влиянием импульсов ксоседним возбуждающим нейронам свключением на пути тормозных клеток(латеральное и прямое торможение).Поскольку тормозные клетки сами могутбыть заторможены другими тормозныминейронами (торможение торможения), этоможет облегчить распространениевозбуждения.
Рольторможения.Обаизвестных вида торможения со всеми ихразновидностями выполняют охранительнуюроль. Отсутствиеторможения привело бы к истощениюмедиаторов в аксонах нейронов ипрекращению деятельности ЦНС.
Торможениеиграет важную роль в обработке поступающейв ЦНС информации. Особенноярко выражена эта роль у пресинаптическоготорможения. Оно более точно регулируетпроцесс возбуждения, поскольку этимторможением могут быть заблокированыотдельные нервные волокна|волокна. К одномувозбуждающему нейрону могут подходитьсотни и тысячи импульсов по разнымтерминалям. Вместе с тем число дошедшихдо нейрона импульсов определяетсяпресинаптическим торможением. Торможениелатеральных путей обеспечивает выделениесущественных сигналов из фона. Блокадаторможения ведёт к широкой иррадиациивозбуждения и судорогам, например привыключении пресинаптического торможениябикукулином.
Торможениеявляется важным фактором обеспечениякоординационной деятельности ЦНС.
2. Вторичноеторможение не связано с тормознымиструктурами, является следствиемпредшествующего возбуждения. Пессимальноеторможение(открыто Н.Е.Введенским в 1886 г.) развиваетсяв полисинаптических рефлекторных дугахпри чрезмерной активации центральныхнейронов и играет предохранительнуюроль. Оно выражается в стойкой деполяризациимембраны, приводящей к инактивациинатриевых каналов. “Торможение вследза возбуждением”развивается в нейронахнепосредственно после потенциаладействия и характерно для клеток сдлительной следовой гиперполяризацией.Таким образом, процессы торможения влокальных нейронных сетях уменьшаютизбыточную активность и участвуют вподдержании оптимальных режимовактивности нейронов.
1. Механизмыкоординации рефлекторной деятельности:реципрокная иннервация, доминанта(А.А.Ухтомский), принципы обратной связии общего конечного пути, принципсубординации.
Принципиррадиации возбуждения. Иррадиация– распространение, расширениерефлекторного ответа. Это феномен“растекания” возбуждения по нейронамцентральной нервной системы, развивающийсяили после действия сверхсильногораздражителя, или на фоне выключенияторможения. Распространение возбуждениявозможно за счёт многочисленных контактовмежду нейронами, возникающих приветвлении аксонов и дендритов вставочныхнейронов. Иррадиация позволяет увеличиватьколичество участвующих в рефлекторномответе групп мышц. Ограничивают иррадиациютормозные нейроны и синапсы.
Нафоне действия стрихнина, блокирующеготормозные синапсы, наступаютгенерализованные судороги при тактильнойстимуляции любого участка тела|тела или прираздражении рецепторов любой|любой сенсорнойсистемы. В коре больших|больших полушарийнаблюдается явление иррадиации процессаторможения.
Воснове координации рефлекторных актовлежат определённые механизмы, основанныена структурно-функциональной организацииЦНС и обозначаемые как “принципы”формирования рефлекторного ответа.
Принципреципрокной иннервации. Реципрокная(сопряжённая) координация открытаН.Е.Введенским в 1896 году. Обусловленареципрокным торможением, т.е. активацияодного рефлекса одновременно сопровождаетсяторможением второго, противоположногопо своей физиологической сущности.
Принципобщего “конечного пути“. Открытанглийским физиологом Ч.Шеррингтоном(1906). Один и тот же рефлекс (например,сокращение мышцы) может быть вызванраздражением различных рецепторов,т.к. один и тот же конечный a-мотонейронпередних рогов спинного мозга входитв состав многих рефлекторных дуг.Рефлексы, дуги|дуги которых имеют общийконечный путь, подразделяются наагонистические и антагонистические.Первые усиливают, вторые тормозят другдруга, как бы конкурируя за конечныйрезультат. В основе подкрепления лежитконвергенция и суммация, в основеконкуренции за конечный путь – сопряжённоеторможение.
Принципобратной связи. Любой|Любой рефлекторныйакт контролируется благодаря обратнойсвязи с центром. Обратная связь состоитво вторичной афферентации, поступающейв ЦНС от рецепторов, которые возбуждаютсяпри изменении функциональной активностирабочего органа|органа. Например, потенциалыдействия, обусловленные возбуждениемрецепторов мышц, сухожилий и суставных|суставныхсумок сгибающейся конечности, в процессеосуществления акта сгибания поступаютво всё|все структуры ЦНС, начиная от центровспинного мозга. Различают обратнуюсвязь положительную (усиливающуюрефлекс, который является источникомобратной афферентации) и отрицательную,когда рефлекс, её вызывающий, тормозится.Обратная связь лежит в основе саморегуляциифункций организма.
Принципотдачи. Феномен отдачи состоит в быстройсмене одного рефлекса другимпротивоположного значения. Например,после сгибания конечности её разгибаниепроисходит быстрее, особенно еслисгибание было сильным. Механизм этогоявления состоит в том, что при сильномсокращении мышц возбуждаются рецепторыГольджи сухожилий, которые черезтормозные вставочные нейроны тормозятмотонейроны сгибательных мышц и образуютветвь, которая возбуждает центр мышц –разгибателей. Благодаря этому механизмуможно получить сумму рефлексов – цепныерефлексы (окончание одного рефлекторногоответа инициирует следующий) и ритмические(многократное повторение ритмичныхдвижений).
Видео по теме : Пресинаптическое торможение
Синаптическое торможение, приводящее к уменьшению эффективности возбуждающих синаптических влияний, может развиваться не только на уровне постсинаптической мембраны (как это было рассмотрено выше), но еще в пресинаптическом звене путем угнетения процесса высвобождения медиатора возбуждающими нервными окончаниями. В этом случае свойства постсинаптической мембраны не подвергаются каким бы то ни было изменениям.
Пресинаптическое торможение обнаружено в различных отделах ЦНС. Наиболее часто оно выявляется в структурах мозгового ствола и особенно в спинном мозге.
Так же как и постсинаптическое, пресинаптическое торможение осуществляется посредством специальных тормозных вставочных нейронов.

Структурной основой пресинаптического торможения являются аксоаксонные синапсы , образованные окончаниями аксонов тормозных вставочных нейронов и аксональными окончаниями возбуждающих нейронов. В этом случае окончание аксона тормозного нейрона является пресинаптическим по отношению к возбуждающему окончанию, которое в свою очередь будучи постсинаптическим по отношению к тормозному окончанию, является пресинаптическим по отношению к активируемой им нервной клетке (рис. 71).
Импульсы в пресинаптическом тормозном аксоне высвобождают медиатор (в спинном мозге это вероятнее всего γ-аминомасляная кислота), который вызывает деполяризацию возбуждающих окончаний за счет увеличения проницаемости их мембраны для С1~. Предполагается, что указанная деполяризация вызывает уменьшение амплитуды потенциала действия, приходящего в возбуждающее окончание, что в свою очередь уменьшает количество высвобождаемого им медиатора, вследствие чего амплитуда возбуждающего постсинаптического потенциала падает.
Другим механизмом пресинаптического торможения может быть уменьшение входящего внутрь потока Са²⁺ , воздействующего на электросекреторную связь. И в этом случае пресинаптическое торможение приводит к уменьшению числа квантов медиатора, высвобождаемого возбуждающим пресинаптическим окончанием.
У млекопитающих продолжительность пресинаптического торможения значительно превосходит продолжительность постсинаптического торможения. Большая длительность пресинаптического торможения, по-видимому, обусловлена ритмической активностью тормозных вставочных нейронов.
Пресинаптическое торможение особенно эффективно при обработке информации, поступающей к нейрону по различным пресинаптическим путям. В этом случае возбуждение, поступающее по одному из синаптических входов, может быть избирательно уменьшено или даже полностью подавлено при отсутствии влияния на другие входы.
Подобного результата нельзя достичь путем воздействия на проводимость постсинаптической мембраны, как это имеет место при постсинаптическом торможении, влияющем на всю нервную клетку.
Читайте также: