
Образование мужского гаметофита у покрытосеменных растений кратко
Обновлено: 18.05.2024
Гаметофит является гаплоидной многоклеточной фазой внутри жизненного цикла высших растений и водорослей.
Фаза получает свое развитие из спор и отвечает за производство половых клеток — гамет. Ближе к середине 19 века впервые стали говорить о чередовании гаметофита и спорофита. Вильгельм Гофмейстер стал автором этой теории.
Развитие гаметофита происходит из гаплоидной споры. Он имеет специализированные органы — гаметангии.
Гаметангии встречаются 2 типов:
- антеридии. Это гаметангии, производящие гаметы мужского типа или спермии;
- архегонии. Это гаметангии, производящие гаметы женского типа или яйцеклетки.
Несмотря на различия форм и размеров, антеридии и архегонии отвечают за выполнение одной функции: функции образования половых клеток, дающих при слиянии обновленный хромосомный набор. Такой тип размножения появился как прогрессивная черта в ходе эволюции растительного мира.
Гаметы оплодотворяются внутри архегония. Далее происходит формирование спорофита с двойным набором хромосом. В первое время существования такой спорофит имел зависимость от гаметофита. Можно говорить о примитивности такого типа образования половых клеток в ходе эволюции.
Почти все многоклеточные водоросли характеризуются типом оплодотворения, сильно зависящим от воды.
Для водорослей свойственны:
- изогамия;
- анизогамия;
- оогамия.
Образованный в результате такого оплодотворения спорофит не зависит от гаметофита и имеет самостоятельное развитие.
Особенности гаметофитов и основные их типы
При сравнении высших растений можно обнаружить, что для различных групп высших растений и водорослей характерны различные степени развития гаметофита. Для некоторых растений вообще свойственен короткий период существования гаметофита. К примеру, если речь идет о гаметофите папоротников. А вот у мхов гаметофит развивается на протяжении всей жизни.
Выделяют следующие типы гаметофитов водорослей:
- раздельнополые нитчатые гаметофиты. Красная водоросль Polysiphonia;
- крупные пластинчатые мужские гаметофиты. Красная водоросль Palmaria;
- микроталломный гаметофит. Бангиевые водоросли;
- обоеполые гаметофиты. Ульвовые водоросли.
Также есть и другие типы гаметофитов, однако для всех из них характерна гаплоидность и многоклеточная структура.
Для высших растений свойственно правильное чередование двух многоклеточных поколений — спорофита и гаметофита. Если речь идет о цветковых растениях, то им свойственны микроскопические гаметофиты. Чтобы были понятны размеры: гаметофиты помещаются внутри пыльцевого зерна. Такие гаметофиты имеют в своем составе несколько клеток. За счет микроскопических размеров, гаметофиты цветковых растений быстро распространяются и способны выживать даже при резко меняющихся условиях среды.
Зародышевый мешок — женский гаметофит цветковых растений, который располагается внутри семяпочки. Чаще всего гаметофит состоит из семи клеток, образованных в результате слияния двух ядер центральных клеток и формирования вторичного ядра.
Гаметофит может быть небольшим самостоятельным растением — это наблюдается у плаунов, хвощей и папоротников.
Заросток — гаметофит папоротников.
Для заростков плаунов, хвощей и папоротников характерна способность к фотосинтезу. Также они являются однолетними. Заростки плаунов обитают под землей — их развитие происходит на протяжении всей жизни. Им свойственно развитие на протяжении многих лет, в течение которых они получают питание, образуя симбиоз с грибами. Небольшие размеры мхов не мешают им доминировать над спорофитом.
Образование гаметофита высших растений происходит исключительно в результате митоза. Это принципиальное отличие от гаметофита животных. Тело гаметофита строится на основе гаплоидных клеток.
Гаметофаза или галофаза — фаза гаметофита высших растений.
На одном гаметофите вполне возможно одновременное развитие мужских и женских половых органов. Этот гаметофит получил название однодомного.
При формировании гаметофитом мужских и женских половых органов на разных растениях, речь идет про двудомные или раздельнополые гаметофиты.
Раздельнополые гаметофиты встречаются у всех семенных растений: мужские гаметофиты развиваются у них из микроспор, которые образуются в пылинках, а женские — из мегаспор, которые образуются в семяпочках.
Процесс полового размножения растений имеет вид конъюгации.
Конъюгация представляет собой процесс полового размножения растений, при котором происходит слияние протопластов независимых вегетативных клеток.
Встречается, что на одной особи развиваются и споры, и гаметы. Также споры могут развиваться на одном растении, а гаметы — на другом.
Спорофит — особь, на которой происходит развитие спор.
Гаметофит — особь, на которой происходит развитие гамет.
Половое поколение хвойных — оригинальное. Мужской гаметофит в этом случае представляет несколько пыльцевых зерен, развивающихся внутри шишки. Такие шишки растут у основания побегов. Определить, что перед вами мужская шишка легко: они мельче, мягче и собираются в пучки. Для каждой чешуйки характерно два пыльника, внутри которых происходит развитие половых клеток мужских особей.
Женские шишки располагаются на верхушках побегов и содержат в себе семязачатки. Внутри женских шишек происходит формирование мегаспор — в результате мейоза. Всего образуется четыре мегаспоры.
В женском гаметофите происходит развитие только одной мегаспоры: остальные образованные клетки отмирают. После этого последовательно происходят опыление, оплодотворение и формирование семян.
Как видно, гаметофит — это половое поколение растений. Эта фаза жизненного цикла свойственна водорослям, споровым и семенным высшим растениям.
Для гаметофита характерна многоклеточная структура и наличие гаплоидного набора хромосом.
У высших семенных растений отмечен только один тип полового процесса — оогамия. Кроме того, у них в результате сочетания бесполого размножения с половым образуются особые зачатки — семена, при помощи которых происходит расселение растений.
У покрытосеменных растений органом размножения является цветок.
Мужской гаметофит-пыльцевое зерно. Тычинка состоит из тычиночной нити и пыльника.
Каждый пыльник образован двумя половинками, в которых развивается по две пыльцевые камеры — микроспорангии. В камерах молодого пыльника имеются особые диплоидные клетки —микроспороциты, или материнские клетки микроспор. Каждый микроспороцит претерпевает мейоз и образует четыре микроспоры. Здесь же, внутри пыльцевого гнезда, микроспора увеличивается в размерах, ядро ее делится митотически,и образуется — вегетативное ядро и генеративная клетка. Клетка делится митозом и образуется 2 спермия.На поверхности бывшей микроспоры образуется прочная целлюлозная оболочка с несколькими округлыми порами, сквозь которые в конечном итоге прорастают пыльцевые трубки. В результате этих процессов каждая микроспора превращается в пыльцевое зерно (пыльцу) —мужской гаметофит цветковых растений.
У однодольных растений в пыльцевом зерне, находящемся в пыльнике, генеративная клетка делится митотически с последующим образованием двух неподвижных мужских гамет — спермиев.
У двудольных образование спермиев происходит позже, когда пыльца попадает на рыльце пестика. Таким образом, зрелое пыльцевое зерно состоит из двух (вегетативной и генеративной) или из трех (вегетативной и двух спермиев) клеток.
Образование женского гаметофита (зародышевый мешок) происходит в семязачатке (семяпочке), находящемся внутри завязи пестика. Семязачаток — это видоизмененный мегаспорангий (нуцеллус), защищенный покровами (интегументами). Покровы на верхушке не срастаются и образуют узкий канал — пыльцевход
( микропиле). В нуцеллусе, вблизи пыльцевхода, начинает развиваться диплоидная клетка — макроспороцит. Он делится мейотически, давая четыре гаплоидные макро- или мегаспоры, обычно расположенные линейно. Три мегаспоры вскоре разрушаются, а четвертая, наиболее удаленная от пыльцевхода, развивается в зародышевый мешок.
Зародышевый мешок растет, его ядро трижды делится митотически, в результате чего образуется восемь дочерних ядер. Они располагаются по четыре двумя группами— вблизи, пыльцевхода зародышевого мешка и на противоположном полюсе. Затем от каждого полюса отходит, но одному ядру в центр зародышевого мешка. Это так называемые полярные ядра. В дальнейшем они могут сливаться, превращаясь в одно центральное, или вторичное диплоидное ядро (или их слияние происходит позднее, при оплодотворении). Остальные шесть ядер, по три на каждом полюсе, разделяются тонкими клеточными перегородками. При этом на полюсе у пыльцевхода образуется яйцевой аппарат, состоящий из яйцеклетки и двух клеток-синергид. На противоположном полюсе возникают так называемые клетки-антиподы, которые определенное время участвуют в доставке к клеткам зародышевого мешка питательных веществ, а затем исчезают. Такая восьмиядернаясемиклеточная структура — зародышевый мешок — является зрелым женским гаметофитом, готовым к оплодотворению. Образование пыльцы и зародышевого мешка у большинства растений завершается одновременно.
Оплодотворение.
Попав на рыльце пестика, пыльцевое зерно начинает прорастать. Из вeгетативнойклетки развивается длинная пыльцевая трубка, дорастающая по тканям столбика до завязи и далее —до семязачатка.Из генеративной клетки к этому моменту образуются два спермия, которые спускаются в пыльцевую трубку. Рост пыльцевой трубки стимулируют ауксины, вырабатываемые пестиками, а к завязи она направляется в результате хемотропизма. Пыльцевая трубка входит в семязачаток через пыльцевход, ее ядро разрушается, а кончик трубки при соприкосновении с оболочкой зародышевого мешка разрывается, освобождая мужские гаметы. Спермии проникают в зародышевый мешок в синергиду или в щель между яйцеклеткой и центральным ядром. Вскоре после вхождения пыльцевой трубки в зародышевый мешок синергиды и антиподы отмирают.

После этого один из спермиев оплодотворяет яйцеклетку. В результате образуется диплоидная зигота, из которой развивается зародыш нового растительного организма. Второй спермий сливается с двумя полярными ядрами (или с центральным диплоидным ядром), образуя триплоиднуюклетку, из которой впоследствии возникает питательная ткань —эндосперм. В его клетках содержится запас питательных веществ, необходимых для развития зародыша растения.
У высших семенных растений отмечен только один тип полового процесса — оогамия. Кроме того, у них в результате сочетания бесполого размножения с половым образуются особые зачатки — семена, при помощи которых происходит расселение растений.
У покрытосеменных растений органом размножения является цветок.
Мужской гаметофит-пыльцевое зерно. Тычинка состоит из тычиночной нити и пыльника.
Каждый пыльник образован двумя половинками, в которых развивается по две пыльцевые камеры — микроспорангии. В камерах молодого пыльника имеются особые диплоидные клетки —микроспороциты, или материнские клетки микроспор. Каждый микроспороцит претерпевает мейоз и образует четыре микроспоры. Здесь же, внутри пыльцевого гнезда, микроспора увеличивается в размерах, ядро ее делится митотически,и образуется — вегетативное ядро и генеративная клетка. Клетка делится митозом и образуется 2 спермия.На поверхности бывшей микроспоры образуется прочная целлюлозная оболочка с несколькими округлыми порами, сквозь которые в конечном итоге прорастают пыльцевые трубки. В результате этих процессов каждая микроспора превращается в пыльцевое зерно (пыльцу) —мужской гаметофит цветковых растений.
У однодольных растений в пыльцевом зерне, находящемся в пыльнике, генеративная клетка делится митотически с последующим образованием двух неподвижных мужских гамет — спермиев.
У двудольных образование спермиев происходит позже, когда пыльца попадает на рыльце пестика. Таким образом, зрелое пыльцевое зерно состоит из двух (вегетативной и генеративной) или из трех (вегетативной и двух спермиев) клеток.
Образование женского гаметофита (зародышевый мешок) происходит в семязачатке (семяпочке), находящемся внутри завязи пестика. Семязачаток — это видоизмененный мегаспорангий (нуцеллус), защищенный покровами (интегументами). Покровы на верхушке не срастаются и образуют узкий канал — пыльцевход
( микропиле). В нуцеллусе, вблизи пыльцевхода, начинает развиваться диплоидная клетка — макроспороцит. Он делится мейотически, давая четыре гаплоидные макро- или мегаспоры, обычно расположенные линейно. Три мегаспоры вскоре разрушаются, а четвертая, наиболее удаленная от пыльцевхода, развивается в зародышевый мешок.
Зародышевый мешок растет, его ядро трижды делится митотически, в результате чего образуется восемь дочерних ядер. Они располагаются по четыре двумя группами— вблизи, пыльцевхода зародышевого мешка и на противоположном полюсе. Затем от каждого полюса отходит, но одному ядру в центр зародышевого мешка. Это так называемые полярные ядра. В дальнейшем они могут сливаться, превращаясь в одно центральное, или вторичное диплоидное ядро (или их слияние происходит позднее, при оплодотворении). Остальные шесть ядер, по три на каждом полюсе, разделяются тонкими клеточными перегородками. При этом на полюсе у пыльцевхода образуется яйцевой аппарат, состоящий из яйцеклетки и двух клеток-синергид. На противоположном полюсе возникают так называемые клетки-антиподы, которые определенное время участвуют в доставке к клеткам зародышевого мешка питательных веществ, а затем исчезают. Такая восьмиядернаясемиклеточная структура — зародышевый мешок — является зрелым женским гаметофитом, готовым к оплодотворению. Образование пыльцы и зародышевого мешка у большинства растений завершается одновременно.
Оплодотворение.
Попав на рыльце пестика, пыльцевое зерно начинает прорастать. Из вeгетативнойклетки развивается длинная пыльцевая трубка, дорастающая по тканям столбика до завязи и далее —до семязачатка.Из генеративной клетки к этому моменту образуются два спермия, которые спускаются в пыльцевую трубку. Рост пыльцевой трубки стимулируют ауксины, вырабатываемые пестиками, а к завязи она направляется в результате хемотропизма. Пыльцевая трубка входит в семязачаток через пыльцевход, ее ядро разрушается, а кончик трубки при соприкосновении с оболочкой зародышевого мешка разрывается, освобождая мужские гаметы. Спермии проникают в зародышевый мешок в синергиду или в щель между яйцеклеткой и центральным ядром. Вскоре после вхождения пыльцевой трубки в зародышевый мешок синергиды и антиподы отмирают.
После этого один из спермиев оплодотворяет яйцеклетку. В результате образуется диплоидная зигота, из которой развивается зародыш нового растительного организма. Второй спермий сливается с двумя полярными ядрами (или с центральным диплоидным ядром), образуя триплоиднуюклетку, из которой впоследствии возникает питательная ткань —эндосперм. В его клетках содержится запас питательных веществ, необходимых для развития зародыша растения.
Отдел покрытосеменные (цветковые) самый многочисленный, он включает 235-250 тысяч видов. Его представители обитают по всему миру: от холодной тундры до жарких тропиков, отдельные виды освоили пресные и морские водоемы.

Покрытосеменные составляют большую часть массы растительного сообщества, являются звеном в цепи питания (продуцентами) - важнейшими производителями органических веществ на суше, как водоросли - в морях и океанах.
Цветок - генеративный орган покрытосеменных (цветковых), высшая ступень полового размножения. Цветок характерен только для покрытосеменных растений, ни один из других отделов подобным генеративным органом не обладает. По своему строению цветок это видоизмененный обоеполый стробил, гомологичный стробилам голосеменных.
В отличие от голосеменных, у которых семязачатки лежат открыто на семенных чешуях, у цветковых семязачаток находится в замкнутом вместилище - завязи, сформированной из плодолистика (-ов).
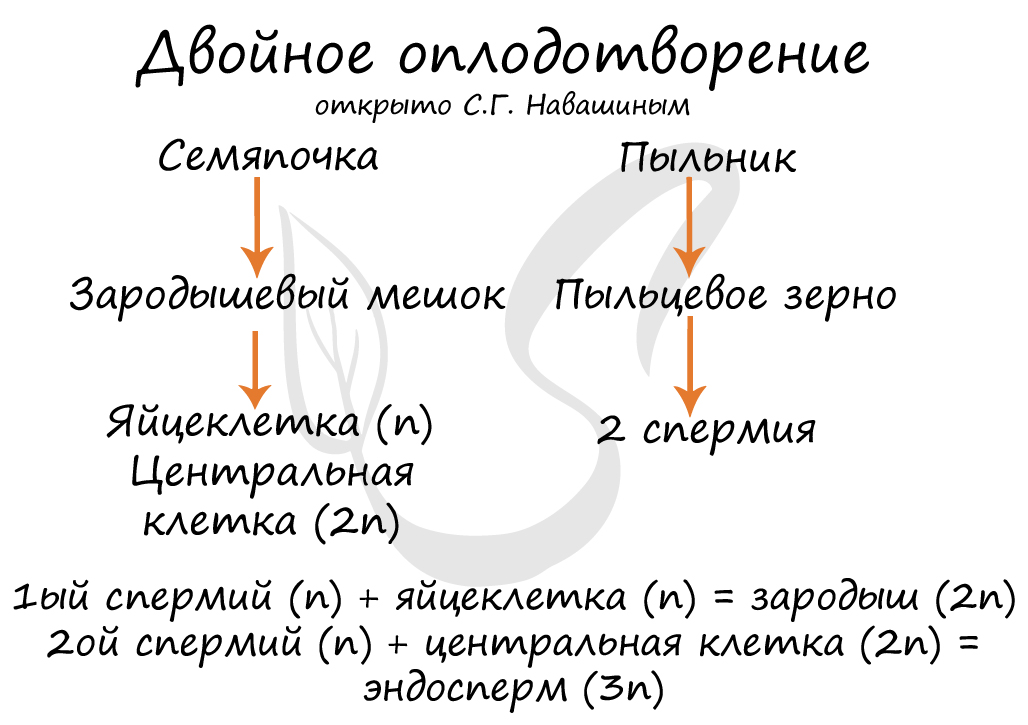
Двойное оплодотворение, открытое Навашиным Сергеем Гавриловичем, уникальное явление, характерное только для цветковых. Оно связано с тем, что в зародышевый мешок попадают два спермия, один из которых (n) сливается с центральной клеткой (2n), с образованием запасного питательного вещества - эндосперма (3n). Другой спермий (n) сливается с яйцеклеткой (n) с образованием зиготы (2n), из которой развивается зародыш.
У цветковых появляется плод - генеративный орган, служащий для защиты и распространения семян.
Ксилема - проводящая ткань, обеспечивающая восходящий ток воды и растворенных в ней минеральных солей, представлена не трахеидами, а сосудами. Во флоэме ситовидные элементы окружены клетками-спутницами.
У покрытосеменных мы не найдем антеридиев и архегониев: гаметофиты максимально редуцированы.
В процессе опыления покрытосеменных участвуют насекомые, летучие мыши, птицы. Также опыление может происходить с помощью воды или ветра.

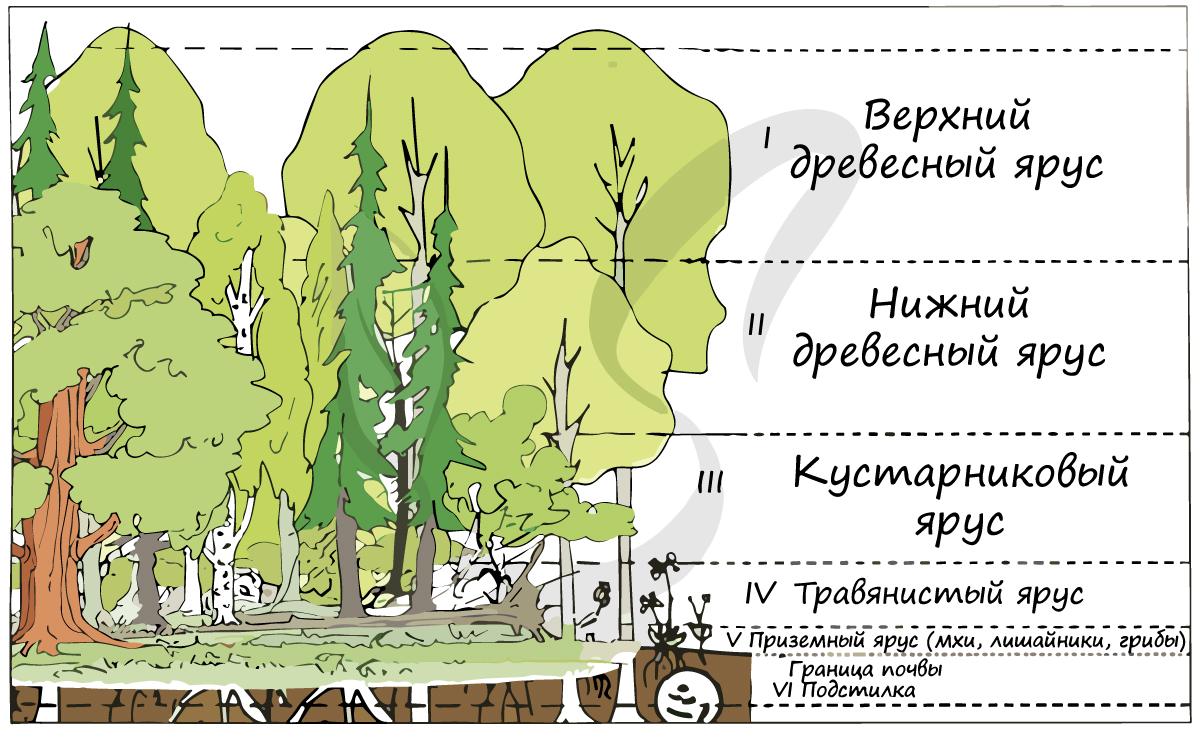
Особенностью цветковых является способность образовывать многоярусные сообщества, более устойчивые и продуктивные.
Многоярусность растительного сообщества служит приспособлением к равномерному распределению света: светолюбивые растения занимают верхний ярус, а теневыносливые растения отлично чувствуют себя в тени светолюбивых :)
Классы покрытосеменных
Отдел покрытосеменные состоит из двух классов: однодольные и двудольные. К классу двудольных относятся семейства: крестоцветные, сложноцветные, розоцветные, бобовые (мотыльковые), пасленовые. Класс однодольные включает в себя семейства: злаковые, лилейные. Для каждого класса имеются характерные признаки.
-
Двудольные - семейства: крестоцветные, сложноцветные, бобовые, розоцветные, пасленовые
-
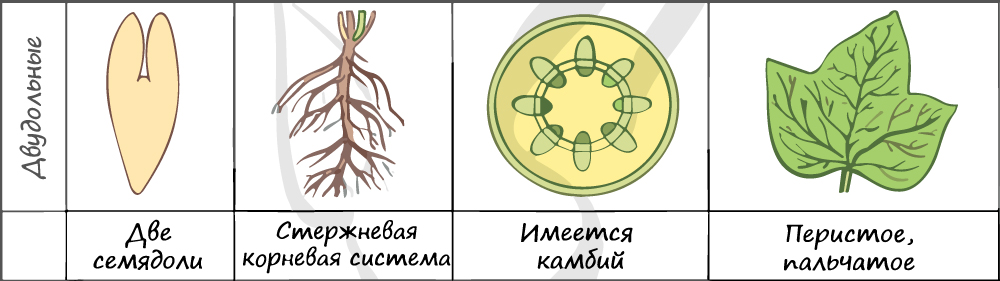
В составе зародыша обычно имеется две семядоли
В семядолях содержится запас питательных веществ. При надземном прорастании семядоли (зародышевые листья) могут выполнять функцию фотосинтеза.

Листья двудольных простые и сложные, для двудольных характерно перистое и пальчатое жилкование.
За счет камбия растения растут в толщину, возможен вторичный рост осевых органов (стебля и корня).
Корневая система чаще всего стержневого типа, с хорошо выраженным главным корнем, от которого отходят боковые корни. Главный корень развивается из зародышевого корешка.
Цветки пятичленные, реже встречаются четырехчленные. Хорошо обособлены чашечка и венчик.

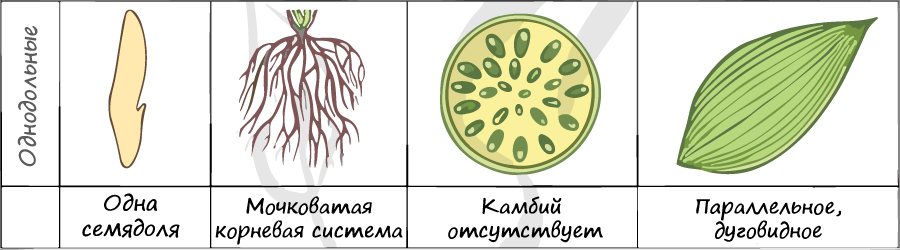
Цветок с простым околоцветником. Цветки чаще трехчленные, четырехчленные. Никогда не бывают пятичленными.
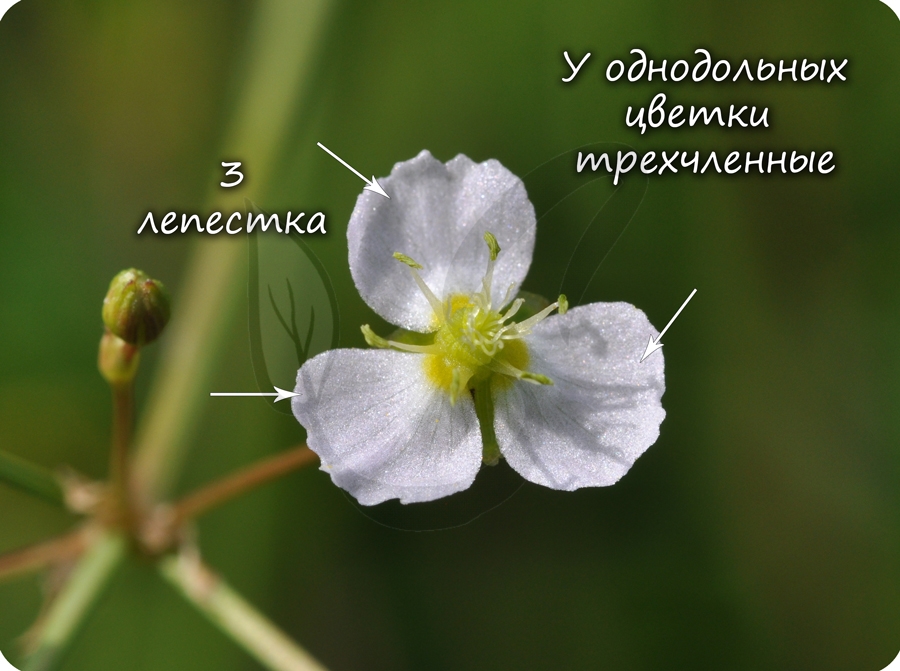
Эндосперм семени
Эндосперм (от греч. endon - внутри + греч. sperma - семя) - запасное питательное вещество, у покрытосеменных триплоидный (3n).
Эндосперм в семени есть у подавляющего большинства однодольных (лука, ландыша, пшеницы) и двудольных (тмина, хурмы, фиалки). Отсутствует эндосперм в семенах тыквенных, крестоцветных (капусты), сложноцветных (подсолнечника), бобовых (гороха, фасоли), также у - березы, липы, дуба, клена, так как на ранней стадии развития растущий зародыш поглощает эндосперм.
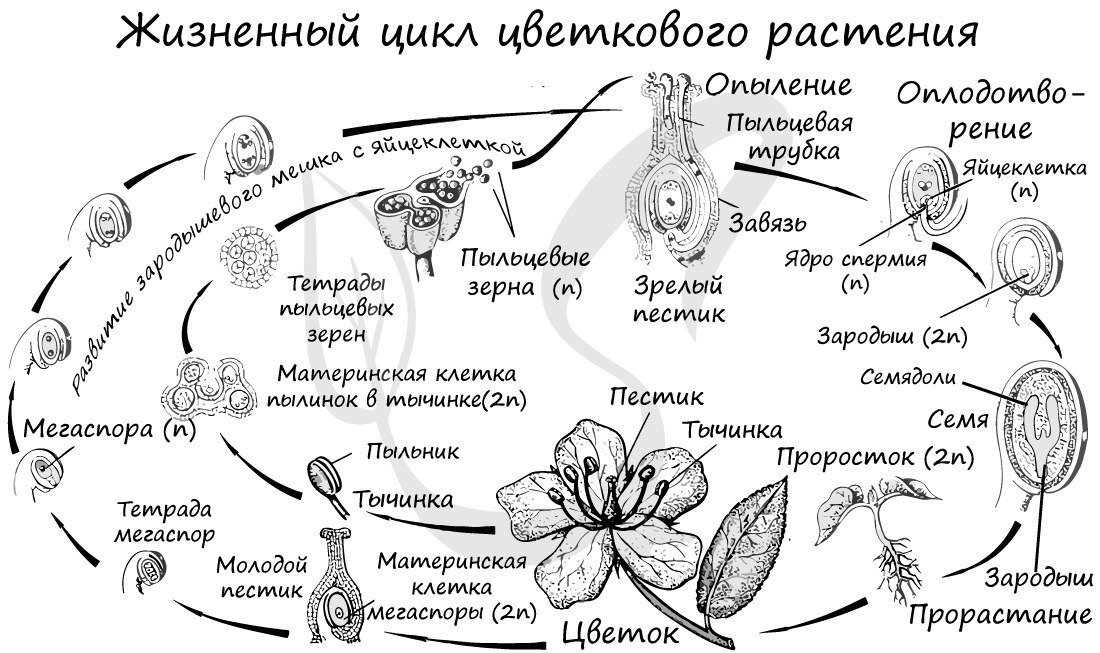
Жизненный цикл
Из генеративных почек спорофита развиваются цветки. У взрослого растения спорофита (2n) в цветке в гнездах пыльников тычинок в ходе микроспорогенеза образуется пыльцевое зерно (n) - мужской гаметофит. В завязи пестика в семязачатке формируется женский гаметофит - зародышевый мешок, внутри которого находятся центральная клетка (2n) и яйцеклетка (n).
В результате опыления (насекомым, ветром, человеком) пыльца с тычинок переносится на рыльце пестика. Пыльцевое зерно состоит из вегетативной и генеративной клеток. Вегетативная клетка начинает растворять ткани пестика, образует пыльцевую трубку и прорастает до зародышевого мешка. Генеративная клетка делится, образуя два спермия (n), из которых один сливается с центральной клеткой (2n) с образование эндосперма (3n) - запасного питательного вещества. Другой спермий (n) сливается с яйцеклеткой (n), образуя зиготу (2n).
В дальнейшем из семязачатка формируется семя, а завязь превращается в околоплодник - образуется плод. Своим внешним видом плоды привлекают животных, и те их охотно поедают) Благодаря семенной кожуре семена не подвергаются расщеплению в желудочно-кишечном тракте человека и животных. Они выходят из ЖКТ в неизменном виде и остаются способны к прорастанию: так происходит расселение растений. Попав в благоприятные условия, они прорастают в спорофит (2n). Цикл замыкается.
Значение покрытосеменных
Покрытосеменным в жизни человека отведено важное место. Только подумайте - почти все культурные растения принадлежат к этому отделу! Цветковые имеют медицинское значение, из многих растений изготавливаются лекарства. Их древесина используется для изготовления бумаги, мебели, применяются в промышленности.

Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Органом полового размножения покрытосеменных растений является цветок. Цветок — видоизмененный, укороченный, неразветвленный побег, предназначенный для образования спор и гамет и полового процесса, завершающегося образованием семян и плода.
Строение цветка
У цветка различают цветоножку, цветоложе, околоцветник, тычинки и пестики. У некоторых цветков отдельные части могут отсутствовать.
Цветки большинства видов растений имеют и тычинки, и пестики. Такие цветки называют обоеполыми (вишня, горох). Цветки, которые имеют только пестики, называют пестичными (женскими). Цветки, которые имеют только тычинки, называют тычиночными (мужскими). В зависимости от распределения однополых цветков на растениях различают: однодомные растения — растения, у которых на одних и тех же экземплярах располагаются и женские, и мужские цветки (огурец, кукуруза, дуб); двудомные растения — растения, у которых на одних экземплярах располагаются женские, а на других — мужские цветки (крапива двудомная, конопля, облепиха); многодомные растения — растения, у которых на одних и тех же экземплярах встречаются как обоеполые, так и однополые цветки в различных количественных соотношениях (гречиха, некоторые виды ясеня, клена).
Цветоножка — междоузлие под цветком. Цветки, лишенные цветоножки, называются сидячими (цветки в соцветии корзинка у подсолнечника, астры, одуванчика).
Цветоложе — укороченная стеблевая часть цветка. На ней располагаются все остальные части цветка.
Околоцветник — стерильная часть цветка, его покров. Околоцветник может быть простым (не дифференцированным на чашечку и венчик, образованным совокупностью однородных листочков, имеющих одинаковые размеры и окраску) и двойным (дифференцированным на чашечку и венчик, отличающиеся друг от друга размерами и окраской. Простой околоцветник может быть венчиковидным (образованным ярко окрашенными листочками) или чашечковидным (образованным зелеными листочками). Цветки, лишенные околоцветника (ива, тополь), называются голыми.
Чашечка — наружная часть двойного околоцветника, представляет собой совокупность чашелистиков — видоизмененных прицветных листьев. Обычно чашелистики имеют небольшие размеры и зеленую окраску. Они сходны с обычными листьями, но устроены проще.
Различают: раздельнолистную чашечку — чашечку, образованную свободными (несросшимися) чашелистиками (капуста, лютик); сростнолистную чашечку — чашечку, образованную частично или полностью сросшимися чашелистиками (картофель, табак, горох).
Венчик — внутренняя, обычно окрашенная часть двойного околоцветника. Представляет собой совокупность лепестков, часто имеющих яркую окраску. Количество лепестков венчика может быть различным. Лепестки могут быть более или менее одинаковыми (лютик,яблоня) либо отличаться размерами и формой (фиалка, горох). В результате венчик может быть правильным, неправильным или асимметричным. Венчик, как и чашечка, может быть раздельнолепестным и сростнолепестным. Раздельнолепестной венчик состоит из свободных, несросшихся лепестков. Сростнолепестной венчик состоит из сросшихся в той или иной степени лепестков. Главная функция венчика — привлечение опылителей.
Андроцей
Андроцей — совокупность тычинок одного цветка. Количество тычинок в цветке — от одной (орхидные) до нескольких сотен (некоторые кактусы). У большинства растений тычинка состоит из тычиночной нити и пыльника. Тычиночная нить — нижняя, как правило, суженная стерильная часть тычинки. Нижний конец тычиночной нити отходит от цветоложа, а верхний конец несет пыльник. Обычно тычиночные нити тонкие, длинные, округлые в сечении. Пыльник — верхняя расширенная фертильная часть тычинки. Пыльник состоит из двух половинок, соединенных связником. Каждая половинка имеет два пыльцевых гнезда (микроспорангия), в которых происходит образование микроспор, а впоследствии пылинок. Связник является продолжением тычиночной нити, через него в пыльник поступают питательные вещества.
Микроспорогенез
Микроспорогенез — процесс образования микроспор в микроспорангиях (гнездах пыльника). Микроспоры формируются из материнских клеток — микроспороцитов, имеющих диплоидный набор хромосом. В результате мейоза каждая материнская клетка образует четыре гаплоидных микроспоры. Микроспоры быстро обособляются друг от друга.
Микрогаметогенез
Микрогаметогенез — процесс образования мужских половых клеток (спермиев), происходит в пыльцевом зерне, которое является мужским гаметофитом покрытосеменных растений. Развитие мужского гаметофита происходит также в гнездах пыльников тычинок и сводится к одному митотическому делению микроспоры и формированию оболочек пыльцевого зерна. Оболочка пыльцевого зерна состоит из двух слоев: интины (внутренней, тонкой) и экзины (наружной, толстой). Каждое пыльцевое зерно содержит две гаплоидные клетки: вегетативную и генеративную. Из генеративной (спермагенной) далее образуются два спермия. Из вегетативной (сифоногенной) впоследствии образуется пыльцевая трубка.
Гинецей
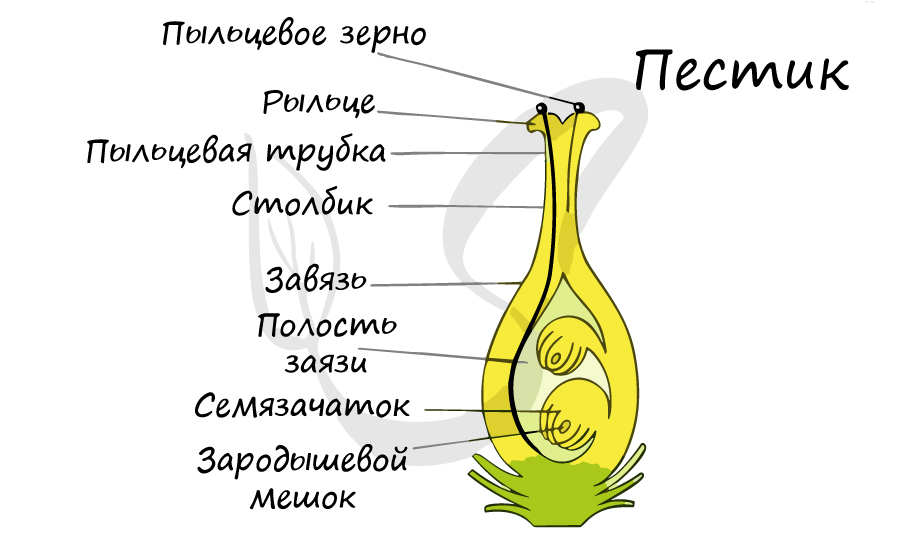
Гинецей — совокупность пестиков одного цветка. Обычно в пестике выделяют три части: завязь, столбик и рыльце.
Завязь — замкнутая, нижняя, полая часть пестика, несущая и защищающая семязачатки. Завязь бывает: верхняя, нижняя, полунижняя. В завязи может располагаться от одного (пшеница, вишня) до нескольких тысяч (мак) семязачатков. Стенки завязи выполняют функцию защиты семязачатков от неблагоприятных факторов среды (высыхание, колебание температур, поедание насекомыми и т.д.). Внутри завязи (в семязачатках) происходит мегаспорогенез и мегагаметогенез, они принимают участие в образовании околоплодника.
Столбик — средняя более или менее удлиненная стерильная часть пестика, отходящая обычно от верхушки завязи, соединяет завязь и рыльце.
Рыльце — верхняя расширенная часть пестика, предназначено для восприятия пыльцы. Рыльце может быть разнообразной формы (двухлопастное, звездчатое, перистое и т.д.) и размера в зависимости от особенностей опыления. При отсутствии столбика рыльце называют сидячим.


Семязачаток состоит из нуцеллуса (ядра) — центральной части, являющейся мегаспорангием, двух покровов — интегументов, которые при смыкании образуют узкий канал — микропиле, или пыльцевход, через который пыльцевая трубка проникает к зародышевому мешку. С помощью семяножки семязачаток прикрепляется к плаценте. Место прикрепления семязачатка к семяножке называют рубчиком. Противоположную микропиле часть семязачатка, где сливаются нуцеллус и интегументы, называют халазой.
В семязачатке происходят мегаспорогенез, мегагаметогенез и процесс оплодотворения. После оплодотворения (реже без него) из семязачатка формируется семя.

Мегаспорогенез
Процесс формирования мегаспор называется мегаспорогенезом. Он происходит в нуцеллусе семязачатка. После заложения семязачатка и формирования нуцеллуса в области микропиле начинает разрастаться одна археспориальная (спорогенная) клетка — мегаспороцит, или материнская клетка мегаспор.
Материнская клетка мегаспор имеет диплоидный набор хромосом. У большинства покрытосеменных из нее путем мейоза образуются четыре гаплоидные мегаспоры. Из них лишь одна (обычно нижняя, обращенная к халазе, реже верхняя, обращенная к микропиле) дает начало женскому гаметофиту — зародышевому мешку. Остальные мегаспоры отмирают.
Мегагаметогенез
Процесс формирования женских половых клеток происходит в зародышевом мешке. Формирование женского гаметофита начинается с разрастания мегаспоры, которая далее три раза делится митозом. В результате этого образуются восемь клеток, которые располагаются следующим образом: три — на одном полюсе зародышевого мешка (микропилярном), три — на другом (хадазальном), две — в центре. Две оставшиеся сливаются в центре клетки, образуя диплоидную центральную клетку зародышевого мешка. Одна из трех клеток, расположенных на микропилярном полюсе, отличается большими размерами и является яйцеклеткой. Две рядом расположенные клетки являются вспомогательными и называются синергидами. Группа из трех клеток, находящихся на противоположном, халазальном полюсе, называется антиподом. Таким образом, сформированный женский гаметофит включает шесть гаплоидных клеток (яйцеклетка, две клетки-синергиды, три клетки-антипода) и одну диплоидную клетку.
Оплодотворение. Образование семян и плодов
Купить проверочные работы
и тесты по биологии




Процессу оплодотворения предшествует опыление — перенос пыльцы от пыльцевых мешков тычинок к рыльцам пестиков. Попав на рыльце пестика, под воздействием веществ, выделяемых пестиком, пыльца начинает прорастать: образуется пыльцевая трубка, внедряющаяся в ткань рыльца. Кончик пыльцевой трубки выделяет вещества, размягчающие ткань рыльца и столбика. В процессе формирования пыльцевой трубки принимает участие сифоногенная клетка. По мере роста пыльцевой трубки в нее переходит спермагенная клетка, которая делится митозом с образованием двух спермиев (у некоторых растений спермагенная клетка дает начало двум спермиям еще до прорастания пыльцы). Пыльцевая трубка продвигается по столбику пестика и врастает в зародышевый мешок, как правило, через микропиле. После проникновения в зародышевый мешок кончик пыльцевой трубки разрывается, и спермии попадают внутрь. Один из спермиев сливается с яйцеклеткой, образуя диплоидную зиготу. Второй спермий сливается с центральной клеткой зародышевого мешка, образуя триплоидную клетку, из которой далее формируется эндосперм (питательная ткань) семени, обеспечивающий питание зародыша. Синергиды и антиподы дегенерируют. Вышеописанный процесс получил название двойного оплодотворения. Двойное оплодотворение у цветковых растений было открыто в 1898 году русским ботаником С.Г. Навашиным.
После двойного оплодотворения из яйцеклетки формируется зародыш семени, из центрального ядра зародышевого мешка — эндосперм, из интегументов — семенная кожура, из всего семязачатка — семя, а из стенок завязи — околоплодник. В целом из завязи пестика формируется плод с семенами.

Видеоурок способствует формированию представлений о способах опыления цветковых растений, выявляет приспособления к различным способам опыления. Основная цель урока – рассмотреть особенности двойного оплодотворения и его преимущества. В ходе видеоурока подробно рассказывается о процессах микроспорогенеза и макроспорогенеза, их сущности.

В данный момент вы не можете посмотреть или раздать видеоурок ученикам
Чтобы получить доступ к этому и другим видеоурокам комплекта, вам нужно добавить его в личный кабинет, приобретя в каталоге.
Получите невероятные возможности



Конспект урока "Двойное оплодотворение покрытосеменных растений"
Размножение – это одно из обязательных свойств любого живого организма. Оно заключается в увеличении числа особей.
Различают бесполое и половое размножение растений.
Бесполое размножение подразделяют на спорообразование и вегетативное.
Половое размножение происходит при помощи особых половых клеток –гамет.
При бесполом размножении быстро увеличивается численность вида, все потомки имеют абсолютно такой же генотип, что и родительская особь. А также не происходит увеличения генетического разнообразия, которое может оказаться очень полезным при изменении условий существования вида.
По этой причине большинство живых организмов на Земле размножаются половым путём.
Сущность полового размножения заключается в слиянии генетической информации родителей, благодаря чему генетическое разнообразие в потомстве увеличивается.
У покрытосеменных растений половое размножение связано с цветком.
Важные части цветка – это пестик и тычинка. С их участием происходят сложные процессы полового размножения –опыление и оплодотворение.
Но сперва в будущем цветке начинают образовываться половые клетки.
В жизненном цикле цветковых растений наблюдается смена поколений.
У растений выделяют диплоидное поколение – бесполое, или спорофит, и гаплоидное поколение – половое, или гаметофит.
Гаметы образуются в результате митоза, а споры – в результате мейоза. И гаметы, и споры образуются в цветке, поэтому цветок является органом и бесполого, и полового размножения.
Пыльцевое зерно является спорой (микроспорой), а не мужской гаметой, так как в нем самом развиваются мужские гаметы.
У цветковых растений мужские гаметофиты столь малы, что помещаются внутри оболочки пыльцевого зерна и состоят всего лишь из нескольких клеток.
Женский гаметофит цветковых (зародышевый мешок) помещается внутри семяпочки и состоит в наиболее распространённом случае из 7 клеток (содержит 8 – либо 7 после слияния двух ядер в центральной клетке и образования вторичного ядра).
Посмотрим, как образуется мужской гаметофит.
Итак, Микроспорогенез
В субэпидермальной ткани молодого пыльника обособляется специальная спорогенная ткань, называемая археспорием. Каждая первичная археспориальная клетка после ряда делений становится материнской клеткой пыльцы (микроспороцитом), которая проходит все фазы мейоза.
В результате двух мейотических делений возникают четыре гаплоидные микроспоры. Последние лежат четвёрками и называются клеточными тетрадами.
При созревании клеточные тетрады распадаются на отдельные микроспоры с образованием внутренней (интина) и наружной (экзина) оболочек. Наружная оболочка, как правило, грубая, поверхность её либо гладкая, либо шероховатая, приспособленная для переноса пыльцы и прилипания её к рыльцу пестика.
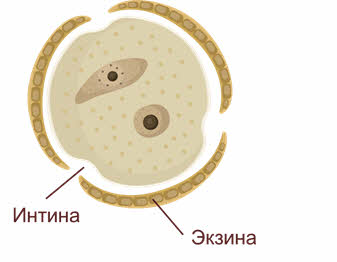
Этим заканчивается микроспорогенез, вслед за образованием одноядерной микроспоры начинается микрогаметогенез.
Первое митотическое деление микроспоры приводит к образованию вегетативной и генеративной клеток. В дальнейшем вегетативная клетка и её ядро не делятся. В ней накапливаются запасные питательные вещества, которые в последующем обеспечивают деление генеративной клетки и рост пыльцевой трубки в столбике пестика.
Генеративная клетка, содержащая меньшее количество цитоплазмы, вновь делится. Это деление может осуществляться ещё в пыльцевом зерне или в процессе его прорастания в пыльцевой трубке. В результате образуются две мужские половые клетки, которые, в отличие от сперматозоидов животных, называются спермиоклетками, или спермиями.
Таким образом, из одной споры (микроспоры) с гаплоидным набором хромосом в результате двух митотических делений образуются три ядра: два из них – спермии и одно – вегетативное. При образовании пыльцевой трубки это вегетативное ядро переходит в пыльцевую трубку.
Процесс деления генеративной клетки и образование спермиев в пыльцевой трубке были впервые подробно изучены российским и советским цитологом и эмбриологом растений Сергеем Гавриловичем Навашиным в 1910 г. на лилейных растениях.
После образования гамет пыльник созревает, и пыльца высыпается. Она несёт только генетическую информацию.
Посмотрим, как происходит мегаспорогенез и мегагаметогенез цветковых.
У покрытосеменных растений женский гаметофит – это зародышевый мешок, который закладывается и развивается внутри семяпочки.
Развитию женского гаметофита у высших покрытосеменных растений предшествует мегаспорогенез.
В субэпидермальном слое молодой семяпочки обособляется археспориальная клетка, чаще она только одна. Клетка археспория растёт, превращаясь в материнскую клетку мегаспоры.
В результате двух делений мейоза материнской клетки мегаспоры образуется тетрада мегаспор. Каждая из клеток тетрады по числу хромосом является гаплоидной. Однако только одна из них продолжает развиваться, остальные три дегенерируют, судьба этих клеток напоминает судьбу редукционных телец при созревании яйцеклеток у животных.
На следующем этапе осуществляется мегагаметогенез. Оставшаяся функционировать мегаспора продолжает расти и затем её ядро претерпевает ряд делений. При этом сама клетка не делится, а делится только ядро.
У разных систематических групп растений число делений ядра мегаспоры может варьировать от одного до трёх. У большинства растений (70 % видов покрытосеменных) этих делений, как правило, в результате возникает восемь наследственно одинаковых ядер, вовремя этих делений ядра занимают полярное положение, четыре из них оказываются лежащими ближе к микропиле (место проникновения спермиев), а четыре других – в противоположном конце зародышевого мешка, называемого халазальным. Дальше эти ядра обособляются в самостоятельные клетки, имеющие значительные количества цитоплазмы.
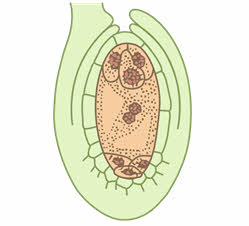
В дальнейшем от каждой из двух полярных четвёрки ядер к центру отходит по одному ядру, которые сливаются, образуя вторичное (центральное) ядро зародышевого мешка. Затем цитоплазма обособляется вокруг ядер гаметофита, который из ядерной стадии развития переходит в клеточную. Три ядра, оставшиеся вблизи халазального полюса, преобразуются в три клетки (антиподы), которые питают гаметофит. Три ядра вблизи микропиле отделяются клеточными перегородками, образуя отдельные клетки: крупную центральную яйцеклетку и две боковые клетки синергиды. Вся цитоплазма, расположенная между антиподами с одной стороны и клетками яйцевого комплекса с другой (яйцеклетка и две синергиды), называется центральной клеткой. В ней находится диплоидное вторичное (центральное) ядро.
На этом этапе женский гаметофит уже полностью сформирован и состоит из шести гаплоидных клеток (одной яйцеклетки, двух синергид и трёх антипод) и одной диплоидной (центральной). Его строение внешне напоминает мешочек, поэтому женский гаметофит покрытосеменных называется зародышевым мешком.
Пылинка попадает на рыльце пестика, и происходит опыление.
Опыление – это перенос пыльцевых зёрен на рыльце пестика, у голосеменных пыльцевые зерна при опылении попадают непосредственно на семязачаток.
Имеется два основных типа опыления: самоопыление (автогамия) (когда растение опыляется собственной пыльцой) и перекрёстное опыление (аллогамия).
При самоопылении исключён обмен генетической информацией, поскольку пыльцевые зерна попадают на рыльце пестика либо с одной из тычинок этого же цветка, либо с другого цветка, расположенного на том же растении. Это приводит к появлению чистых линий гомозиготных популяций в пределах одного вида, неспособных обмениваться мутировавшими генами, поэтому процессы видообразования в этих популяциях идут самостоятельно.
Перекрёстное опыление – это перенос пыльцы одного растения на рыльце другого. Этот тип опыления встречается более часто, чем самоопыление, между разными особями одного вида происходит обмен аллелями, что приводит к увеличению доли гетерозиготных организмов.
Безусловно, перекрёстное самоопыление имеет большие преимущества по сравнению с самоопылением, поскольку возникшие мутации свободно распространяются в пределах популяции.
Способы опыления у перекрёстноопыляемых цветковых растений весьма разнообразны. Их можно разделить на две группы. Первая: перенос пыльцы осуществляется главным образом насекомыми, а также некоторыми позвоночными (птицами и летучими мышами). Соответственно, различают энтомофилию, орнитофилию и зоофилию. Растения, опыляемые животными, обычно имеют яркоокрашенные крупные цветки. Мелкие цветки, как правило, собраны в соцветия, что зрительно их увеличивает. Для привлечения опылителей служит нектар или большое количество пыльцы, которую опылители охотно поедают.
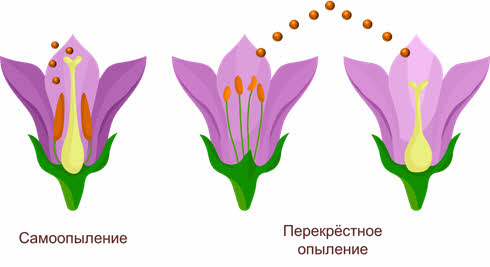
Вторая группа: пыльца переносится абиотическими факторами – ветром и реже водой, в связи с чем различают анемофилию и гидрофилию.
После попадания пыльцы на рыльце начинается прорастание пыльцевого зерна.
Экзина мужского гаметофита прорывается в области борозды или поры прорастания, и начинает формироваться пыльцевая трубка, которая растёт, проникает в рыльце, через столбик движется по направлению к завязи, где находится семязачаток.
Трубка растёт на кончике. В растущую пыльцевую трубку из пыльцевого зерна перемещается ядро клетки-трубки, а также спермин.
Обычно развитие пыльцевой трубки происходит при уже сформированных семязачатках в завязи. Достигнув завязи, пыльцевая трубка через микропиле проникает внутрь одного из находящихся там семязачатков.
Там пыльцевая трубка направляется к яйцевому аппарату, проникает в одну из синергид и освобождает спермин. Один из спермиев сливается с яйцеклеткой, в результате чего образуется зигота.
Другой спермий сливается с полярными ядрами центральной клетки, образуя триплоидное ядро (с тройным набором хромосом). Зигота даёт начало зародышу, а из триплоидной центральной клетки образуется ткань эндосперма.
Таким образом оплодотворение, при котором одна мужская гамета сливается с яйцеклеткой, а вторая ― с вторичным ядром, называется двойным оплодотворением.
Механизм оплодотворения цветковых –двойное оплодотворение, открыто отечественным учёным Сергеем Гавриловичем Навашиным в 1898 году.
Таким образом, оплодотворённый семязачаток развивается в семя, из зиготы возникает зародыш, из триплоидной центральной клетки – эндосперм, а из внешней части семязачатка (интегументов) образуется семенная кожура, из стенок завязи цветка – стенки плода.
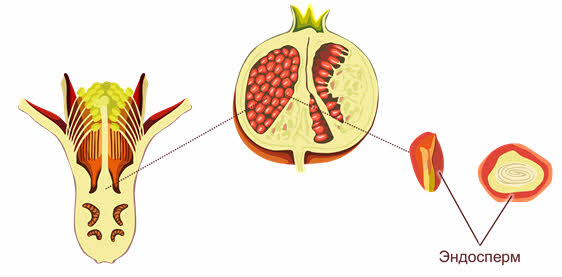
Эндосперм цветковых полностью отличается от первичного эндосперма голосеменных. У них первичный эндосперм представляет собой гаплоидную вегетативную ткань женского гаметофита, где накапливаются питательные вещества семени.
Эндосперм у покрытосеменных происходит из триплоидной центральной клетки, в образовании которой участвуют вторичное ядро женского гаметофита и ядро спермия.
У одних цветковых (например, злаков) эндосперм сильно разрастается и занимает большую часть семени, оттесняя на периферию маленький зародыш.
У других (к пример у бобовых) весь эндосперм поглощается зародышем и используется его семядолями, которые становятся самыми большими структурами зрелого семени.
Читайте также: