
Генетические процессы популяции кратко
Обновлено: 05.07.2024
Как уже говорилось, наследуются не сами признаки, а код наследственной информации, определяющий комплекс возможностей развития — норму реакции генотипа, в пределах которой возможно не приводящее к гибели взаимодействие развивающейся особи со средой. При этом наследственная реализация каждого признака или свойства определяется не одним, а, как правило, очень многими генами (принцип полимерии в действии генов); с другой стороны, любой ген оказывает влияние не на один, а на многие признаки (принцип плейотропии в действии гена). Границы характерной для данного генотипа нормы реакции могут быть выражены, таким образом, лишь совокупностью фенотипов, развившихся из этого генотипа при всех возможных условиях среды.
Частоты генов, генотипов и фенотипов. В основе генетической изменчивости популяций лежат закономерности наследования признаков, определяемые механизмом распределения хромосом и генов в мейозе и случайным сочетанием гамет при оплодотворении.
Сейчас известно, что все природные популяции гетерогенны, они насыщены мутациями. Генетическая гетерогенность любой популяции при отсутствии давления внешних факторов должна быть неизменной, находиться в определенном равновесии.
Рассмотрим два упрощенных примера.
Предположим, что в популяции число форм, гомозиготных по разным аллелям одного гена (AA и aa), одинаково. Если особи — носители данных аллелей — равновероятно свободно скрещиваются друг с другом (панмиксия), то возможны следующие комбинации:
| Самки | Самцы | |
| 0,5A | 0,5a | |
| 0,5A | 0,25AA | 0,25Aa |
| 0,5a | 0,25Aa | 0,25aa |
Цифры показывают, что в данном поколении в популяции будут возникать гомозиготы AA и aa с частотой 0,25, а гетерозиго-ты Aa — с частотой 0,50. Это же соотношение сохранится и в следующем поколении: частота гамет с рецессивным аллелем а составит 0,5 (0,25 от гомозигот aa + 0,25 от гетерозигот Aa), так же как и частота гамет с доминантным аллелем А (0,25 от гомозигот AA + 0,25 от гетерозигот Aa). Это же соотношение сохранится во всех следующих поколениях, если не будет нарушено каким-либо внешним давлением.
Конечно, в подавляющем большинстве случаев в популяции встречается разное число гомозигот AA и aa. Разберем пример, когда частота аллелей данного гена в популяции будет 0,7a, 0,3A:
| Самки | Самцы | |
| 0,3A | 0,7a | |
| 0,3A | 0,09AA | 0,21Aa |
| 0,7a | 0,21Aa | 0,49aa |
Таким образом, в потомстве на 100 зигот будет 9 гомозигот AA, 49 гомозигот aa и 42 гетерозиготы Aa. В следующем поколении гаметы с аллелем А будут возникать с частотой 0,3 (0,09 от гомозигот AA + 0,21 от гетерозигот Aa), а гаметы с аллелем а будут возникать опять-таки с частотой 0,7 (0,49 от гомозигот aa + 0,21 от гетерозигот Aa). Как и в первом примере, без внешнего давления это соотношение сохранится в каждой последующей генерации.
Если частоту встречаемости одного аллеля данного гена определить как о, то частота альтернативного аллеля того же гена может быть обозначена как 1—q. В потомстве свободно скрещивающихся особей должны быть следующие отношения таких аллелей:
Это при суммировании дает q 2 +2q( 1—q) + (1—q) 2 , или [q + (1—q)] 2
Эта формула называется формулой Харди — Вайнберга и позволяет рассчитывать относительную частоту генотипов и фенотипов в популяции. Например, предположим, что в популяции каких-либо жуков обнаружены красные формы с частотой 25% (или 0,25) и черные с частотой 75% (или 0,75); черный цвет определяется доминантным аллелем А, а красный — рецессивным аллелем а. При этом частота встречаемости генотипов aa составит (1 —q) 2 = 0,25, а частота встречаемости аллеля a будет √(1—q) 2 = √0,25 =0,5.
Согласно той же формуле, частота доминантного аллеля А составит 1 — 0,5 = 0,5, а частота гомозиготных доминантных генотипов AA составит в популяции q 2 = 0,5 2 = 0,25.
Формула Харди — Вайнберга пригодна лишь для предельно упрощенной ситуации, для идеальной бесконечно большой популяции и при отсутствии давления каких-либо факторов. Кроме того, как известно, на частоту фенотипа оказывает влияние не только частота данного аллеля, но и такие его свойства, как доминантность, рецессивность, пенетрантность и экспрессивность. Следовательно, при анализе природных популяций эта формула применима лишь с большими оговорками.
При анализе природных популяций необходимо различать понятия частота гена (количественное соотношение аллелей одного какого-либо локуса), частота генотипа (количественное соотношение разных генотипов) и частота фенотипа (количественное соотношение разных фенотипов).
Генотипы особей одной популяции сходны, но все же не идентичны. Наибольший интерес представляет собой наличие в популяции тех или иных аллелей генов.
Гены, для которых имеются несколько аллелей, называются полиморфными (схема 1).

Схема 1. Аллели – разные формы гена А: А, А* и А**
Для одного и того же гена набор аллелей в разных популяциях может отличаться.
Кроме того, одни и те же аллели могут быть распределены среди особей по-разному, т. е. быть в гомо- или гетерозиготном состоянии. Количество аллелей и количество типов распределения аллелей определяет генетическое разнообразие популяции.
От уровня генетического разнообразия зависит интенсивность эволюционных процессов и стабильность генотипа популяции.
Количество аллелей и соотношение гомо- и гетерозигот определяет генетическое разнообразие популяции.

Схема 2. Разные варианты комбинации трех аллелей гена А.
От уровня генетического разнообразия зависит интенсивность эволюционных процессов.
Закономерности генетических процессов в популяции можно изучать только при условии свободного скрещивания. Т. е. такого скрещивания, при котором вероятность соединения любых гамет и комбинации их признаков одинакова.
Изучая генетические процессы в естественных популяциях, английский ученый Пирсон (рис. 1) в 1904 году вывел закон стабилизирующего скрещивания, или закон Пирсона.

Он может быть сформулирован так: при любом исходном соотношении частот гомозигот и гетерозигот при первом скрещивании внутри популяции устанавливается состояние равновесия, если исходные частоты аллелей одинаковы у обоих полов.
Конкретный вид этого равновесия можно описать с помощью элементарной математической формулы, которую независимо вывели английский математик Дж. Харди и немецкий врач и биолог В. Вайнберг (рис. 2).

Рис. 2. Дж. Харди и В. Вайнберг
Закон Харди – Вайнберга гласит, что частота гомозиготных и гетерозиготных организмов в условия свободного скрещивания при отсутствии давления отбора и других факторов пребывает в состоянии равновесия.
В простейшем виде закон описывается следующей формулой (схема 3):

Схема 3. Закон Харди – Вайнберга
Практическая значимость закона Харди – Вайнберга
Расчёт частот аллелей, по закону Харди – Вайнберга, имеет большое значение для медицины. Медики рассчитывают частоту возникновения генетических заболеваний. Величина человеческой популяции достаточно велика, чтобы можно было опираться на вычисления с помощью этого закона. Также этот закон используется селекционерами, для определения сроков закрепления нужных признаков.
Популяционные волны
Популяционные волны возникают с определенной регулярностью и имеют значительную амплитуду. Так частота популяционных волн у зайцев составляет около 10 лет, бабочки белянки – 10–12 лет, саранчи – 11 лет. При этом популяция зайцев в Канаде на пике популяционной волны увеличивается в 10 раз, а майского жука в 1 миллион раз (рис. 3).

Рис. 3. Частота всплесков численности различных травоядных животных. Сегодня эти циклы связывают с периодами повышенной солнечной активности, которые, в свою очередь, влияют на продуктивность растений – кормовой базы этих животных.
Факторы нарушающие генетическую стабильность популяции
Важным следствием этого закона является постоянство частот встречаемости каждой аллели в популяции.
Необходимо отметить, что закон Харди – Вайнберга, как и другие генетические закономерности, основанные на менделевском принципе случайного комбинирования, точно выполняется только при бесконечно большой численности популяции.
На практике это означает, что в небольших популяциях закон Харди – Вайнберга выполняется нечетко.
Русский ученый С.С. Четвериков также отмечал, что в результате свободного скрещивания происходит постоянное поддержание равновесия генотипических частот в популяции.
Нарушение равновесия связано, как правило, с действием внешних сил и наблюдается только до тех пор, пока эти силы оказывают влияние.
Остановимся на этих силах подробнее. К факторам, нарушающим генетическую стабильность популяции, т. е. увеличивающим генетическое разнообразие, относятся миграции, изоляции, дрейф генов, популяционные волны и естественный отбор (схема 4).

Схема 4. Факторы, нарушающие генетическую стабильность популяции
Миграции и изоляции приводят к резкому изменению генофонда, поскольку происходит изменение частоты встречаемости генотипов за счет добавления новых или исключения старых.
Дрейф генов – это явление случайного статистического отклонения в частотах аллелей, в небольших популяциях.
Закон Харди – Вайнберга четко соблюдается только для бесконечно больших популяций.
Чем меньше популяция, тем больше величина отклонения, т. е. дрейфа генов. В результате дрейфа генов, в маленьких популяциях может происходить даже полное элиминирование некоторых аллелей гена (схема 5).

Схема 5. Популяционные волны и дрейф генов
Как видно из графика, в пятидесятом поколении произошло полное выпадение носителей одного из аллелей.
Популяционные волны – это резкие изменения численности популяции, связанные с действиями внешней среды (наличием пищи, хищников или абиотических факторов). Например, увеличение численности лисиц приводит к падению численности зайцев. Но затем это приводит к падению численности самих лисиц, лишившихся основного пищевого ресурса (схема 6).

Схема 6. Популяционные волны хищников и их жертв на примере долговременных наблюдений за лисами и зайцами.
Изменение численности сразу сказывается на частоте встречаемости аллелей. Кроме того, популяционные волны обычно сопровождаются обострением борьбы за выживание и стимуляцией естественного отбора.
Естественный отбор, в свою очередь, приводит к отбору некоторых аллельных вариантов, соответствующих наиболее приспособленному фенотипу. Таким образом, естественный отбор приводит не к случайному образованию гамет, а к выборочному.
Перечисленные факторы, приводящие к изменению частот аллелей в популяциях, приводят к накоплению различий между этими популяциями, и в дальнейшем способствуют видообразованию.
В настоящее время известно, что все природные популяции гетерогенны и насыщены мутациями. Генетическая гетерогенность любой популяции при отсутствии давления внешних факторов должна быть неизменной, находиться в определенном равновесии. А. В. Яблоков, А. Г. Юсупов (1998) приводят расчеты на двух примерах, впервые сделанные Г. Харди (1908).
Предположим, что в популяции число форм гомозиготных по разным аллелям одного гена (АА и аа) одинаково. Если особи — носители данных аллелей совершенно свободно скрещиваются друг с другом, то возможны следующие комбинации:
| Самки | Самцы | |
| 0,5А | 0,5а | |
| 0,5А 0,5а | 0.25АА 0,25Аа | 0,25Аа 0,25аа |
Цифры показывают, что в данном поколении в популяции будут возникать гомозиготы АА и аа с частотой по 0,25, а гетерозиготы Аа — с частотой 0,50. Это же соотношение сохранится и в следующем поколении: частота гамет с рецессивным аллелем а составит 0,5 (0,25 от гомозигот аа+0,25 от гетеро-зигот Аа), также как и частота гамет с доминантным аллелем А (0,25 от гомозигот АА+0,25 от гетерозигот Аа). Это же соотношение сохранится во всех следующих поколениях, если не будет нарушено каким-либо внешним давлением.
Определенно, в подавляющем большинстве случаев в популяции встречается разное число гомозигот АА и аа. Разберем пример, когда частота аллелей данного гена в популяции будет 0,7 а, 0,ЗА:
| Самки | Самцы | |
| 0,ЗА | 0,7а | |
| 0,ЗА 0,7а | 0.09АА 0,21Аа | 0,21Аа 0,49аа |
Следовательно, в потомстве на 100 зигот будет 9 гомозигот АА, 49 гомозигот аа и 42 гетерозиготы Аа. В следующем поколении гаметы с аллелем А будут возникать с частотой 0,3 (0,09 от гомозигот АА+0,21 от гетерозигот Аа), а гаметы с аллелем а будут возникать опять-таки с частотой 0,7 (0,49 от гомозигот аа+0,21 от гетерозигот Аа). Как и в первом примере, это соотношение сохранится в каждой последующей генерации.
Если частоту встречаемости одного аллеля данного гена определить как q, то частота альтернативного аллеля того же гена может быть определена как 1—q. В потомстве свободно скрещивающихся особей должны быть следующие отношения таких аллелей:
| Самки | Самцы | |
| q | (1-q) | |
| q (1 - q) | q×q q× (1 - q) | q×(l - q) (1 - q)×(1 - q) |
При суммировании это дает:
Эта формула носит название формулы Харди—Вайнберга и позволяет рассчитывать относительную частоту генотипов и феноти пов в популяции. Так, предположим, что в популяции каких-либо жуков обнаружены красные формы с частотой 25% (или 0,25) и черные с частотой 75% (или 0,75); черный цвет определяется доминантным аллелем А, а красный — рецессивным аллелем а. При этом частота встречаемости генотипов аа составит (1 —q) 2 = 0,2 5, а частота встречаемости аллеля а составит Согласно той же формуле частота доминантного аллеля А составит 1—0,5 = 0,5, а частота гомозиготных доминантных генотипов АА составит в популяции q 2 = 0,5 2 = 0,25.
Итак, при анализе природных совокупностей особей (популяций) необходимо различать понятия: частота гена (количественное соотношение аллелей одного какого-либо локуса), частота генотипа (количественное соотношение разных генотипов) и частота фенотипа (количественное соотношение разных фенотипов).
Существование двух (или более) генетически различных форм в популяции в состоянии длительного равновесия в таких соотношениях, что частоту даже наиболее редкой формы нельзя объяснить только повторными мутациями, называется полиморфизмом. В качестве примера полиморфизма можно привести три формы цветков у примулы (Primula vulgaris), рис. 9.8.

Рис. 9.8. Три формы цветков у примулы (Primula vulgaris),
из Ф. Шеппарда, 1970
В природных популяциях примулы (Primula vulgaris) всегда есть особи с длинным пестиком и короткими пыльниками (А), длинными пыльниками и коротким пестиком (Б) и одинаковыми по длине пыльниками и пестиком (В). Самоопыление возможно только у цветов типа В. Гетеростилия способствует перекрестному оплодотворению.
Полиморфизм по механизму возникновения и поддержания разделяется на две большие группы: гетерозиготный и адаптационный.
Гетерозиготный полиморфизм устанавливается в результате давления на популяцию естественного отбора, положительно отбирающего гетерозигот.
Адаптационный полиморфизм — это две или несколько генетически различных форм внутри популяции, подвергающихся положительному отбору в разных экологических условиях.
В качестве примера можно привести адаптационный полиморфизм в популяции двухточечной тлевой (божьей) коровки Adalia bipunctata (рис. 9.9).
Рис. 9.9. Адаптивный полиморфизм в популяции тлевых
(божьих) коровок Adalia bipunctata:
I — процентное содержание черной и красной форм при весеннем (В) и осеннем (О) сборах; II — частота доминантного гена А (черная окраска) в популяциях весной и осенью каждого года (в%) (по Н. В. Тимофееву-Ресовскому и Ю. М. Свирежеву, 1965)
На протяжении 10 лет в изучении популяции тлевой коровки осенью — при уходе на зимовку — черных форм было от 50 до 70%, а весной — при выходе из зимовки — от 30 до 45%. Красных форм осенью было меньше 50%, а весной — больше. Красные формы, как было установлено, лучше переносят в зимний период холод, а черные — интенсивнее размножаются летом. Отсюда отбор направлен на сохранение большего числа красных жуков зимой и черных — летом. Разнонаправленное давление отбора в отдельные периоды жизни популяции способствует выработке устойчивого адаптационного полиморфизма.
Положение о генетическом единстве популяции является одним из наиболее важных выводов популяционной генетики: любая популяция представляет сложную генетическую систему, находящуюся в динамическом равновесии.
В настоящее время известно, что все природные популяции гетерогенны и насыщены мутациями. Генетическая гетерогенность любой популяции при отсутствии давления внешних факторов должна быть неизменной, находиться в определенном равновесии. А. В. Яблоков, А. Г. Юсупов (1998) приводят расчеты на двух примерах, впервые сделанные Г. Харди (1908).
Предположим, что в популяции число форм гомозиготных по разным аллелям одного гена (АА и аа) одинаково. Если особи — носители данных аллелей совершенно свободно скрещиваются друг с другом, то возможны следующие комбинации:
| Самки | Самцы | |
| 0,5А | 0,5а | |
| 0,5А 0,5а | 0.25АА 0,25Аа | 0,25Аа 0,25аа |
Цифры показывают, что в данном поколении в популяции будут возникать гомозиготы АА и аа с частотой по 0,25, а гетерозиготы Аа — с частотой 0,50. Это же соотношение сохранится и в следующем поколении: частота гамет с рецессивным аллелем а составит 0,5 (0,25 от гомозигот аа+0,25 от гетеро-зигот Аа), также как и частота гамет с доминантным аллелем А (0,25 от гомозигот АА+0,25 от гетерозигот Аа). Это же соотношение сохранится во всех следующих поколениях, если не будет нарушено каким-либо внешним давлением.
Определенно, в подавляющем большинстве случаев в популяции встречается разное число гомозигот АА и аа. Разберем пример, когда частота аллелей данного гена в популяции будет 0,7 а, 0,ЗА:
| Самки | Самцы | |
| 0,ЗА | 0,7а | |
| 0,ЗА 0,7а | 0.09АА 0,21Аа | 0,21Аа 0,49аа |
Следовательно, в потомстве на 100 зигот будет 9 гомозигот АА, 49 гомозигот аа и 42 гетерозиготы Аа. В следующем поколении гаметы с аллелем А будут возникать с частотой 0,3 (0,09 от гомозигот АА+0,21 от гетерозигот Аа), а гаметы с аллелем а будут возникать опять-таки с частотой 0,7 (0,49 от гомозигот аа+0,21 от гетерозигот Аа). Как и в первом примере, это соотношение сохранится в каждой последующей генерации.
Если частоту встречаемости одного аллеля данного гена определить как q, то частота альтернативного аллеля того же гена может быть определена как 1—q. В потомстве свободно скрещивающихся особей должны быть следующие отношения таких аллелей:
| Самки | Самцы | |
| q | (1-q) | |
| q (1 - q) | q×q q× (1 - q) | q×(l - q) (1 - q)×(1 - q) |
При суммировании это дает:
Эта формула носит название формулы Харди—Вайнберга и позволяет рассчитывать относительную частоту генотипов и феноти пов в популяции. Так, предположим, что в популяции каких-либо жуков обнаружены красные формы с частотой 25% (или 0,25) и черные с частотой 75% (или 0,75); черный цвет определяется доминантным аллелем А, а красный — рецессивным аллелем а. При этом частота встречаемости генотипов аа составит (1 —q) 2 = 0,2 5, а частота встречаемости аллеля а составит Согласно той же формуле частота доминантного аллеля А составит 1—0,5 = 0,5, а частота гомозиготных доминантных генотипов АА составит в популяции q 2 = 0,5 2 = 0,25.
Итак, при анализе природных совокупностей особей (популяций) необходимо различать понятия: частота гена (количественное соотношение аллелей одного какого-либо локуса), частота генотипа (количественное соотношение разных генотипов) и частота фенотипа (количественное соотношение разных фенотипов).
Существование двух (или более) генетически различных форм в популяции в состоянии длительного равновесия в таких соотношениях, что частоту даже наиболее редкой формы нельзя объяснить только повторными мутациями, называется полиморфизмом. В качестве примера полиморфизма можно привести три формы цветков у примулы (Primula vulgaris), рис. 9.8.
Рис. 9.8. Три формы цветков у примулы (Primula vulgaris),
из Ф. Шеппарда, 1970
В природных популяциях примулы (Primula vulgaris) всегда есть особи с длинным пестиком и короткими пыльниками (А), длинными пыльниками и коротким пестиком (Б) и одинаковыми по длине пыльниками и пестиком (В). Самоопыление возможно только у цветов типа В. Гетеростилия способствует перекрестному оплодотворению.
Полиморфизм по механизму возникновения и поддержания разделяется на две большие группы: гетерозиготный и адаптационный.
Гетерозиготный полиморфизм устанавливается в результате давления на популяцию естественного отбора, положительно отбирающего гетерозигот.
Адаптационный полиморфизм — это две или несколько генетически различных форм внутри популяции, подвергающихся положительному отбору в разных экологических условиях.
В качестве примера можно привести адаптационный полиморфизм в популяции двухточечной тлевой (божьей) коровки Adalia bipunctata (рис. 9.9).
Рис. 9.9. Адаптивный полиморфизм в популяции тлевых
(божьих) коровок Adalia bipunctata:
I — процентное содержание черной и красной форм при весеннем (В) и осеннем (О) сборах; II — частота доминантного гена А (черная окраска) в популяциях весной и осенью каждого года (в%) (по Н. В. Тимофееву-Ресовскому и Ю. М. Свирежеву, 1965)
На протяжении 10 лет в изучении популяции тлевой коровки осенью — при уходе на зимовку — черных форм было от 50 до 70%, а весной — при выходе из зимовки — от 30 до 45%. Красных форм осенью было меньше 50%, а весной — больше. Красные формы, как было установлено, лучше переносят в зимний период холод, а черные — интенсивнее размножаются летом. Отсюда отбор направлен на сохранение большего числа красных жуков зимой и черных — летом. Разнонаправленное давление отбора в отдельные периоды жизни популяции способствует выработке устойчивого адаптационного полиморфизма.
Положение о генетическом единстве популяции является одним из наиболее важных выводов популяционной генетики: любая популяция представляет сложную генетическую систему, находящуюся в динамическом равновесии.
Вопрос 1. Сформулируйте закон Харди—Вайнберга.
Закон Харди—Вайнберга. Частота гомозиготных и гетерозиготных организмов в условиях свободного скрещивания при отсутствии давления отбора и других факторов (мутации, миграции, дрейфа генов и др.) остается постоянной, т. е. пребывает в состоянии равновесия.
Закон Харди-Вайнберга устанавливает математическую зависимость между частотами аллелей аутосомных генов и генотипов и выражается следующими формулами:
рА + qа = 1;
р 2 (АА) + 2рq(Аа) + q 2 (аа) = 1,
где р(А) – частота доминантного аллеля гена,
q(а) - частота рецессивного аллеля гена,
р 2 (АА) - частота особей, гомозиготных по доминантному аллелю,
2рq(Аа) – частота гетерозиготных особей,
q 2 (аа) - частота особей, гомозиготных по рецессивному аллелю, то есть частота особей с рецессивным признаком,
р 2 (АА)+ 2рq(Аа) - частота особей с доминантным признаком,
2рq(Аа) + q 2 (аа) – частота особей, в генотипе которых имеется рецессивный аллель.
Необходимо отметить, что закон Харди—Вайнберга, как и другие генетические закономерности, основывается на принципе случайного комбинирования Менделя и точно выполняется при бесконечно большой численности популяции. На практике это означает, что популяции с численностью ниже некоторой минимальной величины не удовлетворяют требованиям закона Харди—Вайнберга.
Теоретически закон Харди-Вайнберга справедлив только для идеальных, или равновесных, популяций. Равновесными популяциями называются такие популяции, в которых выполняются следующие условия:
• популяция бесконечно велика; к ней можно применять законы вероятности, то есть когда в высшей степени маловероятно, что одно случайное событие может изменить частоты аллелей;
• имеет место панмиксия, то есть случайное образование родительских пар, без тенденции вступления особей в брак с партнерами, подобными или противоположными по генотипу;
• все аллели равно влияют на жизнеспособность гамет и потомки от всех возможных скрещиваний имеют равную выживаемость;
• популяция полностью изолирована, то есть, нет миграции особей, дающей приток или отток аллелей;
• новые мутации в данной популяции не появляются;
• отсутствует отбор;
• поколения не перекрываются во времени и не образуются родительские пары из особей, относящихся к разным поколениям.
Несмотря на то, что ни в одной реальной популяции эти условия не соблюдаются, равновесие частот генотипов в них все равно выполняется. Очень часто рассчитанные по формулам закона Харди-Вайнберга величины настолько близки к реальным, что этот закон оказывается вполне пригодным для анализа генетической структуры реальных популяций. Из этого следует, что можно рассчитать ожидаемые частоты генотипов, зная только некоторые из них.
Более того, на основе формул закона Харди-Вайнберга были разработаны подходы для характеристики факторов, изменяющих частоты аллелей в популяциях. Такими факторами являются:
• генетический дрейф;
• мутационный процесс;
• миграция;
• отбор;
• неслучайные скрещивания.
Вопрос 2. Какие процессы приводят к изменению частоты встречаемости генов в популяциях?
Изменение частоты генов в популяциях может происходить как под давлением факторов внешней среды в ходе естественного отбора, так и в виде ненаправленного случайного изменения частоты генов. Например, при миграции животных на новом месте обитания поселяется незначительная часть исходной популяции. Генофонд вновь образованной популяции будет меньше генофонда исходной популяции. Гены, ранее редко встречавшиеся вследствие полового размножения, быстро распространятся среди членов новой популяции. Другим примером могут послужить явления природных катастроф. Так, при лесных пожарах происходит массовая гибель животных и растений. Особи, избежавшие гибели, остаются в живых благодаря случайности. Они и дают начало новой популяции. Генетический состав группы выживших особей определяет генетическую структуру будущей популяции. Изменить частоту генов в популяции могут также периодические колебания численности, связанные со взаимоотношениями хищник—жертва, изоляция популяций друг от друга, близкородственные скрещивания (инбридинг).
Вопрос 3. Почему разные популяции одного вида отличаются по частоте генов?
Популяции, изолированные друг от друга, постоянно находятся под давлением различных комплексов факторов внешней среды. В неодинаковых условиях среды естественный отбор действует в сторону сохранения особей, чей генофонд обеспечивает их набором признаков и свойств, оптимальных для данной ситуации. Поэтому в популяциях, обитающих в различных условиях окружающей среды, частота встречаемости того или иного гена различна.
Вопрос 4. Что такое микроэволюция?
Микроэволюция — это процесс перестройки внутри вида, приводящий к образованию новых популяций, подвидов и заканчивающийся образованием новых видов.
Таким образом, микроэволюция — это самый начальный этап эволюционного процесса, она может происходить в относительно короткие промежутки времени, и ее можно наблюдать и изучать непосредственно. В результате наследственной (мутационной) изменчивости происходят случайные изменения генотипа. Самопроизвольная частота мутаций довольно высока, и 1-2 % половых клеток имеют мутированные гены или измененные хромосомы. Мутации чаще всего рецессивны и, кроме того, редко бывают полезными для вида. Однако если в результате мутации возникают полезные для какой-либо особи изменения, то она получает некоторые преимущества перед другими особями популяции: получает больше пищи или делается устойчивее к влияниям болезнетворных бактерий и вирусов и т.п. Например, возникновение длинной шеи позволило предкам жирафа питаться листьями с высоких деревьев, что обеспечивало им больше корма, чем особям популяции с короткой шеей.

Из видеоурока вы узнаете о том, что такое популяция. О том, что популяциям свойственна изменчивость, которая приводит не только к разнообразию в популяции, но и к образованию новых подвидов и видов. Также вы узнаете о том, что в популяциях имеются большие запасы аллелей, которые не приносят им какой-либо пользы в данном месте или в данное время. Они сохраняются в популяции в гетерозиготном состоянии, пока в результате изменения условий среды вдруг не окажутся полезными. В данном уроке приводятся следующие понятия: популяция, естественный отбор, популяционная генетика, генофонд, мутационный процесс, гетерозиготность, изменчивость популяции.
В данный момент вы не можете посмотреть или раздать видеоурок ученикам
Чтобы получить доступ к этому и другим видеоурокам комплекта, вам нужно добавить его в личный кабинет, приобретя в каталоге.
Получите невероятные возможности



Конспект урока "Популяция. Генетический состав популяций"
Вид — это исторически сложившаяся совокупность популяций, особи которых обладают наследственным сходством морфологических, физиологических и биохимических особенностей, могут свободно скрещиваться и давать плодовитое потомство, приспособлены к определённым условиям жизни и занимают определённую область — ареал.
В природе совокупность особей вида, группируются в популяции. Популяции населяют какие-то отдельные небольшие участки ареала.
Популяция — это группа одновидовых организмов, занимающих определённый участок территории внутри ареала вида, свободно скрещивающихся между собой и частично или полностью изолированных от других популяций.

Вильгельм Людвиг Иогансен
Численность популяций, да и количество особей в популяциях из-за изменения каких-либо факторов, например климатических, может меняться. Но сами популяции как таковые все равно сохраняются.
Конечно, природа не стабильна и внутри популяций могут возникать мелкие группировки, в которые объединяются особи со сходным поведением или на основе родственных связей. Такие группировки известны нам как стайки рыб или воробьёв прайды львов. Напомним, что прайды ― это семейные группы львов. В которые входят один или нескольких взрослых самцов (обычно не более 3-х), нескольких половозрелых самок и их детёнышей (львят обоих полов). Численность прайда может достигать до 30−40 животных. В прайдах всегда доминируют львы — альфа-самцы.
Но, как ни странно, охотой занимаются только самки. Молодые самцы по достижении половой зрелости изгоняются из прайда. Живя отдельно, они крепнут, взрослеют, а затем создают свои прайды.

В отличие от популяции, группы не способны устойчиво поддерживать сами себя. Они могут смешиваться с другими или распадаться под влиянием каких-либо факторов.
Популяция же занимает ту часть ареала, в которой отсутствуют изоляционные барьеры.
Каждая популяция приспосабливалась (адаптировалась) к среде своего обитания. Характерные черты этого приспособления передаются из поколения в поколение.

Взаимоотношения организмов в популяциях
Составляющие популяцию организмы связаны друг с другом различными взаимоотношениями.
Часто особи одной популяции конкурируют друг с другом за те или иные виды ресурсов. Например, самка гиены рожает от 1 до 3 детенышей. Если оба детеныша самки, то они начинают сражение за доминирование, используя зубы. Эта конкуренция ведет к гибели почти 25% всех гиен на их первом месяце жизни. В результате оставшийся в живых детёныш получает больше различных ресурсов и созревает быстрее.
Особи одной популяции могут поедать друг друга. Например, речной окунь в отсутствии корма питается собственной молодью.
Также особи одного вида могут группироваться и вместе обороняться от хищника.
Таким образом, можно сказать что внутренние взаимоотношения в популяциях парой очень сложны и противоречивы.
В природе действует естественный отбор. Ослабленные организмы погибают. Это улучшает качественный состав популяции (в том числе качество наследственного материала, которым располагает популяция). Естественный отбор повышает способность популяции к выживанию в меняющихся условиях среды.
Сильные организмы занимают место погибших, размножаются и дают начало новому поколению.

В пределах каждой популяции организмов, размножающихся половым путём, идёт постоянный обмен генетическим материалом. А вот скрещивание особей из разных популяций происходит гораздо реже, поэтому генетический обмен между разными популяциями ограничен.
Существование в форме популяции повышает внутреннее разнообразие вида, его устойчивость к местным изменениям условий жизни, позволяет ему закрепляться в новых для себя условиях.
Генетический состав популяции
Научные генетические открытия и теории Дарвина привели к рождению особого направления исследований — популяционной генетики.
Популяционная генетика позволяет объяснить процессы изменения генетического состава популяций, возникновения новых свойств организмов и их закрепление под воздействием естественного отбора.
Каждая популяция характеризуется своим специфическим набором генов (генофондом) с присущим только данной популяции соотношением частот встречаемости разных аллелей.
Генофонд — это совокупное количество генетического материала, который слагается из генотипов отдельных особей.
Как уже отмечалось, природные популяции в разных частях ареала вида обычно более или менее различны.
Почему популяции одного вида могут отличаться? Дело в том, что чаще всего скрещивание между особями одного вида происходит в пределах одной популяции, которая находится на определённой территории.
В каждой достаточно долго существующей совокупности особей могут спонтанно возникать различные мутации, которые в дальнейшем комбинируются (перемешиваются) с разными, уже имеющимися наследственными свойствами.
Под действием мутаций, которые закрепляются в генофондах популяций, изменяются виды, что ведёт к эволюции организмов.
Поэтому можно сказать, что мутационный процесс — это постоянный источник наследственной изменчивости.
В популяции, состоящей из нескольких миллионов особей, в каждом поколении может возникать по несколько мутаций практически каждого гена, который имеется в этой популяции.
Наследственная изменчивость обусловлена возникновением разных типов мутаций и их комбинаций, которые передаются по наследству и впоследствии проявляются у потомства.
Природные популяции насыщены самыми разнообразными мутациями. На это обратил внимание ещё русский учёный Сергей Сергеевич Четвериков, который установил, что значительная часть изменчивости генофонда скрыта от глаз, так как подавляющее большинство возникающих мутаций рецессивны и не проявляются внешне.
Вспомним, что гетерозиготность ─ это присущее всякому гибридному организму состояние, при котором его гомологичные хромосомы несут разные формы (аллели) того или иного гена или различаются по взаиморасположению генов.

Ген, который способен подавлять действие другого гена в онтогенезе, называется доминантным.
Ген, который оказывается подавленным и не может проявить себя в фенотипе, называется рецессивным.
Обычно доминантный ген обозначается прописной буквой ─ А-большое, рецессивный ген (а-малое). Рецессивный ген проявляется в фенотипе только в тех случаях, когда он находится в паре с аналогичным геном (а-малое, а-малое), т.е. когда оба аллельных гена являются рецессивными.
Если один аллельный ген является доминантным, а второй рецессивным (А-большое, а-малое, то в фенотипе проявляются только признаки, свойственные доминантному гену (А-большое).

Рецессивные мутации можно выявить в экспериментах со скрещиванием близкородственных особей.
При таком скрещивании некоторые рецессивные аллели, находившиеся в гетерозиготном и потому скрытом состоянии, перейдут в гомозиготное состояние и смогут проявиться.
Таким образом, благодаря мутациям популяции могут изменяться.
Изменчивость популяции — это важный фактор эволюции. Она повышает внутреннее разнообразие вида. Позволяет закрепиться виду в новых местах обитания.
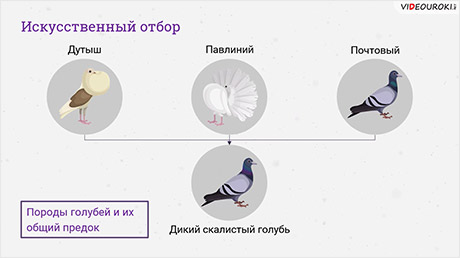
Генетическую изменчивость можно увидеть в ходе искусственного отбора. Когда из популяции выбирают тех особей, у которых какие-либо ценные в хозяйственном отношении признаки выражены наиболее сильно, и скрещивают этих особей между собой.
В фенотипе первого поколения в результате такого скрещивания проявляются необходимые наилучшие выбранные признаки.
Например, у моркови и свёклы ценятся только сладкие корнеплоды, у картофеля — крупные клубни, у мака ― нераскрывающиеся коробочки.
Следовательно, в популяциях возможна генетическая изменчивость буквально по каждому признаку данного организма.
Ещё Сергей Сергеевич Четвериков показал, что большинство вновь возникших мутаций оказываются вредными и, как правило, снижают жизнеспособность особей.
Они сохраняются в популяциях лишь благодаря отбору в пользу гетерозигот.
Однако мутации, вредные в одних условиях, могут повысить жизнеспособность в других условиях.

Так, мутация, вызывающая недоразвитие или полное отсутствие крыльев у насекомых, безусловно, вредна в обычных условиях, и бескрылые особи быстро вытесняются нормальными. Но на островах, где дуют сильные ветры, такие насекомые имеют преимущества перед особями с нормально развитыми крыльями.
Поскольку всякая популяция обычно хорошо приспособлена к своей среде обитания, крупные изменения обычно снижают эту приспособленность.
В популяциях имеются большие запасы таких аллелей, которые не приносят ей какой-либо пользы в данном месте или в данное время; они сохраняются в популяции в гетерозиготном состоянии, пока в результате изменения условий среды вдруг не окажутся полезными.
Как только условия среды изменяются, частота новых аллелей под действием отбора начинает возрастать, и в конечном счёте они становятся основным генетическим материалом.
Таким образом, благодаря приспособлениям к новым условиям среды, популяции адаптируются.
Например, к изменению климата, появлению нового хищника или конкурента и даже к загрязнению среды человеком.
Примером подобной адаптации служит эволюция видов насекомых, устойчивых к инсектицидам. При введении в практику нового инсектицида (яда, действующего на насекомых) для успешной борьбы с насекомым-вредителем бывает достаточно небольшого его количества.
С течением времени концентрацию инсектицида приходится повышать, пока, наконец, он не оказывается недейственным.
Гены, способные обеспечить устойчивость к инсектицидам, очевидно, имелись в каждой из популяций этих видов; их действие и обеспечило в конечном итоге снижение эффективности ядов, использованных для борьбы с вредителями.
Таким образом, мутационный процесс создаёт материал для эволюционных преобразований, формируя резерв наследственной изменчивости в генофонде каждой популяции и виде в целом. Поддерживая высокую степень генетического разнообразия популяций, он создаёт основу для действия естественного отбора и микроэволюции.
Читайте также: