
Какова роль клеток в процессах онтогенеза кратко
Обновлено: 02.07.2024
Проблема реализации генотипа в фенотип, в виде клеточных и системных процессов остается актуальной на сегодняшний день. Ученые-биологи стремятся выяснить степень и конкретные пути контроля со стороны генома и одновременно уровень автономности различных процессов в ходе онтогенеза.
В ходе индивидуального развития активно реализуются клеточные механизмы онтогенеза, к которым можно отнести:
Клеточное деление – сложная функция организма, подчиняющаяся регулирующим влияниям на различных уровнях его организации: генетическом, тканевом, онтогенетическом.
Деление клеток играет важную роль в процессах онтогенеза:
Во-первых – благодаря делению зиготы, возникает многоклеточный организм.
Во-вторых – пролиферация клеток, происходящая в стадии дробления обеспечивает рост организма.
В-третьих – избирательному размножению клеток принадлежит заметная роль в обеспечении морфогенетических процессов. В постнатальном периоде индивидуального развития, благодаря клеточному делению осуществляется обновление тканей, а также восстановление утраченных органов, заживление ран.
В эмбриогенезе, в периоде дробления, клетки делятся быстрее, чем в другие поздние периоды. Во время гаструляции и органогенеза клетки делятся избирательно в определенных областях зародыша. В сформировавшемся организме разные клетки делятся с разной скоростью или вообще не делятся, например, нейроны, а эпителиальные и кроветворные клетки, наоборот, активно размножаются.
В эмбриональном периоде обнаружено, что делящиеся клетки группируются гнездами. Однако, само деление не придает эмбриональному зачатку форму, в результате миграции и перераспределения клеток зачаток приобретает форму.
За последние годы установлено, что многие структуры зародыша образуются клетками, происходящими от небольшого числа или даже одной клетки. Совокупность клеток, являющихся потомками одной родоначальной клетки, называют клоном. Показано, например, что большие по объему участки центральной нервной системы формируются из определенных клеток раннего зародыша. Пока не ясно, в какой именно срок происходит отбор родоначальных клеток, каков механизм этого отбора. Важным следствием такой селекции является то, что многим клеткам раннего зародыша не суждено участвовать в дальнейшем развитии. В опытах на мышах показано, что организм развивается всего из трех клеток внутренней клеточной массы на стадии, когда бластоциста состоит из 64 клеток, а сама внутренняя клеточная масса содержит примерно 15 клеток. Клональные клетки могут быть причиной мозаицизма, когда большие группы клеток отличаются по набору хромосом или аллельному составу.
Таким образом, деление клеток является чрезвычайно важным процессом в онтогенетическом развитии. Оно протекает с разной интенсивностью в разное время и в разных местах, носит клональный характер и подвержено генетическому контролю.
Миграции клеток, (клеточные перемещения), наряду с другими клеточными процессами имеют очень большое значение, начиная с процесса гаструляции и далее, в процессах морфогенеза. Клетки мезенхимного типа мигрируют одиночно и группами, а клетки эпителиев обычно согласованно, пластом. Мезенхима – это скопление веретеновидных или звездчатых клеток, погруженных в межклеточный матрикс. Эпителий – группы клеток, плотно прилежащих друг к другу боковыми стенками и имеющих апикальную и базальную поверхности. Как мезенхима, так и эпителии могут быть образованы из любого из трех зародышевых листков. Клетки мезенхимного типа наиболее подвижны, так как не образуют между собой стойких контактов. Нарушение, миграции клеток в ходе эмбриогенеза приводит к недоразвитию органов или к их гетеротопиям, изменениям нормальной локализации. То и другое представляет собой врожденные пороки развития.
Существуют гипотезы о дистантных воздействиях на клетки на основе хемотаксиса и о контактных воздействиях. Мезенхимные клетки способны к амебоидным движениям. Движение их по типу хемотаксиса показано для некоторых видов специализированных клеток (гоноциты, сперматозоиды, некоторые клетки крови). Контактные взаимодействия более распространены. Они представляют собой взаимодействие клеток со структурированным субстратом. Клетки ощущают микроструктуру субстрата и движутся вдоль волокон, как это происходит при помещении фибробластов на каплю кровяной плазмы, натянутой в виде пленки между сторонами стеклянного треугольника. На неструктурированной подложке, например на ненатянутой пленке, фибробласты перемещаются беспорядочно.
Таким образом, для миграции клеток очень важны их способность к амебоидному движению и свойства клеточных мембран. И то, и другое генетически детерминировано, так что и сама миграция клеток находится под генетическим контролем, с одной стороны, и влияниями окружающих клеток и тканей – с другой.
В процессе эмбриогенеза клетки образуют скопления и пласты только с определенными клетками. Значительные координированные перемещения клеток характерны для периода гаструляции. Смысл этих перемещений заключается в образовании обособленных друг от друга зародышевых листков с совершенно определенным взаимным расположением. Клетки как бы сортируются в зависимости от свойств.
Необходимым условием сортировки являются степень подвижности клеток и особенности их мембран. В поздней бластуле амфибий, например, клетки будущей эктодермы обладают тенденцией слипаться друг с другом и распространяться в виде сплошного слоя над мезодермой и энтодермой. Эта тенденция проявляется и в культуре тканей. Клетки мезодермы имеют тенденцию впячиваться в любой находящийся поблизости комок клеток, а клетки энтодермы относительно неподвижны.
Эти особенности трудно объяснить, если наблюдать только за нормальным течением развития. Впервые на различия в подвижности и способности к слипанию (адгезии) между клетками, относящимися к разным зародышевым листкам, обратил внимание Г.Гольтфретер в 30-х гг. XX столетия. Затем производили модификации его опытов с самыми разными клетками. Смысл этих опытов состоит в том, что зародыши тритонов или других животных на стадии гаструлы диссоциируют с помощью фермента трипсина, который разрушает материал, соединяющий клетки друг с другом. Диссоциированные (дезагрегированные) клетки тщательно перемешивают и затем создают такие условия, чтобы клетки могли свободно перемещаться и воссоединяться друг с другом. Сначала клетки представляют собой беспорядочную смесь, затем клетки эктодермы, мезодермы и энтодермы разделяются (сегрегируют) и собираются в отдельные группы, каждая из которых представляет собой клеточный агрегат из однородных клеток. Образуются снова зародышевые листки, располагающиеся иногда даже в обычном для них порядке.
Подобную агрегацию клеток зародышевого листка с себе подобными можно объяснить способностью к избирательному слипанию клеток одного типа между собой. Одновременно это является проявлением ранней дифференцировки клеток на стадии гаструлы.
Существует ряд гипотез, объясняющих избирательную сортировку клеток. Возможно, что контакты между подобными клетками сильнее, чем между чужеродными клетками, из-за различий в поверхностном заряде их мембран. Обнаружено, что поверхностный заряд клеток мезодермы ниже, чем клеток экто- или энтодермы, благодаря чему клетки мезодермы легче деформируются и втягиваются в бластопор в начале гаструляции. По другой гипотезе, контактные взаимодействия между одинаковыми клетками основываются на антигенных свойствах их мембран.
Избирательная адгезия клеток определенного зародышевого листка друг с другом является необходимым условием нормального развития. Это показано путем сопоставления двух процессов: течения гаструляции в естественных условиях и способности клеток определенного зародышевого листка агрегировать в комплекс с себе подобными. Показано, что если у межвидовых гибридов зародышей амфибий нарушена гаструляция, то диссоциированные в опыте клетки зародышевых листков таких гибридов не способны к агрегации с себе подобными, если же в некоторых случаях гибридам удавалось образовать гаструлу, то и в эксперименте диссоциированные клетки зародышевых листков были способны к реагрегации. По-видимому, в обеспечении сортировки клеток важное место принадлежит генетическим механизмам. Одним из примеров потери клетками способности к избирательной сортировке и слипанию является их беспорядочное поведение в злокачественной опухоли.
Таким образом, сортировка клеток и их избирательная адгезия наряду с другими клеточными процессами играет важную роль в морфогенезе развивающегося зародыша и одновременно подвержена многоуровневым регуляционным воздействиям, отражая целостность организма как системы.
Эмбриональный гистогенез. Пролиферация клеток. Клеточный рост, миграция и межклеточные взаимодействия.
В развитии низших и высших позвоночных отчетливо прослеживается единая общебиологическая закономерность, выражающаяся в появлении зародышевых листков и обособлении основных зачатков органов и тканей. Процесс образования тканей из материала эмбриональных зачатков составляет суть учения о гистогенезах.
Эмбриональный гистогенез, по определению А.А. Клишова (1984), — это комплекс координированных во времени и пространстве процессов пролиферации, клеточного роста, миграции, межклеточных взаимодействий, дифференциации, детерминации, программированной гибели клеток и некоторых других. Все названные процессы в той или иной мере протекают в зародыше, начиная с самых ранних стадий его развития.
Пролиферация. Основной способ деления тканевых клеток — это митоз. По мере увеличения числа клеток возникают клеточные группы, или популяции, объединенные общностью локализации в составе зародышевых листков (эмбриональных зачатков) и обладающие сходными гистогенетическими потенциями. Клеточный цикл регулируется многочисленными вне- и внутриклеточными механизмами. К внеклеточным относятся влияния на клетку цитокинов, факторов роста, гормональных и нейрогенных стимулов. Роль внутриклеточных регуляторов играют специфические белки цитоплазмы. В течение каждого клеточного цикла существуют несколько критических точек, соответствующих переходу клетки из одного периода цикла в другой. При нарушении внутренней системы контроля клетка под влиянием собственных факторов регуляции элиминируется апоптозом, либо на некоторое время задерживается в одном из периодов цикла.
Метод радиографического анализа клеточных циклов в различных тканях выявил особенности соотношения клеточной репродукции и дифференцировки. Например, если в тканях (кроветворные ткани, эпидермис) имеется постоянный фонд пролиферирующих клеток, за счет которых обеспечивается непрерывное возникновение новых клеток взамен погибающих, то эти ткани относятся к обновляющимся. Другие ткани, например, некоторые соединительные, характеризуются тем, что в них увеличение количества клеток происходит параллельно с их дифференцировкой, клетки в этих тканях характеризуются низкой митотической активностью. Это растущие ткани. Наконец, нервная ткань характеризуется тем, что все основные процессы репродукции заканчиваются в период эмбрионального гистогенеза (когда формируется основной запас стволовых клеток, достаточный для последующего развития ткани). Поэтому она отнесена к стабильным (стационарным) тканям. Продолжительность жизни клеток в обновляющихся, растущих и стабильных тканях разная.
Наряду с обновлением клеточной популяции, в самих клетках постоянно наблюдается обновление внутриклеточных структур (внутриклеточная физиологическая регенерация).
Клеточный рост, миграция и межклеточные взаимодействия. Рост клеток проявляется в изменении их размеров и формы. При усилении функциональной активности и внутриклеточных биосинтезов наблюдается увеличение объема клетки. Если объем клетки превышает некую норму, то говорят о ее гипертрофии, и наоборот, при снижении функциональной активности происходит уменьшение объема клетки, а при переходе некоторых нормативных параметров возникает атрофия клетки. Рост клетки не беспределен и определяется оптимальным ядерно-цитоплазменным отношением.
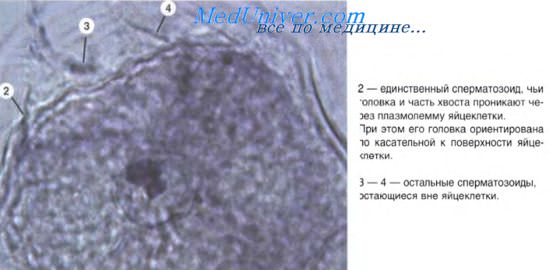
Важное значение для гистогенеза имеют процессы перемещения клеток. Миграция клеток наиболее характерна для периода гаструляции. Однако и в период гисто- и органогенеза происходят перемещения клеточных масс (например, смещения миобластов из миотомов в места закладки скелетных мышц; движение клеток из нервного гребня с образованием спинномозговых ганглиев и нервных сплетений, миграция гоноцитов и т. д.). Миграция осуществляется с помощью нескольких механизмов. Так, различают хемотаксис — движение клеток в направлении градиента концентрации какого-либо химического агента (перемещения спермиев к яйцеклетке, предшественников Т-лимфоцитов из костного мозга в закладку тимуса).
Гаптотаксис — механизм перемещения клеток по градиенту концентрации адгезионной молекулы (движение клеток протока пронефроса у амфибий по градиенту щелочной фосфатазы на поверхности мезодермы). Контактное ориентирование — когда в какой-либо преграде остается один канал для перемещения (описан у рыб при образовании плавников).
Контактное ингибирование — этот способ перемещения наблюдается у клеток нервного гребня. Суть способа заключается в том, что при образовании ламеллоподии одной клеткой и контакта ее с другой клеткой, ламеллоподия прекращает рост и постепенно исчезает, но в другой части мигрирующей клетки при этом формируется новая ламеллоподия.
В процессе миграции клеток важную роль играют межклеточные взаимодействия. Существует несколько механизмов такого взаимодействия (контактного и дистантного). Выделяется большая группа молекул клеточной адгезии (МКА). Так, кадгерины — это Са2+-зависимые МКА, отвечают за межклеточные контакты при образовании тканей, за формообразование и др. В молекуле кадгерина различают внеклеточный, трансмембранный и внутриклеточный домены. Например, внеклеточный домен ответственен за адгезию клеток с одинаковыми кадгеринами, а внутриклеточный — за форму клетки. Другой класс МКА — это иммуноглобулиновое суперсемейство Са2+-независимых МКА, обеспечивающих, например, адгезию аксонов к сарколемме мышечных волокон, или миграцию нейробластов вдоль радиальных глиоцитов в закладке коры большого мозга и др. Следующий класс МКА — это мембранные ферменты — гликозилтранферазы. Последние по типу "ключ-замок" соединяются с углеводными субстратами — гликозаминогликанами надмембранного комплекса клетки, осуществляя таким образом прочное сцепление клеток.
Кроме механизмов межклеточного взаимодействия, существуют механизмы взаимодействия клеток с субстратом. Они включают формирование рецепторов клетки к молекулам внеклеточного матрикса. К последним относят производные клеток, среди которых наиболее изученными адгезионными молекулами являются коллаген, фибронектин, ламинин, тенасцин и некоторые другие. Коллагены, среди которых различают несколько десятков типов, входят в состав межклеточного вещества рыхлой волокнистой соединительной ткани, базальной мембраны и пр. Фибронектин, секретируемый клетками, является связывающей молекулой между мигрирующей клеткой и межклеточным матриксом. Ламинин — компонент базальной мембраны, также связывает мигрирующие клетки с межклеточным матриксом (справедливо по отношению к эпителиоцитам и нейробластам).
Для осуществления связи мигрирующих клеток с межклеточным матриксом клетки формируют специфические рецепторы. К ним относятся, например, синдекан, который обеспечивает контакт эпителиоцита с базальной мембраной за счет сцепления с молекулами фибронектина и коллагена. Интегрины клеточных поверхностей связывают с внеклеточной стороны молекулы внеклеточного матрикса, а внутри клетки — белки цитоскелета (например, актиновые микрофиламенты). Так возникает связь внутри- и внеклеточных структур, что позволяет клетке использовать для перемещения собственный сократительный аппарат. Наконец, существует большая группа молекул, формирующих клеточные контакты, осуществляющие коммуникацию между клетками (щелевые контакты), механическую связь (десмосомы, плотные контакты).
Дистантные межклеточные взаимодействия осуществляются путем секреции гормонов и факторов роста (ФР). Последние — это вещества, оказывающие стимулирующее влияние на пролиферацию и дифференцировку клеток и тканей. К ним относятся, например, ФР, полученный из тромбоцитов и влияющий на переход клеток в фазу размножения (гладких миоцитов, фибробластов, глиоцитов); эпидермальный ФР — стимулирует пролиферацию эпителиоцитов, производных эктодермы; ФР фибробластов — стимулирует пролиферацию фибробластов. Особо выделяется большая группа пептидов (соматотропины, соматомедины, инсулин, лактоген), влияющих на развитие клеток плода.
Факторы, тормозящие пролиферацию и дифференцировку клеток, также принимают кооперативное участие в процессах развития клеток и тканей. К ним относятся, например, бета-интерферон и трансформирующий ФР. Последний, однако, в отношении разных клеточных типов действует по-разному: блокирует размножение многих типов эпителиоцитов, но стимулирует размножение соединительнотканных клеток.
- Вернуться в оглавление раздела "гистология"
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Эмбриональное (зародышевое) развитие охватывает процессы от первого деления зиготы до выхода из яйца или рождения и у большинства животных включает три основных этапа: дробление, гаструляцию и органогенез.
При дроблении дочерние клетки ( бластомеры ) не расходятся и не увеличиваются в размерах. С каждым следующим делением их размеры уменьшаются.

Яйцеклетки с небольшим запасом питательных веществ делятся полностью, т. е. происходит полное дробление. Если яйцеклетка содержит большое количество желтка, то наблюдается частичное дробление — делится только диск цитоплазмы с ядром, а сам желток остаётся без изменений (например, у птиц).
Бластула — это шарообразный зародыш, стенка которого ( бластодерма ) образована одним слоем клеток, а внутри — полость ( бластоцель ).
После дробления начинается гаструляция — часть клеток бластодермы перемещается внутрь зародыша. В результате этих перемещений образуется гаструла.
Гаструла — двухслойный зародыш, состоящий из двух зародышевых листков: наружного ( эктодермы ) и внутреннего ( энтодермы ).
У ланцетника образование гаструлы происходит в результате впячивания части бластодермы внутрь бластоцеля.
Внутренняя полость гаструлы называется первичной кишкой . Её связывает с внешней средой отверстие ( бластопор ), которое становится первичным ртом .

На стадии гаструлы (двух зародышевых листков) прекращается развитие двухслойных животных — губок и кишечнополостных.
У всех остальных животных развитие продолжается, и образуется третий зародышевый листок — мезодерма . Она формируется из энтодермы и всегда расположена между экто- и энтодермой в первичной полости тела.
Дальнейшая специализация клеток зародышевых листков обеспечивает формирование тканей и органов, т. е. гисто- и органогенез .
Из энтодермы образуется хорда — внутренний скелет в виде гибкого тяжа, расположенный на спинной стороне. Позже вместо хорды у позвоночных развивается позвоночник, и только у некоторых животных (например, у хрящевых рыб) её остатки сохраняются в течение всей жизни.
Из эктодермы, расположенной над самой хордой, выделяется нервная пластинка . Затем края пластинки поднимаются и смыкаются. Образуется нервная трубка — зачаток центральной нервной системы. Формируется нейрула .

Нервная трубка, хорда и кишечник создают осевой комплекс органов зародыша, который определяет двустороннюю симметрию тела.

Из эктодермы у позвоночных животных образуется нервная система, органы чувств, покровный эпителий с его железами и производными структурами (волосы, перья, копыта, когти и т. п.).
Из энтодермы формируются органы пищеварительной и дыхательной системы: эпителий средней кишки, печень и поджелудочная железа, жабры, лёгкие, плавательный пузырь, а также щитовидная железа.
Из мезодермы формируются все виды мышечной и соединительной ткани (например, дерма кожи, тела позвонков), кровеносная система, органы выделения, половые железы.
Раздел ЕГЭ: 3.3. Онтогенез и присущие ему закономерности. Эмбриональное и постэмбриональное развитие организмов. Причины нарушения развития организмов
Онтогенез (индивидуальное развитие) включает весь период жизни особи от зиготы до смерти. Различают два периода: эмбриональный и постэмбриональный.

Эмбриональный период
Эмбриональный период (зародышевое развитие) — период онтогенеза от образования зиготы в процессе оплодотворения до рождения или выхода из яйцевых оболочек.
Этапы эмбрионального периода
В процессе дробления зигота многократно делится, при этом размеры дочерних клеток-бластомеров становятся всё меньше, так как после деления они не растут. В результате дробления образуется однослойный многоклеточный зародыш — бластула. Он напоминает полый шар с эпителиальной стенкой (бластодермой) и центральной полостью (бластоцелью, или первичной полостью тела), которая заполнена жидкостью.
После дробления наступает процесс гаструляции, который характеризуется перемещением части клеточного материала с поверхности внутрь, на места будущих органов. В результате образуется гаструла. Гаструла — чашевидный зародыш, состоящий из двух слоёв (зародышевые листки): наружный (эктодерма) и внутренний (энтодерма). На этой стадии заканчивается развитие низших животных. Нейрула — стадия закладывания внутренних органов (полости первичной кишки, нервной пластинки, хорды в случае хордовых животных). На этом этапе происходит образование третьего зародышевого листка — мезодермы, которая формируется в виде карманов — целом (вторичная полость тела).
Органогенез — дифференцировка клеток каждого зародышевого листка. В процессе органогенеза образуются ткани и органы.
Из эктодермы формируются нервная система, органы чувств, эпителий кожи с его железами и производными структурами (волосы, перья, когти и др.), эмаль зубов. Производными мезодермы являются все виды соединительной и мышечная ткани, кровеносная, выделительная и половая системы. Из энтодермы образуются эпителий средней кишки и его придаточные железы (печень, поджелудочная железа), жабры и их производные — лёгкие, плавательный пузырь и др., а также щитовидная железа.

Органогенез у хордовых связан возникновением комплекса осевых органов: нервной трубки, хорды и кишечника — и дальнейшей их дифференцировкой.
Постэмбриональный период
Постэмбриональный период развития начинается рождением или выходом из яйцевых оболочек и заканчивается смертью организма.
Постэмбриональное развитие начинается с выхода новой особи из яйцевых оболочек или (при живорождении) из организма матери. Оно подразделяется на три периода — ювенильный, пубертатный и период старения.
Этапы постэмбрионального периода
1) Ювенильный, продолжается до окончания полового созревания. Развитие организма в этот период может протекать по двум различным путям. Прямое развитие происходит, если из яйца или из организма матери выходит особь, похожая на взрослую, но меньшая по размерам и с несформированной половой системой. Другой тип развития называется непрямым и проходит с метаморфозом. Ювенильный период практически всегда сопровождается ростом организма. С одной стороны, процесс роста запрограммирован генетически, а с другой — зависит от условий существования. У человека рост контролируется целым рядом гормонов, выделяемых гипоталамусом, гипофизом, щитовидной и половыми железами.
2) Пубертатный (т. е. период зрелости). У большинства позвоночных животных он занимает, как правило, большую часть жизни.
3) Старение — это общебиологическая закономерность, свойственная живым организмам. В определенном для каждого вида возрасте в организме начинаются изменения, снижающие возможности этого организма к приспособлению к изменяющимся условиям существования.
Смерть — это прекращение жизнедеятельности организма. Однако смерть необходима для эволюционного процесса. Без смерти не происходила бы смена поколений — одна из основных движущих сил эволюции.
Типы постэмбрионального развития
- Прямое: появившийся на свет организм имеет все основные органы, свойственные взрослому, и в дальнейшем происходит только его рост и половое созревание (рептилии, птицы, млекопитающие).
- Непрямое: эмбриональное развитие приводит к образованию личинки, отличающейся по ряду анатомических и морфологических признаков от взрослой особи (имаго у насекомых), часто ведёт иной образ жизни (лягушки, некоторые насекомые, черви).

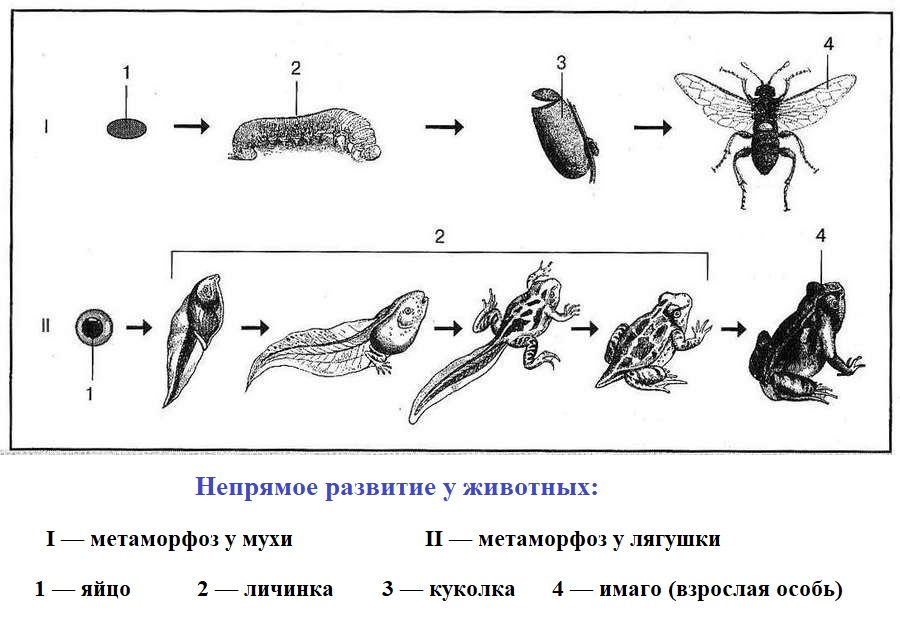
- Ограниченный (закрытый): происходит в определённые периоды онтогенеза, в основном до полового созревания (насекомые, птицы, млекопитающие).
- Неограниченный (открытый): продолжается в течение всего онтогенеза, вплоть до смерти (моллюски, рыбы, земноводные, пресмыкающиеся).

Читайте также: