
Молекулярно биохимические методы изучения эволюции кратко
Обновлено: 05.07.2024
Прогресс эволюционной биохимии неотделим от поисков новых методов исследования.
Представление о биохимии древнейших форм жизни в настоящее время базируется преимущественно на данных сравнительной биохимии современных организмов. Палеобиохимия сделала первые самостоятельные шаги в 20‑е годы XX в. (Я.В. Самойлов). Изучение органического состава ископаемых костей, раковин, углеродсодержащих осадочных пород приводит к выводу о глубокой древности многих веществ (хлорофилл, аминокислоты;. На протяжении длительных геологических периодов оставались неизменными некоторые белки. Для познания биохимии ископаемых организмов большие возможности открывают изотопные измерения. Методами химического анализа были обнаружены и определены аминокислоты в остатках динозавра, жившего в меловой период (П. Эйбелсон), в коже амфибий пермского периода (Т.В. Дроздова), в скелете рыб из олигоценовых отложении (Т.В. Дроздова, А.В. Коченов), у палеозойских граптолитов (С.М. Майская, Т.В. Дроздова) и пектин у ископаемых организмов, найденных в горючих сланцах (А.П. Виноградов, Е.А. Бойченко).
Сравнительная биохимия широко использовала методы серодиагностики. Реакция преципитации использовалась для выяснения филогенетических отношений как в ботанике, так и в зоологии. На основании данных серодиагностики создавались даже целостные филогенетические системы примером чему может служить система растительного мира К. Меца (см. главу 2). Несмотря на критику, серологические методы продолжают сохранять известное значение в сравнительно‑филогенетических исследованиях.
Крепнет убеждение, согласно которому основные биохимические системы сформировались на очень ранних этапах эволюции. Дж. Бернал, например, предлагает ограничить проблему физико‑химического биопоэза стадией клетки, ограниченной мембраной, а дальнейшее исследование процесса эволюции проводить в рамках старых морфологических дисциплин, ибо эволюция простейших и высших организмов не была связана, по‑видимому, с появлением принципиально новых биохимических механизмов. Аналогичной точки зрения придерживался Н. Пири (1959), по мысли которого достижение морфологической сложности кладет предел усложнению биохимической структуры.

Методы геносистематики быстро прогрессируют. Было обнаружено, что определенное таксономическое значение могут иметь наличие, природа и характер распределения минорных метилированных оснований в ДНК, а также нуклеотидная последовательность ДНК[173]. Сравнительное изучение последней оказывается незаменимым подспорьем в тех случаях, когда нуклеотидный состав ДНК далеко отстоящих форм случайно совпадает (например, по составу ДНК представители губок, моллюсков, иглокожих и хордовых почти неотличимы, а представители кишечнополостных, моллюсков, ракообразных и хордовых – практически идентичны). Получены данные, позволяющие с большой долей вероятности предполагать существование прямой зависимости между степенью сблоченности пиримидинов в ДНК и высотой организации живых форм. Должны быть также упомянуты попытки использовать в таксономических целях состав и количество сателлитных ДНК, существующих в виде примеси к суммарной ДНК организмов. В этом направлении сделаны только первые шаги. Большие перспективы открывает перед таксономией метод молекулярной гибридизации, позволяющий определять величину сходных нуклеотидных последовательностей, по которой судят о родстве организмов.
Развитие эволюционной биохимии не создает конфликта с эволюционной морфологией. Изучение закономерностей развития органического мира средствами морфологии не утрачивает своего значения, как это иногда представлялось в 30‑40‑е годы XX в. (Дж. Холдейн, М. Флоркэн). Существует и крепнет тенденция комплексного изучения эволюции данной биологической структуры или системы с использованием методов морфологии, биохимии и физиологии. Опыт морфологии оказывается полезен для эволюционной биохимии. В этой связи заслуживают упоминания попытки истолкования процессов биохимической эволюции в понятиях эволюционной морфологии. Так, Г.Ф. Гаузе и Д.И. Сапожников обращались к морфобиологической теории эволюции А.Н. Северцова. Согласно Гаузе (1959), появление в ходе биохимической эволюции механизмов, которые препятствовали рацемизации протоплазмы и обеспечивали ее оптическую активность, явилось ароморфозом, обеспечившим повышение интенсивности химических процессов в организме. Ароморфозом считают также обогащение набора ферментов, вызванное ассимиляцией первичными существами органических веществ биогенного происхождения (Д.И. Сапожников, 1959). Обосновывалась приложимость к биохимической эволюции закона Л. Долло о необратимости эволюции (М. Флоркэн, 1944) и понятия брадителической эволюции, введенного Г. Симпсоном (А.В. Благовещенский, 1966).
Развитие биологии в последарвиновский период дает много убедительных примеров эвристической ценности морфологического подхода к изучению сложнейших закономерностей эволюции органического мира. Возможности эволюционной морфологии все еще далеко не исчерпаны. Вместе с тем становится все очевиднее, что представление о процессе эволюции, чтобы стать более удовлетворительным, должно охватывать физиологический, биохимический и биофизический аспекты эволюции живого и опираться на знание эволюции элементарных, основополагающих процессов в организме. С каждым новым успехом физиологии, биохимии и молекулярной биологии настоятельнее становилась потребность в специальных исследованиях, направленных на познание функциональных и биохимических закономерностей эволюции. Распространение исторического метода из области морфологии в область генетики, экспериментальной эмбриологии, гистологии, физиологии, биохимии и молекулярной биологии, а также синтез сравнительного, экспериментального и исторического методов составляют характерные черты развития биологии в XX в. Глубоко ошибочно противопоставление теории эволюции молекулярной биологии и теории систем, получающей все большее распространение в биологии. На протяжении XX в. исторический метод не оставался неизменным.
Умелая конкретизация метода применительно к данному уровню организации живого, требовавшая всякий раз творческого решения сложных теоретических вопросов, способствовала достижению новых результатов.
Чем глубже исследовательская мысль проникает в сущность жизни, в тонкую организацию живого, тем настоятельнее испытывает биология нужду в синтетическом направлении, способном создать целостную картину эволюционного процесса. Тенденция к интеграции эволюционных направлений возникших в разных областях биологии – морфологии, экологии, генетике, физиологии, биохимии, гистологии, экспериментальной эмбриологии, молекулярной биологии и других – до сих пор развивавшихся в значительной мере изолированно друг от друга, – характерная особенность прогресса всего комплекса биологических наук в XX веке. Усилиями биологов разных специальностей постепенно создается единое здание эволюционной биологии, фундаментом которому продолжает служить дарвинизм.
Глава 22
Прогресс эволюционной биохимии неотделим от поисков новых методов исследования.
Представление о биохимии древнейших форм жизни в настоящее время базируется преимущественно на данных сравнительной биохимии современных организмов. Палеобиохимия сделала первые самостоятельные шаги в 20‑е годы XX в. (Я.В. Самойлов). Изучение органического состава ископаемых костей, раковин, углеродсодержащих осадочных пород приводит к выводу о глубокой древности многих веществ (хлорофилл, аминокислоты;. На протяжении длительных геологических периодов оставались неизменными некоторые белки. Для познания биохимии ископаемых организмов большие возможности открывают изотопные измерения. Методами химического анализа были обнаружены и определены аминокислоты в остатках динозавра, жившего в меловой период (П. Эйбелсон), в коже амфибий пермского периода (Т.В. Дроздова), в скелете рыб из олигоценовых отложении (Т.В. Дроздова, А.В. Коченов), у палеозойских граптолитов (С.М. Майская, Т.В. Дроздова) и пектин у ископаемых организмов, найденных в горючих сланцах (А.П. Виноградов, Е.А. Бойченко).
Сравнительная биохимия широко использовала методы серодиагностики. Реакция преципитации использовалась для выяснения филогенетических отношений как в ботанике, так и в зоологии. На основании данных серодиагностики создавались даже целостные филогенетические системы примером чему может служить система растительного мира К. Меца (см. главу 2). Несмотря на критику, серологические методы продолжают сохранять известное значение в сравнительно‑филогенетических исследованиях.
Крепнет убеждение, согласно которому основные биохимические системы сформировались на очень ранних этапах эволюции. Дж. Бернал, например, предлагает ограничить проблему физико‑химического биопоэза стадией клетки, ограниченной мембраной, а дальнейшее исследование процесса эволюции проводить в рамках старых морфологических дисциплин, ибо эволюция простейших и высших организмов не была связана, по‑видимому, с появлением принципиально новых биохимических механизмов. Аналогичной точки зрения придерживался Н. Пири (1959), по мысли которого достижение морфологической сложности кладет предел усложнению биохимической структуры.
Методы геносистематики быстро прогрессируют. Было обнаружено, что определенное таксономическое значение могут иметь наличие, природа и характер распределения минорных метилированных оснований в ДНК, а также нуклеотидная последовательность ДНК[173]. Сравнительное изучение последней оказывается незаменимым подспорьем в тех случаях, когда нуклеотидный состав ДНК далеко отстоящих форм случайно совпадает (например, по составу ДНК представители губок, моллюсков, иглокожих и хордовых почти неотличимы, а представители кишечнополостных, моллюсков, ракообразных и хордовых – практически идентичны). Получены данные, позволяющие с большой долей вероятности предполагать существование прямой зависимости между степенью сблоченности пиримидинов в ДНК и высотой организации живых форм. Должны быть также упомянуты попытки использовать в таксономических целях состав и количество сателлитных ДНК, существующих в виде примеси к суммарной ДНК организмов. В этом направлении сделаны только первые шаги. Большие перспективы открывает перед таксономией метод молекулярной гибридизации, позволяющий определять величину сходных нуклеотидных последовательностей, по которой судят о родстве организмов.
Развитие эволюционной биохимии не создает конфликта с эволюционной морфологией. Изучение закономерностей развития органического мира средствами морфологии не утрачивает своего значения, как это иногда представлялось в 30‑40‑е годы XX в. (Дж. Холдейн, М. Флоркэн). Существует и крепнет тенденция комплексного изучения эволюции данной биологической структуры или системы с использованием методов морфологии, биохимии и физиологии. Опыт морфологии оказывается полезен для эволюционной биохимии. В этой связи заслуживают упоминания попытки истолкования процессов биохимической эволюции в понятиях эволюционной морфологии. Так, Г.Ф. Гаузе и Д.И. Сапожников обращались к морфобиологической теории эволюции А.Н. Северцова. Согласно Гаузе (1959), появление в ходе биохимической эволюции механизмов, которые препятствовали рацемизации протоплазмы и обеспечивали ее оптическую активность, явилось ароморфозом, обеспечившим повышение интенсивности химических процессов в организме. Ароморфозом считают также обогащение набора ферментов, вызванное ассимиляцией первичными существами органических веществ биогенного происхождения (Д.И. Сапожников, 1959). Обосновывалась приложимость к биохимической эволюции закона Л. Долло о необратимости эволюции (М. Флоркэн, 1944) и понятия брадителической эволюции, введенного Г. Симпсоном (А.В. Благовещенский, 1966).
Развитие биологии в последарвиновский период дает много убедительных примеров эвристической ценности морфологического подхода к изучению сложнейших закономерностей эволюции органического мира. Возможности эволюционной морфологии все еще далеко не исчерпаны. Вместе с тем становится все очевиднее, что представление о процессе эволюции, чтобы стать более удовлетворительным, должно охватывать физиологический, биохимический и биофизический аспекты эволюции живого и опираться на знание эволюции элементарных, основополагающих процессов в организме. С каждым новым успехом физиологии, биохимии и молекулярной биологии настоятельнее становилась потребность в специальных исследованиях, направленных на познание функциональных и биохимических закономерностей эволюции. Распространение исторического метода из области морфологии в область генетики, экспериментальной эмбриологии, гистологии, физиологии, биохимии и молекулярной биологии, а также синтез сравнительного, экспериментального и исторического методов составляют характерные черты развития биологии в XX в. Глубоко ошибочно противопоставление теории эволюции молекулярной биологии и теории систем, получающей все большее распространение в биологии. На протяжении XX в. исторический метод не оставался неизменным.
Умелая конкретизация метода применительно к данному уровню организации живого, требовавшая всякий раз творческого решения сложных теоретических вопросов, способствовала достижению новых результатов.
Чем глубже исследовательская мысль проникает в сущность жизни, в тонкую организацию живого, тем настоятельнее испытывает биология нужду в синтетическом направлении, способном создать целостную картину эволюционного процесса. Тенденция к интеграции эволюционных направлений возникших в разных областях биологии – морфологии, экологии, генетике, физиологии, биохимии, гистологии, экспериментальной эмбриологии, молекулярной биологии и других – до сих пор развивавшихся в значительной мере изолированно друг от друга, – характерная особенность прогресса всего комплекса биологических наук в XX веке. Усилиями биологов разных специальностей постепенно создается единое здание эволюционной биологии, фундаментом которому продолжает служить дарвинизм.
Рассмотрим главнейшие из методов изучения эволюционного процесса, представляемых биологическими дисциплинами в последовательности, которая отражает проникновение эволюционных идей в эти дисциплины: сначала палеонтологические, биогеографические, морфологические, эмбриологические и систематические, а затем данные генетики, биохимии, молекулярной биологии.
Палеонтологические методы. По существу, все без исключения методы палеонтологии как науки об ископаемых организмах могут рассматриваться как методы изучения эволюционного процесса. Рассмотрим более подробно лишь главнейшие палеонтологические методы изучения эволюции: выявление ископаемых промежуточных форм, восстановление филогенетических рядов и обнаружение последовательности ископаемых форм.
Ископаемые переходные формы – формы организмов, сочетающие признаки более древних и молодых групп. Поиски и детальные описания таких форм служат важными методами восстановления филогенеза отдельных групп.
Переходными формами от рептилий к птицам являются юрские первоптицы Archaeopteryx с длинным, как у рептилии, хвостом, несросшимися позвонками и брюшными ребрами, развитыми зубами. Но это были уже настоящие птицы: тело покрыто хорошо развитыми перьями, передние конечности превращены в типичные крылья.
Палеонтологические ряды – ряды ископаемых форм, связанных друг с другом в процессе эволюции и отражающие ход филогенеза. По числу найденных промежуточных звеньев такой ряд может быть более или менее подробным, но во всех случаях это должен быть именно ряд форм, близких не только основными, но и частными деталями строения и, несомненно, генеалогически связанных друг с другом в процессе эволюции.
Биогеографические методы. Биогеография дает в руки исследователей методы, позволяющие проанализировать общий ход эволюционного процесса в самых разных масштабах.
Островные формы. Фауна и флора островов оказывается тем более своеобразной, чем глубже и дольше эти острова были изолированы от основной суши. Так, например, сравнительно недавно потерявшие связь с материком Британские острова имеют фауну с небольшим числом автохтонных (возникших именно здесь) видов. К последним относятся куропатка-граус (Lagopus scoticus), два вида полевок, несколько уже упоминавшихся пресноводных сигов, ряд улиток и некоторое число видов мелких насекомых. С другой стороны, давно обособившийся (десятки миллионов лет) от африканского материка остров Мадагаскар имеет фауну настолько своеобразную, что выделяется в особую зоогеографическую подобласть: из 36 родов млекопитающих 32 рода эндемичны; из 127 родов птиц эндемична почти половина.
Реликты. О флоре и фауне далекого прошлого Земли свидетельствуют и реликтовые формы. Реликты – отдельные виды или небольшие группы видов с комплексом признаков, характерных для давно вымерших групп прошлых эпох. Среди животных одной из наиболее ярких реликтовых форм является гаттерия (Sphenodon punctatus) – единственный представитель целого подкласса рептилий. В ней отражены черты рептилий, живших на Земле десятки миллионов лет назад – в мезозое (рис. 6.13). Гаттерия живет в глубоких норах на островах залива Пленти (Новая Зеландия). Другой известный реликт – кистеперая рыба латимерия (Latimeria chalumnae), сохранившаяся малоизмененной с девона в глубоководных участках прибрежных вод Восточной Африки.
Морфологические методы. Использование морфологических (сравнительно-анатомических, гистологических и др.) методов изучения эволюции основано на принципе: глубокое внутреннее сходство организмов может показать родство сравниваемых форм.
Гомология органов. Органы с общим планом строения, развивающиеся из сходных зачатков, находящиеся в сходном соотношении с другими органами и выполняющие как сходные, так и различные функции, называются гомологичными. Другими словами, это органы, имеющие глубокое сходство в строении, основанное на единстве происхождения соответствующих организмов.
У человека атавизмами являются хвост, мощный волосяной покров на поверхности тела, наличие не двух, а нескольких пар сосков и т.п. Развитие у лошади вместо грифельных косточек (рудиментарных пальцев) настоящих боковых пальцев, так же как и развитие у китообразных выступающих на поверхность тела задних конечностей,– примеры атавизмов. Отличие рудиментарных органов от атавизмов состоит в том, что первые встречаются у всех членов данной популяции, вторые – лишь у немногих особей.
Рудиментарные органы и атавизмы – убедительные доказательства процесса эволюции.
Эмбриологические методы. Эмбриология располагает целым арсеналом методов изучения эволюционного процесса. Среди них главные: выявление зародышевого сходства и изучение рекапитуляции.
Выявление зародышевого сходства. В первой половине XIX в. выдающийся натуралист К. Бэр сформулировал «закон зародышевого сходства: чем более ранние стадии индивидуального развития исследуются, тем больше сходства обнаруживается между различными организмами. Например, на ранних стадиях развития эмбрионы позвоночных не отличаются друг от друга. Лишь на средних стадиях развития в сравниваемом ряду у зародышей появляются особенности, характерные для рыб и амфибий; на ее более поздних стадиях – особенности рептилий, птиц и млекопитающих
Принцип рекапитуляции отнюдь не ограничивается лишь морфологическими изменениями. В процессе эволюции позвоночных происходит постепенная утрата ферментов, необходимых для распада мочевой кислоты (конечного продукта обмена пуринов). Так, у некоторых рептилий и птиц конечный продукт такого обмена – мочевая кислота, у земноводных и большинства рыб – мочевина, у беспозвоночных – аммиак. Эмбриобиохимические исследования показали, что зародыш птиц на ранних стадиях развития выделяет аммиак, на более поздних – мочевину, а на последних стадиях развития – мочевую кислоту. Сходным образом у головастиков – личинок бесхвостых амфибий – конечный продукт обмена – аммиак, а у взрослых лягушек – мочевина.
Эти факты показывают на действие принципа рекапитуляции и среди физиолого-биохимических признаков.
Генетические методы изучения эволюции разнообразны. Это и прямое определение генетической совместимости сравниваемых форм (например, посредством гибридизации), и анализ цитогенетических особенностей организмов. Изучением повторных инверсий в определенных хромосомах у разных популяций одного и того же или близких видов можно с большой точностью восстановить последовательность возникновения таких инверсий, т.е. восстановить микрофилогенез таких групп (рис. 6.27). Анализ числа и особенностей строения хромосом в группах близких видов часто позволяет выявлять направления возможной эволюции генома таких форм, т.е. выяснять их эволюционные взаимоотношения. На рис. 6.28 приведены хромосомные наборы ряда близких форм, которые могли возникнуть лишь в процессе последовательной полиплоидизации генома исходного типа, а в табл. 6.4– примеры рядов полиплоидных видов у некоторых растений.
Выяснение строения нуклеиновых кислот и белков. На молекулярном уровне процесс эволюции связан с изменением состава нуклеотидов (в ДНК и РНК) и аминокислот (в белках). На современном этапе развития молекулярной биологии можно анализировать число различий в последовательностях элементов нуклеиновой кислоты или белка разных видов, судить по этому показателю о степени их отличий. Поскольку каждая замена аминокислоты в белке может быть связана с изменением одного, двух или трех нуклеотидов в молекуле ДНК, компьютерными методами можно вычислить максимальное и минимальное число нуклеотидных замен, необходимых для замещения аминокислот в белке.
Биохимический полиморфизм. Начиная с 60-x годов в практику микроэволюционных исследований широко входит метод изучения вариаций белков, обнаруживаемых с помощью электрофореза (Р. Левонтин). С помощью этого метода можно с большей точностью, чем прежде, определять уровень генетической изменчивости в популяциях, а также степень сходства и различия между популяциями по сумме таких вариаций. При биохимических сравнениях ныне живущих организмов показано, что ранние метаболические звенья синтеза стеринов, каротиноидов, вторичных метаболитов, реакции гликолиза и дыхания у многих растений и животных сходны и не нуждаются в участии кислорода. Завершающие же стадии многих метаболических путей, особенно у высокоорганизованных существ, протекают с его участием. Это свидетельствует об их эволюционной молодости и сложении в аэробной фазе истории Земли. Таким образом, с помощью биохимических методов (и в том числе молекулярно-биологических) можно исследовать эволюционные процессы самого разного масштаба. Несомненно, что в ближайшие годы арсенал этих методов должен существенно расшириться, учитывая значительное развитие исследований в этой области.
Выяснение строения нуклеиновых кислот и белков. На молекулярном уровне процесс эволюции связан с изменением состава нуклеотидов (в ДНК и РНК) и аминокислот (в белках). На современном этапе развития молекулярной биологии можно анализировать число различий в последовательностях элементов нуклеиновой кислоты или белка разных видов, судить по этому показателю о степени их отличий. Поскольку каждая замена аминокислоты в белке может быть связана с изменением одного, двух или трех нуклеотидов в молекуле ДНК, компьютерными методами можно вычислить максимальное и минимальное число нуклеотидных замен, необходимых для замещения аминокислот в белке.
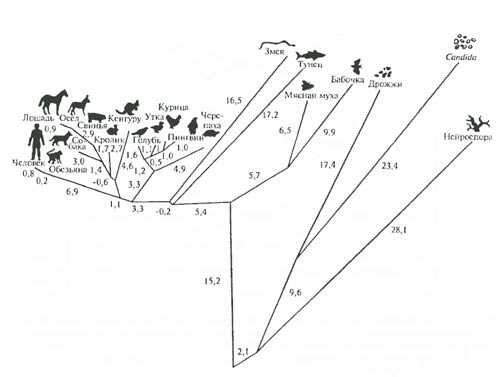
Рис. 6.29. Филогенетические дистанции между разными организмами, построенные на основании количественных изменений молекулярной структуры белка цитохрома C. Цифры – число мутаций (по А.Ф. Айяла, 1981)
Получаемая таким образом информация поддается дальнейшей количественной оценке: при сравнении ряда организмов можно установить и степень различий (меру эволюционной дивергенции) макромолекул. Другое важное преимущество изучения эволюции методами молекулярной биологии – возможность сравнения сколь угодно далеких организмов – растений и животных, грибов и микроорганизмов. На рис. 6.29 показан результат изучения филогенетических отношений 20 разных организмов на основе определения минимального числа возможных нуклеотидных различий между генами, кодирующими синтез белка цитохрома C. Можно видеть, что в общем эти данные хорошо совпадают с выводами классической систематики. Сейчас построены многие сотни филогенетических деревьев макромолекул. При общем принципиальном совпадении с классическими эволюционными схемами эволюции крупных групп, молекулярными методами оказалось возможным вскрыть немало важных особенностей. Так, при анализе РНК, кодируемой не только ядерными генами, но и клеточными органеллами у растений, выяснилось, например (К. Вууз), что ядерная фракция p-РНК кукурузы (Zea mays) относится к ветви эукариот, а фракции из митохондрий и хлоропластов – к ветви эубактерий. Этот факт является мощным аргументом в пользу симбиотического происхождения эукариот: возникновения митохондрий от пурпурных бактерий, а хлоропластов – от цианобактерий (см. рис. 5.3). При реконструкции древа глобинов (В.А. Ратнер) удалось показать, что средняя скорость эволюции этих белков у животных заметно возрастала 400–500 млн. лет назад (период выхода позвоночных на сушу), когда глобин позвоночных приобрел тетрамерную структуру. Построение филогенетического древа генов, кодирующих синтез гемагглютининов H3 вируса гриппа показало, что скорость эволюции эпидемических вариантов этого вируса (испанка 1918–1919 гг., гонконгский грипп в середине – второй половине XX в. и др.) в несколько раз выше скорости эволюции неэпидемических штаммов. Аналогичные построения для вируса ВИЧ показали, что этот вирус очень близок к одному из вирусов обезьян. Он существовал в Центральной Африке до 1960 г., появился на о. Гаити в середине 70-х гг. и в США к 1978 г. Он распространяется ныне благодаря способности изменяться с невероятной скоростью – увеличивает агрессивность даже в теле одного человека на протяжении 1,5–2 лет.
Молекулярный филогенетический анализ сегодня стал одним из главнейших методов выяснения филогенетических взаимоотношений самых разных групп живых существ и помогает решать немало спорных вопросов макроэволюции.
Изучение особенностей эволюции на молекулярном уровне привело к идее молекулярных часов, не только отражающих, но и регулирующих эволюционный процесс. Такие расчеты основаны на предположениях о накоплении изменений в информационных макромолекулах с постоянной скоростью. Однако оказалось, что для растений, отличающихся в целом исключительно высокой скоростью эволюции первичной структуры ДНК, концепция молекулярных часов неприемлема: ДНК растений из разных семейств отличается в такой же степени, как ДНК животных разных классов. У растений в ходе эволюции очень быстро могут меняться и повторяющиеся, и уникальные последовательности нуклеотидов в молекулах ДНК. Скорость эволюции макромолекул в разных филогенетических линиях растений оказывается далеко не одинаковой. Так, темпы накопления изменений в нуклеотидных последовательностях у линий, ведущих к цветковым, более чем в 10 раз выше, чем в линии высших нецветковых. При изучении генов актинов было выявлено, что различия между близкими видами у растений превышают таковые разных классов у животных. Поэтому скорость хода молекулярных часов оказывается резко различной в разных группах, а также, по-видимому, в разные геологические эпохи (см. также гл. 15, 20).
Ответ. Это свидетельствует о едином происхождении всего живого. Все клетки способны к самостоятельному существованию, самовоспроизведению и развитию. Несмотря на многообразие форм, все живые клетки организмов состоят из одних и тех же химических веществ, подчинены единым структурным принципам. Каждый из компонентов выполняет свою особую функцию, а в совокупности все они определяют жизнедеятельность клетки в целом. Реализуются все признаки и свойства клеток и организмов благодаря белкам, структуру которых в первую очередь и определяют последовательности нуклеотидов ДНК. Поэтому первостепенное значение в процессах метаболизма играет именно биосинтез нуклеиновых кислот и белка. Структурной единицей наследственной информации является ген.
Вопрос 1. В чём заключается сущность молекулярно-биохимических методов изучения эволюционного процесса? Приведите примеры таких методов.
Ответ. Методы биохимии и молекулярной генетики изучают строение белков и нуклеиновых кислот организмов, относящихся к разным семействам, отрядам, классам. По степени различий в строении белков и нуклеотидов можно установить степень филогенетического родства различных таксонов. Методы биохимии и молекулярной генетики изучают строение белков и нуклеиновых кислот организмов, относящихся к разным семействам, отрядам, классам. По степени различий в строении белков и нуклеотидов можно установить степень филогенетического родства различных таксонов. Химический анализ клеток различных организмов показал, что все они имеют сходный элементарный состав. В них встречаются одни и те же белки, углеводы, липиды и нуклеиновые кислоты. В состав белков всех организмов входят 20 обязательных аминокислот. У представителей разных систематических групп встречаются сходные по строению, составу и функциям белки. Так, сходное строение имеют хлорофилл растений и гемоглобин животных. Идентичное строение имеют многие гормоны позвоночных животных. Например, гормоны гипофиза найдены у всех групп позвоночных. Гормон инсулин имеется у всех млекопитающих.
Изучение аминокислотной последовательности белков различных организмов позволяет установить их эволюционное родство. Наглядный пример тому – родство белков миоглобина и гемоглобина.
Большое сходство наблюдается у различных организмов и в процессах обмена веществ. Например, у зародышей птиц на ранних стадиях эмбрионального развития в качестве конечного продукта белкового обмена образуется аммиак, на более поздних стадиях – мочевина, а у взрослых организмов – мочевая кислота. У головастиков лягушки белковый обмен также заканчивается образованием аммиака, что свойственно рыбам, а у взрослых земноводных конечный продукт – мочевина.
Универсальность генетического кода доказывает единство всех организмов на Земле. Изучение первичной структуры ДНК показало, что у многих организмов имеются ряды сходных последовательностей нуклеотидов. У близкородственных видов это сходство очень велико. Например, анализ структуры митохондриальной ДНК гориллы, орангутанга и человека позволил построить родословную гоминид. Оказалось, что митохондриальный ген гориллы отличается от аналогичного гена человека на 10%, а орангутана – на 17%. Вероятно, эти различия обусловлены мутациями, которые возникали у каждого вида в процессе эволюции.
Вопрос 2. Какие данные молекулярной биологии и биохимии доказывают филогенетическое родство организмов? Приведите примеры биохимической гомологии.
Ответ. Данные молекулярной биологии. Носителем наследственной информации во всех клетках являются молекулы ДНК, у всех известных организмов в основе размножения – репликация этой молекулы. В ДНК всех организмов используются 4 нуклеотида (аденин, гуанин, тимин, цитозин), хотя в природе встречаются не менее 102 различных нуклеотидов. Кроме того, в природе встречается 390 различных аминокислот, но белки всех организмов составляются из одного и того же набора, в котором всего 22 аминокислоты.
Данные биохимии. Также можно отметить крайне похожие пути метаболизма в клетках всех организмов. Так, гликолиз у всех эукариот и у большинства прокариот проходит за 10 одинаковых шагов, в одной и той же последовательности, с использованием одних и тех же десяти ферментов (при том что возможны тысячи различных, но термодинамически эквивалентных путей гликолиза). У всех изученных видов основным переносчиком энергии в клетке является аденозинтрифосфат (АТФ), хотя эту роль могли бы выполнять сотни других молекул.
Пример биохимической гомологии – факт, что у позвоночных имеются сходные или идентичные гормоны, выполняющие целый ряд различных функций. Например, выделяемый гипофизом гормон, сходный с пролактином млекопитающих, найден у представителей всех групп позвоночных. По имеющимся данным, пролактин вызывает множество разнообразных эффектов, которые можно разбить на две большие группы: одни эффекты связаны с размножением, а другие – с осморегуляцией
Таким образом, общность химического состава клеток разных организмов указывает на биохимическую гомологию, подобную морфологической, что свидетельствует в пользу эволюции.
Вопрос 3. С какой целью учёные используют метод компьютерного моделирования хода эволюционного процесса? Что доказывает такое моделирование?
Так, отечественными учёными в Институте эволюционной физиологии и биохимии РАН была составлена компьютерная программа, в которую заложили описание прототипа хордовых животных, аналогичного дожившему до наших дней ланцетнику.
Этот исходный вид мог преобразовываться в другие виды организмов. Эволюционные усложнения и упрощения были равновероятными, исключались из филогенетического процесса только крупные скачки, например внезапное возникновение хорошо развитого органа – головного мозга, глаза и т. п.
Читайте также: