
Генетическая характеристика популяции кратко
Обновлено: 05.07.2024
Ø Наследственным разнообразием генофонда — генетической гетерогенностью генофонда, обусловленной мутационным процессом, потоком генов (миграцией), рекомбинацией.
Вначале генетики предполагали, что особи в популяции имеют гены в основном в гомозиготном состоянии. Позже, после широкого применения в исследованиях гель-электрофореза (метода, выявляющего отличия белков и ферментов по различию в подвижности в электрическом поле), было обнаружено, что гетерозиготность особей популяции — это обычное состояние генофонда у организмов с перекрестным оплодотворением. Генетическая изменчивость природных популяций, судя поданным, полученным с помощью электрофореза в геле, удивительно велика.
Так, беспозвоночные более изменчивы, чем позвоночные (13,4 % и 6,6 %), а растения изменчивы еще более (17 %). Степень гетерозиготности человека 6,7 %. Что это значит? Если допустить, что у человека в генотипе имеется 100 тыс. генных локусов, то каждый индивидуум будет гетерозиготен по 6700 локусам. Вот почему человеку свойственен высокий уровень наследственного разнообразия. Это проявляется в многообразии фенотипов: люди отличаются друг от друга цветом кожи, глаз, волос, формой губ, носа, ушной раковины, рисунком эпидермальных гребней на подушечках пальцев, голосом, отпечатком губ, запахом, группой крови и многими другими признаками.
Установлено, что чем шире диапазон генетической изменчивости популяции, тем быстрее она эволюирует.
Резерв наследственной изменчивости. Это "мобилизационный" резерв рецессивных мутаций, который формируется многими поколениями. С. С. Четвериков с сотрудниками в 1926 г. начал исследовать природные популяции дрозофилы. Инбридировалось потомство диких самок, пойманных в разных частях СССР. Обнаружилось, что многие гетерозиготные особи, будучи фенотипически нормальными, несли в скрытом виде рецессивные мутации.
Эти мутации оказались вне воздействия естественного отбора: они сохранялись и накапливались в гетерозиготах под покровом доминантных аллелей. Этот резерв, благодаря комбинативной изменчивости, используется для создания в каждом поколении новых комбинаций аллелей, а значит и новых вариантов признаков и фенотипов.
В популяции имеются большие запасы таких аллелей, которые не обладают максимальной приспособленностью в данном месте и в данное время. Они сохраняются в генофонде, встречаясь с низкой частотой в гетерозиготном состоянии до тех пор, пока вследствие изменения условий в экосистеме аллели не окажутся способствующими большей выживаемости и размножаемости тем особям, у которых эти аллели имеются. Отбор после этого быстро увеличивает их частоту. Селективная ценность аллеля — непостоянная величина, независимо от того, какой признак он контролирует. Среда изменяется во времени и в пространстве. Из-за этого одни условия в одном местообитании будут благоприятны для особей с данным вариантом гена, а другие — в другом местообитании — для другого варианта гена.
Большая популяция имеет больший генофонд и больший резерв наследственной изменчивости, поэтому она более защищена от изменений среды: при резком изменении среды большая популяция мобилизует генетические резервы и может пережить стрессовый период, а малая популяция имеет малый мобилизационный резерв и поэтому не сможет "дать ответ" экстремальным условиям в форме выживших и размножающихся особей. Один из механизмов сохранения гетерозигот— их превосходство (сверхдоминирование).
Генетический груз. В генофонде популяции имеются вредные мутации, которые при переходе в гомозиготное состояние снижают жизнеспособность особей или вызывают их гибель. Генетический груз — это совокупность летальных и полулетальных мутаций, мутаций стерильности и мутаций, понижающих жизнеспособность особей. Также и аллели, входящие в генетический груз, могут иметь селективную ценность. Так, рецессивный аллель, вызывающий в гомозиготном состоянии серповидную форму эритроцитов и гибель людей, в популяциях, страдающих от малярии, имеет частоту 30 %. Это связано с тем, что гетерозиготы лучше выживают в малярийных районах, чем гомозиготы по доминантному аллелю этого гена.
Биохимический полиморфизм. Этот термин вначале применялся в основном по отношению к морфологическим признакам. Биохимический полиморфизм - это белковый полиморфизм, результат множественного аллелизма: наличие в генофонде популяции нескольких вариантов генов ведет к полиморфизму одного и того же вещества. Гистонесовместимость вызывается именно полиморфизмом белков — у каждого человека свои варианты белков. Антитела также формируются определенными аллелями гена, и они у каждого человека "свои". Группа крови системы АВО — тоже пример полиморфизма белков. У человека обнаружено 14 различных систем групп крови, АВО — одна из них. У людей известно 130 структурных вариантов гемоглобина, 70 вариантов глюкозо-6-фос-фатдегидрогеназы, осуществляющей анаэробное расщепление глюкозы до лактата в эритроцитах, 20 вариантов трансферина — бетаглобулина, переносящего железо из кишечника в костный мозг, более десятка вариантов сывороточного альбумина, 5 разных изоферментов лактатдегидрогеназы. У крупного рогатого скота обнаружено 4 типа гемоглобина, 10 видов трансферинов, несколько типов казеина, несколько типов белков молока.
69. Популяция - элементарная единица микроэволюции. Генетические процессы в популяциях. Закон Харди-Вайнберга.
1)популяция - элементарная единица эволюции. Генетические процессы в популяциях. Закон Харди-Вайнберга
Как уже говорилось, наследуются не сами признаки, а код наследственной информации, определяющий комплекс возможностей развития — норму реакции генотипа, в пределах которой возможно не приводящее к гибели взаимодействие развивающейся особи со средой. При этом наследственная реализация каждого признака или свойства определяется не одним, а, как правило, очень многими генами (принцип полимерии в действии генов); с другой стороны, любой ген оказывает влияние не на один, а на многие признаки (принцип плейотропии в действии гена). Границы характерной для данного генотипа нормы реакции могут быть выражены, таким образом, лишь совокупностью фенотипов, развившихся из этого генотипа при всех возможных условиях среды.
P1 a1 + p2 a2 + p3 a3 + . + pi ai = 1
Q1 b1 + q2 B2 + q3 b3 + . + qi bi = 1
R1 c1 + r2 c2 + r3 c3 + . + ri ci = 1
В этих уравнениях символами Pi, Qi, Ri обозначены относительные частоты аллелей разных генов. Генофонд. Генофондом называют Совокупность всех генотипов в популяции. При изучении природных популяций часто приходится сталкиваться с полным доминированием: фенотипы гомозигот АА и гетерозигот Аа неразличимы. Кроме того, в природе широко распространено полигенное определение признаков, причем типы взаимодействия неаллельных генов (комплементарность, эпистаз, полимерия) не всегда известны. Поэтому на практике часто изучают не генофонд, а Фенофонд популяций, то есть соотношение фенотипов.
Структура популяции по качественным признакам. Харди и Вайнберг показали, что генетические расщепления, которые происходят в каждом поколении у диплоидных организмов, сами по себе не изменяют общего состава генофонда. В идеальной популяции соблюдаются пять условий: 1) новые мутации в данной популяции не появляются; 2) популяция полностью изолирована, т. е. нет миграции особей — носителей генов в популяцию (иммиграция) и из популяции (эмиграция); 3) популяция бесконечно велика, к ней можно применять законы вероятности, т. е. когда в высшей степени маловероятно, что одно случайное событие может изменить частоты аллелей; 4) скрещивания случайны, т. е. происходит чисто случайное образование родительских пар — панмиксия; 5) все аллели равно влияют на жизнеспособность гамет. Харди и Вайнберг с помощью математического уравнения показали, что пропорции аллелей А и а в Такой идеальной популяции не изменяются от поколения к поколению. Остаются постоянными частоты трех возможных комбинаций этих аллелей — генотипы АА, Аа И Аа. Частоты генов находятся в состоянии равновесия по этим аллелям. Это равновесие выражается уравнением: Р2 + 2Pq + Q2 = 1, где Р — частота одного аллеля, Q — другого. Р + Q Всегда составляет единицу, Р2 И Q2 — частоты особей, гомозиготных по соответствующему аллелю, 2Pq — частота гетерозигот.
70. Способы видообразования.
Способы видообразования
Видообразование может быть рассмотрено в территориальном и филогенетическом аспектах. Новый вид может возникнуть из одной или группы смежных популяций, расположенных на периферии ареала исходного вида. Такое видообразование называется аллопатрическим (от греч. allos — иной и patris — родина). В других случаях новый вид может возникнуть внутри ареала исходного вида, как бы внутри вида; этот путь видообразования называется симпатрическим (от греч. syn — вместе и patris — родина). Новый вид может возникнуть посредством постепенного изменения одного и того же вида во времени, без какой-либо дивергенции исходных групп. Такое видообразование называется филетическим. Новый вид может возникнуть путем разделения единого предкового вида (дивергентное видообразование). Наконец, новый вид может возникнуть в результате гибридизации двух уже существующих видов — гибридогенное видообразование. Кратко опишем эти основные пути видообразования.
Называемое иногда географическим) иллюстрируется приведенными выше примерами возникновения вида у больших чаек и в группе австралийских мухоловок. При аллопатрическом видообразовании новые виды могут возникнуть путем фрагментации, распадения ареала широко распространенного родительского вида. Примером такого процесса может быть возникновениевидов ландыша. Другой способ аллопатрического видообразования – видообразование при расселении исходного вида, в процессе которого все более удаленные от центра расселения периферийные популяции и их группы, интенсивно преобразуясь в новых условиях, становятся родоначальниками видов. Примеры, аналогичные видообразованию в группе больших чаек, известны и для других птиц, некоторых рептилий, амфибий, насекомых.
В основе аллопатрического видообразования лежат те или иные формы пространственной изоляции, и этот путь видообразования всегда сравнительно медленный, происходящий на протяжении сотен тысяч поколений. Именно за такие длительные промежутки времени в изолированных частях населения вида вырабатываются те биологические особенности, которые приводят к репродуктивной самостоятельности даже при нарушении первичной изолирующей преграды. Аллопатрическое видообразование всегда связано с историей формирования видового ареала.
При видообразовании симпатрическим путем новый вид возникает внутри ареала исходного вида.
Полиплоидные формы, как правило, крупнее и способны существовать в более суровых физико-географических условиях. Именно поэтому в высокогорьях и в Арктике число полиплоидных видов растений резко увеличено. Среди животных полиплоидия при видообразовании играет несравненно меньшую роль, чем у растений, и во всех случаях связана с партеногенетическим способом размножения (например, у иглокожих, членистоногих, аннелид и других беспозвоночных).
Второй способ симпатрического видообразования — путем гибридизации с последующим удвоением числа хромосом — аллополиплоидия.
Наконец, последним, достаточно изученным способом симпатрического видообразования является возникновение новых форм в результате сезонной изоляции. Известно существование ярко выраженных сезонных рас у растений, например у погремка Alectorolophus major, раннецветущие и позднецветущие формы которого полностью репродуктивно изолированы друг от друга, и, если условие отбора сохраняется, лишь вопрос времени — когда эти формы приобретут ранг новых видов. Аналогично положение с яровыми и озимыми расами проходных рыб; возможно, что эти формы уже являются разными видами, очень схожими морфологически, но изолированными генетически (виды-двойники).
Особенность симпатрического пути видообразования — возникновение новых видов, морфофизиологически близких к исходному виду. Так, при полиплоидии увеличиваются размеры, но общий облик растений сохраняется, как правило, неизменным; при хромосомных перестройках наблюдается та же картина; при экологической (сезонной) изоляции возникающие формы также обычно оказываются морфологически слабо различимыми. Лишь в случае гибридогенного возникновения видов появляется новая видовая форма, отличная от каждого из родительских видов (но также имеющая признаки, характерные для двух исходных видов).
Этот тип видообразования обычен у растений: по некоторым подсчетам более 50 % видов растений представляют собой гибридогенные формы — аллополиплоиды. Укажем лишь на некоторые. Культурная слива (Prunus domestica) с 2n=48 возникла путем гибридизации терна (Р. spinosa, 2n=32) с алычой (Р. divaricata, 2n=16) с последующим удвоением числа хромосом. Некоторые виды пикульников, малины, Табаков, брюквы, полыни, ирисов и других растений — такие же аллополиплоиды гибридогенного происхождения.
Интересен случай возникновения нового симпатрического вида у Spartir townsendii (2n=120) на основе гибридизации с последующим удвоением числа хромосом местного английского вида S. stricta (2n=50) и завезенной в 70-х годахXIX в. из Северной Америки S. alternifolia (2n=70). Сейчас ареал этого вида интенсивно расширяется за счет сокращения ареала местного европейского вида. Другой пример гибридогенного вида, возникшего симпатрически, — рябинокизильник (Sorbocotaneaster), сочетающий признаки рябины и кизильника и распространенный в середине 50-х годов в лесах южной Якутии по берегам среднего течения р. Алдан. В результате гибридогенного видообразования особенно часто могут образовываться комплексы видов (или так называемых полувидов), связанных между собой гибридизацией — сингамеоны (В. Грант). В случае таких гибридных комплексов иногда бывает трудно обнаружить четкие границы между отдельными видами, хотя виды как устойчивые генетические системы выделяются вполне определенно.
59.
71. Популяционная структура человечества. Человек как объект действия эволюционных факторов.
ПОПУЛЯЦИОННАЯ СТРУКТУРА ВИДА
В природе практически не существует видов, которые были бы повсеместно распространены. Обычно каждый вид имеет свою область распространения - ареал, границы которого определяются границами пригодных для данного вида условий обитания. Космополитами - видами, обитающими повсеместно - являются прежде всего человек, сумевший освободиться из-под влияния окружающей среды, и (с определенной долей условности) некоторые обитающие совместно с ним животные, такие как серая крыса и рыжий таракан
Границы ареалов видов со временем изменяются. Как правило, это связано с изменением условий существования, а также с адаптацией видов к новой среде обитания. В настоящее время ареалы многих видов меняются под влиянием хозяйственной деятельности человека. При этом ареал может сокращаться, как у соболя, или расширяться, как у зайца-русака.
Популяция - совокупность особей того или иного вида, в течение большого числа поколений населяющих определенное пространство, внутри которого особи могут относительно свободно скрещиваться друг с другом, в то время как обмен особями с соседними популяциями в значительной мере затруднен.
Популяционная структура видов зачастую бывает довольно сложной, и локальные поселения можно объединить в более крупные группировки. Популяции высокого ранга, объединяющие несколько более мелких единиц, стабильны во времени, и могут обладать собственной эволюционной судьбой.
В силу того, что разные популяции в большей или меньшей степени изолированы друг от друга, между ними накапливаются генетические и морфологические различия - формируется внутривидовая географическая изменчивость.
Популяция– это совокупность особей одного вида, длительно населяющих определенное пространство, способных к панмиксии и отделенных от других особей этого вида какой-либо формой изоляции.
Экологическая характеристика популяции – определенный ареал, численность особей, возрастной состав, половой состав, плотность. Места обитания – луг, поле, лес, озеро и т.д.
По численности различают популяции от нескольких сот особей до нескольких тысяч. Чем меньше популяция, тем больше угроза ее вымирания.
Устойчивая популяция включает в себя все возрастные группы - от новорожденных до стариков.
По половому составу популяции в основном характеризуются равным соотношением мужских и женских особей. У человека при рождении 106 мальчиков на 100 девочек; к 18 годам 100: 100; к 50 годам 85: 100; а к 80 годам 50:100. Плотность популяции – количество особей приходящихся на единицу площади.
Генетическая характеристика популяции.
Генетически популяция характеризуется ее генофондом (совокупность аллелей образующих генотипы организмов в этой популяции.)
Генофонды природных популяций отличаются:
1) наследственным разнообразием (генетическим полиморфизмом)
2) генетическим единством
3) динамическим равновесием доли особей с разными генотипами.
Наследственное разнообразие – это присутствие одновременно различных аллелей отдельных генов. Оно создается мутационным процессом.
Генетическое единство – популяции обуславливается достаточным уровнем панмиксии. В условиях случайного подбора скрещивающихся особей источником аллелей является весь генофонд популяции.
Динамическое равновесие – в популяцию одновременно могут входить особи с доминантными и рецессивными признаками. В пределах генофонда популяции доля их, при соблюдении некоторых условий, из поколения в поколение не изменяются. Это было доказано законом Харди – Вайнберга.
Закон Харди – Вайнберга.
"В популяции из бесконечно большого числа свободно скрещивающихся особей в отсутствии мутаций, избирательной миграции организмов с различными генотипами и давления естественного отбора первоначальные частоты аллелей сохраняются из поколения в поколение".(1908 г.)
Допустим, что в генофонде популяции, удовлетворяющей описанным условиям, некий ген представлен аллелями А1 и А2, которые встречаются с частотой p и q. Так как других аллелей в данном генофонде не встречается, то p + q = 1. При этом q = 1- p , p = 1-q. Гаметы с аллелям А1 будут встречаться с частотой p, а с А2 – q. Если скрещивание случайное, то при оплодотворении образуются зиготы со следующими генотипами:
| А1p | A2p | |
| A1 p | A1 A1 p 2 | A1 A2 p q |
| A2 q | A1 A2 p q | A2 A2 q 2 |
т.о. генотипы A1 A1 с частотой p 2
т.о. генотипы A1 A2 с частотой 2 pq
т.о. генотипы A2 A2 с частотой q 2
частота всех генотипов p 2 + p q+ q 2
По достижении половой зрелости особи A1 A1 и A2 A2 дадут в среднем одинаковое число гамет с частотой p 2 и q 2 соответственно, а
то доля А1 гамет равна p 2 + pq= p(p+ q)=p(1)=p
то доля А2 гамет равна q 2 + pq= q (p+ q)= q (1)= q
т.е. частота генов во втором поколении осталась такой же, что была в популяции родителей. Аналогичные расчеты можно произвести для локусов с любым числом аллелей. Уравнение , Харди – Вайнберга рассмотренное выше, справедливо для аутосомных генов.
Популяция– это совокупность особей одного вида, длительно населяющих определенное пространство, способных к панмиксии и отделенных от других особей этого вида какой-либо формой изоляции.
Экологическая характеристика популяции – определенный ареал, численность особей, возрастной состав, половой состав, плотность. Места обитания – луг, поле, лес, озеро и т.д.
По численности различают популяции от нескольких сот особей до нескольких тысяч. Чем меньше популяция, тем больше угроза ее вымирания.
Устойчивая популяция включает в себя все возрастные группы - от новорожденных до стариков.
По половому составу популяции в основном характеризуются равным соотношением мужских и женских особей. У человека при рождении 106 мальчиков на 100 девочек; к 18 годам 100: 100; к 50 годам 85: 100; а к 80 годам 50:100. Плотность популяции – количество особей приходящихся на единицу площади.
Генетическая характеристика популяции.
Генетически популяция характеризуется ее генофондом (совокупность аллелей образующих генотипы организмов в этой популяции.)

Генофонды природных популяций отличаются:
1) наследственным разнообразием (генетическим полиморфизмом)
2) генетическим единством
3) динамическим равновесием доли особей с разными генотипами.
Наследственное разнообразие – это присутствие одновременно различных аллелей отдельных генов. Оно создается мутационным процессом.
Генетическое единство – популяции обуславливается достаточным уровнем панмиксии. В условиях случайного подбора скрещивающихся особей источником аллелей является весь генофонд популяции.
Динамическое равновесие – в популяцию одновременно могут входить особи с доминантными и рецессивными признаками. В пределах генофонда популяции доля их, при соблюдении некоторых условий, из поколения в поколение не изменяются. Это было доказано законом Харди – Вайнберга.
Закон Харди – Вайнберга.
"В популяции из бесконечно большого числа свободно скрещивающихся особей в отсутствии мутаций, избирательной миграции организмов с различными генотипами и давления естественного отбора первоначальные частоты аллелей сохраняются из поколения в поколение".(1908 г.)
Допустим, что в генофонде популяции, удовлетворяющей описанным условиям, некий ген представлен аллелями А1 и А2, которые встречаются с частотой p и q. Так как других аллелей в данном генофонде не встречается, то p + q = 1. При этом q = 1- p , p = 1-q. Гаметы с аллелям А1 будут встречаться с частотой p, а с А2 – q. Если скрещивание случайное, то при оплодотворении образуются зиготы со следующими генотипами:
| А1p | A2p | |
| A1 p | A1 A1 p 2 | A1 A2 p q |
| A2 q | A1 A2 p q | A2 A2 q 2 |
т.о. генотипы A1 A1 с частотой p 2
т.о. генотипы A1 A2 с частотой 2 pq
т.о. генотипы A2 A2 с частотой q 2
частота всех генотипов p 2 + p q+ q 2
По достижении половой зрелости особи A1 A1 и A2 A2 дадут в среднем одинаковое число гамет с частотой p 2 и q 2 соответственно, а
то доля А1 гамет равна p 2 + pq= p(p+ q)=p(1)=p
то доля А2 гамет равна q 2 + pq= q (p+ q)= q (1)= q
т.е. частота генов во втором поколении осталась такой же, что была в популяции родителей. Аналогичные расчеты можно произвести для локусов с любым числом аллелей. Уравнение , Харди – Вайнберга рассмотренное выше, справедливо для аутосомных генов.
Популяци я – совокупность особей 1 вида, живущих в течение многих поколений на одной и той же территории и свободно скркщивающихся между собой и дающих плодовитое потомство.
Популяция – элементарная эволюционная единица.
Критерии популяции:
· Длительно живут на одной территории
Экологическая характеристика популяций:
1. Размеры ареала. Они зависят от радиусов индивидуальной активности организмов данного вида и особенностей природных условий на соответствующей территории.
2. Численность особейв популяциях организмов разных видов различается. Существуют минимальное минимальное значение численности, при которых популяция способна поддерживать себя во времени. Сокращение численности ниже этого минимума приводит к вымиранию популяции.
3. Возрастная структурапопуляций организмов разных видов варьирует в зависимости от продолжительности жизни, интенсивности размножения, возраста достижения половой зрелости.
4. Половой состав популяций обуславливается эволюционно закрепленными механизмами формирования первичного (на момент зачатия), вторичного (на момент рождения) и третичного (во взрослом состоянии) соотношения полов.
Экологическая ниша
Наличие изоляции
Генетическая характеристики популяции:
Генетически популяция характеризуется ее генофондом (аллелофондом). Он представлен совокупностью аллелей, образующих генотипы организмов данной популяции. Генофонды природных популяций отличает наследственное разнообразие, генетическое единство, динамическое равновесие доли особей с разными генотипами.
Наследственное разнообразие заключается в присутствии в генофонде одновременно различных аллелей отдельных видов. Первично оно создается мутационным процессом. Мутации, будучи обычно рецессивными и не влияя на фенотипы гетерозиготных организмов, сохраняются в генофондах популяций в скрытом от естественного отбора состоянии. Накапливаясь, они образуют резерв наследственной изменчивости. Благодаря комбинативной изменчивости этот резерв используется для создания в каждом поколении новых комбинаций аллелей. Объем такого резерва огромен.
Генетического единство популяции обуславливается достаточным уровнем панмиксии. В условиях случайного подбора скрещивающихся особей источником аллелей для генотипов организмов последовательных поколений является весь генофонд популяции. Генетическое единство проявляется так же в общей генотипической изменчивости популяции при изменении условий существования, что обуславливает как выживание вида, так т образование новых видов.
Механизмы определяющие генетическую структуру популяций:
1. Поддерживающие равновесия – действие закона Харди-Вайбренга
2. Нарушающие равновесие – действие элементарных эволюционных факторов.
Естественный отбор — движущая сила эволюции. Формы естественного отбора.
Особенности действия естественного отбора в человеческих популяциях.
Естественный отбор - избирательное воспроизведение одних генотипов и гибель других.
Основная функция естественного отбора заключается в устранении из популяций организмов с неудачными комбинациями генов и сохранение генотипов, которые не нарушают процесса приспособительного формообразования. Действие естественного отбора выражается в пределах популяции, но объектами этого действия являются отдельные особи, т.е. естественный отбор работает на элементарном эволюционном материале.
Результатом действия естественного отбора является или элиминация того или иного аллеля, или возникновение полиморфизма, заключающегося в том, что в популяции присутствуют 2 или более аллеля одного и того же локуса. Отбор может действовать относительно рецессивного или доминантного гена.
Формы естественного отбора:
· Движущий – отбор, способствующий сдвигу среднего значения признака или свойства. Эта форма приводит к возникновению адаптивных признаков. При направленном изменении среды чаще выживают особи, обладающие индивидуальными особенностями, соответствующими этому изменению; особи с уклонениями в противоположную сторону, не адекватними изменению внешних условий, чаще гибнут. Утрата признака – обычно результат действия движущей формы естественного отбора. Утрата признака обычно результат действия движущей формы отбора. (утрата крыльев у части птиц и насекомых, пальцев у копытных, конечностей у змей, глаз у пещерных животных). Таким образом, движущая форма отбора приводит к выработке новых приспособлений через направленную перестройку популяции, а это, в свою очередь, сопровождается перестройкой генотипа особей.
· Стабилизирующий – форма естественного отбора, направленная на поддержание и повышение устойчивости реализации в популяции среднего, ранее сложившегося значения признака или свойства. Примером может служить связь между массой новорожденных детей и их смертностью: чем сильнее отклонение в любую сторону от средней нормы (3,6 кг), тем реже такие дети выживают. Таким образом, важнейшим результатом стабилизирующего отбора является сохранение, стабилизация уже существующих признаков и уже сформировшейся нормы реакции по этим признакам.
Другие формы естественного отбора:
- групповой отбор и др.
Естественный отбор в человеческих популяции действует слабо, в результате условий жизни людей, развитию медицины. В результате происходит накапливание рецессивных мутаций.
Наиболее жестоко отбор идет в эмбриональном периоде:

Из видеоурока вы узнаете о том, что такое популяция. О том, что популяциям свойственна изменчивость, которая приводит не только к разнообразию в популяции, но и к образованию новых подвидов и видов. Также вы узнаете о том, что в популяциях имеются большие запасы аллелей, которые не приносят им какой-либо пользы в данном месте или в данное время. Они сохраняются в популяции в гетерозиготном состоянии, пока в результате изменения условий среды вдруг не окажутся полезными. В данном уроке приводятся следующие понятия: популяция, естественный отбор, популяционная генетика, генофонд, мутационный процесс, гетерозиготность, изменчивость популяции.
В данный момент вы не можете посмотреть или раздать видеоурок ученикам
Чтобы получить доступ к этому и другим видеоурокам комплекта, вам нужно добавить его в личный кабинет, приобретя в каталоге.
Получите невероятные возможности



Конспект урока "Популяция. Генетический состав популяций"
Вид — это исторически сложившаяся совокупность популяций, особи которых обладают наследственным сходством морфологических, физиологических и биохимических особенностей, могут свободно скрещиваться и давать плодовитое потомство, приспособлены к определённым условиям жизни и занимают определённую область — ареал.
В природе совокупность особей вида, группируются в популяции. Популяции населяют какие-то отдельные небольшие участки ареала.
Популяция — это группа одновидовых организмов, занимающих определённый участок территории внутри ареала вида, свободно скрещивающихся между собой и частично или полностью изолированных от других популяций.

Вильгельм Людвиг Иогансен
Численность популяций, да и количество особей в популяциях из-за изменения каких-либо факторов, например климатических, может меняться. Но сами популяции как таковые все равно сохраняются.
Конечно, природа не стабильна и внутри популяций могут возникать мелкие группировки, в которые объединяются особи со сходным поведением или на основе родственных связей. Такие группировки известны нам как стайки рыб или воробьёв прайды львов. Напомним, что прайды ― это семейные группы львов. В которые входят один или нескольких взрослых самцов (обычно не более 3-х), нескольких половозрелых самок и их детёнышей (львят обоих полов). Численность прайда может достигать до 30−40 животных. В прайдах всегда доминируют львы — альфа-самцы.
Но, как ни странно, охотой занимаются только самки. Молодые самцы по достижении половой зрелости изгоняются из прайда. Живя отдельно, они крепнут, взрослеют, а затем создают свои прайды.

В отличие от популяции, группы не способны устойчиво поддерживать сами себя. Они могут смешиваться с другими или распадаться под влиянием каких-либо факторов.
Популяция же занимает ту часть ареала, в которой отсутствуют изоляционные барьеры.
Каждая популяция приспосабливалась (адаптировалась) к среде своего обитания. Характерные черты этого приспособления передаются из поколения в поколение.

Взаимоотношения организмов в популяциях
Составляющие популяцию организмы связаны друг с другом различными взаимоотношениями.
Часто особи одной популяции конкурируют друг с другом за те или иные виды ресурсов. Например, самка гиены рожает от 1 до 3 детенышей. Если оба детеныша самки, то они начинают сражение за доминирование, используя зубы. Эта конкуренция ведет к гибели почти 25% всех гиен на их первом месяце жизни. В результате оставшийся в живых детёныш получает больше различных ресурсов и созревает быстрее.
Особи одной популяции могут поедать друг друга. Например, речной окунь в отсутствии корма питается собственной молодью.
Также особи одного вида могут группироваться и вместе обороняться от хищника.
Таким образом, можно сказать что внутренние взаимоотношения в популяциях парой очень сложны и противоречивы.
В природе действует естественный отбор. Ослабленные организмы погибают. Это улучшает качественный состав популяции (в том числе качество наследственного материала, которым располагает популяция). Естественный отбор повышает способность популяции к выживанию в меняющихся условиях среды.
Сильные организмы занимают место погибших, размножаются и дают начало новому поколению.

В пределах каждой популяции организмов, размножающихся половым путём, идёт постоянный обмен генетическим материалом. А вот скрещивание особей из разных популяций происходит гораздо реже, поэтому генетический обмен между разными популяциями ограничен.
Существование в форме популяции повышает внутреннее разнообразие вида, его устойчивость к местным изменениям условий жизни, позволяет ему закрепляться в новых для себя условиях.
Генетический состав популяции
Научные генетические открытия и теории Дарвина привели к рождению особого направления исследований — популяционной генетики.
Популяционная генетика позволяет объяснить процессы изменения генетического состава популяций, возникновения новых свойств организмов и их закрепление под воздействием естественного отбора.
Каждая популяция характеризуется своим специфическим набором генов (генофондом) с присущим только данной популяции соотношением частот встречаемости разных аллелей.
Генофонд — это совокупное количество генетического материала, который слагается из генотипов отдельных особей.
Как уже отмечалось, природные популяции в разных частях ареала вида обычно более или менее различны.
Почему популяции одного вида могут отличаться? Дело в том, что чаще всего скрещивание между особями одного вида происходит в пределах одной популяции, которая находится на определённой территории.
В каждой достаточно долго существующей совокупности особей могут спонтанно возникать различные мутации, которые в дальнейшем комбинируются (перемешиваются) с разными, уже имеющимися наследственными свойствами.
Под действием мутаций, которые закрепляются в генофондах популяций, изменяются виды, что ведёт к эволюции организмов.
Поэтому можно сказать, что мутационный процесс — это постоянный источник наследственной изменчивости.
В популяции, состоящей из нескольких миллионов особей, в каждом поколении может возникать по несколько мутаций практически каждого гена, который имеется в этой популяции.
Наследственная изменчивость обусловлена возникновением разных типов мутаций и их комбинаций, которые передаются по наследству и впоследствии проявляются у потомства.
Природные популяции насыщены самыми разнообразными мутациями. На это обратил внимание ещё русский учёный Сергей Сергеевич Четвериков, который установил, что значительная часть изменчивости генофонда скрыта от глаз, так как подавляющее большинство возникающих мутаций рецессивны и не проявляются внешне.
Вспомним, что гетерозиготность ─ это присущее всякому гибридному организму состояние, при котором его гомологичные хромосомы несут разные формы (аллели) того или иного гена или различаются по взаиморасположению генов.

Ген, который способен подавлять действие другого гена в онтогенезе, называется доминантным.
Ген, который оказывается подавленным и не может проявить себя в фенотипе, называется рецессивным.
Обычно доминантный ген обозначается прописной буквой ─ А-большое, рецессивный ген (а-малое). Рецессивный ген проявляется в фенотипе только в тех случаях, когда он находится в паре с аналогичным геном (а-малое, а-малое), т.е. когда оба аллельных гена являются рецессивными.
Если один аллельный ген является доминантным, а второй рецессивным (А-большое, а-малое, то в фенотипе проявляются только признаки, свойственные доминантному гену (А-большое).

Рецессивные мутации можно выявить в экспериментах со скрещиванием близкородственных особей.
При таком скрещивании некоторые рецессивные аллели, находившиеся в гетерозиготном и потому скрытом состоянии, перейдут в гомозиготное состояние и смогут проявиться.
Таким образом, благодаря мутациям популяции могут изменяться.
Изменчивость популяции — это важный фактор эволюции. Она повышает внутреннее разнообразие вида. Позволяет закрепиться виду в новых местах обитания.
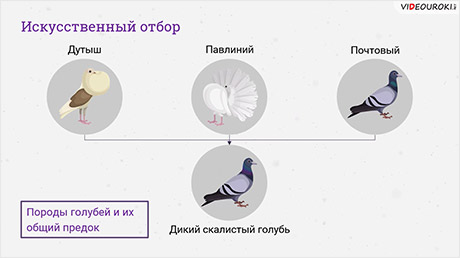
Генетическую изменчивость можно увидеть в ходе искусственного отбора. Когда из популяции выбирают тех особей, у которых какие-либо ценные в хозяйственном отношении признаки выражены наиболее сильно, и скрещивают этих особей между собой.
В фенотипе первого поколения в результате такого скрещивания проявляются необходимые наилучшие выбранные признаки.
Например, у моркови и свёклы ценятся только сладкие корнеплоды, у картофеля — крупные клубни, у мака ― нераскрывающиеся коробочки.
Следовательно, в популяциях возможна генетическая изменчивость буквально по каждому признаку данного организма.
Ещё Сергей Сергеевич Четвериков показал, что большинство вновь возникших мутаций оказываются вредными и, как правило, снижают жизнеспособность особей.
Они сохраняются в популяциях лишь благодаря отбору в пользу гетерозигот.
Однако мутации, вредные в одних условиях, могут повысить жизнеспособность в других условиях.

Так, мутация, вызывающая недоразвитие или полное отсутствие крыльев у насекомых, безусловно, вредна в обычных условиях, и бескрылые особи быстро вытесняются нормальными. Но на островах, где дуют сильные ветры, такие насекомые имеют преимущества перед особями с нормально развитыми крыльями.
Поскольку всякая популяция обычно хорошо приспособлена к своей среде обитания, крупные изменения обычно снижают эту приспособленность.
В популяциях имеются большие запасы таких аллелей, которые не приносят ей какой-либо пользы в данном месте или в данное время; они сохраняются в популяции в гетерозиготном состоянии, пока в результате изменения условий среды вдруг не окажутся полезными.
Как только условия среды изменяются, частота новых аллелей под действием отбора начинает возрастать, и в конечном счёте они становятся основным генетическим материалом.
Таким образом, благодаря приспособлениям к новым условиям среды, популяции адаптируются.
Например, к изменению климата, появлению нового хищника или конкурента и даже к загрязнению среды человеком.
Примером подобной адаптации служит эволюция видов насекомых, устойчивых к инсектицидам. При введении в практику нового инсектицида (яда, действующего на насекомых) для успешной борьбы с насекомым-вредителем бывает достаточно небольшого его количества.
С течением времени концентрацию инсектицида приходится повышать, пока, наконец, он не оказывается недейственным.
Гены, способные обеспечить устойчивость к инсектицидам, очевидно, имелись в каждой из популяций этих видов; их действие и обеспечило в конечном итоге снижение эффективности ядов, использованных для борьбы с вредителями.
Таким образом, мутационный процесс создаёт материал для эволюционных преобразований, формируя резерв наследственной изменчивости в генофонде каждой популяции и виде в целом. Поддерживая высокую степень генетического разнообразия популяций, он создаёт основу для действия естественного отбора и микроэволюции.
- Для учеников 1-11 классов и дошкольников
- Бесплатные сертификаты учителям и участникам
Основные генетические характеристики популяции
Популяция (франц. population – население) – совокупность особей
одного вида, занимающих определенный ареал, свободно скрещивающихся друг с
другом, имеющих общее происхождение, генетическую основу и в той или иной
степени изолированных от других популяций данного вида.
Популяция – элементарная эволюционная структура. Основу современной
эволюционной теории, которую называют неодарвинизмом или синтетической
теорией эволюции, составляет изучение популяционной генетики. Гены,
действуя независимо или совместно с факторами среды, определяют
фенотипические признаки организмов и обуславливают изменчивость в
популяциях. Фенотипы, приспособленные к условиям данной среды или
фенотипы подавляются и, в конце концов, элиминируются. Естественный отбор,
влияя на выживание отдельных особей с данным фенотипом, тем самым
определяет судьбу их генотипа, однако лишь общая генетическая реакция всей
популяции определяет выживание данного вида, а также образование новых
видов. Только те организмы, которые, прежде чем погибнуть успешно произвели
потомство, вносит вклад в будущее своего вида. Для истории данного вида
судьба отдельного организма не имеет существенного значения.
Генетически популяция характеризуется ее генофондом (аллелофондом).
Генофонд представлен совокупностью аллелей, образующих генотипы
организмов данной популяции. В каждой данной популяции состав генофонда из
поколения в поколение может постоянно изменяться. Новые сочетания генов
образуют уникальные генотипы, которые в своем физическом выражении, т. е. в
форме фенотипов, подвергаются давлению факторов среды, производящих
непрерывный отбор и определяющих, какие гены будут переданы следующему
Популяция, генофонд которой непрерывно изменяется из поколения в
поколение, претерпевает эволюционное изменение. Статичный генофонд отражает
отсутствие генетической изменчивости среди особей данного вида и отсутствие
Генофонды природных популяций отличает наследственное разнообразие
(генетическая гетерогенность, или полиморфизм), генетическое единство,
динамическое равновесие доли особей с разными генотипами.
Наследственное разнообразие заключается в присутствии в генофонде
одновременно различных аллелей отдельных генов. Первично оно создается
мутационным процессом. Мутации, будучи обычно рецессивными и не влияя на
фенотипы гетерозиготных организмов, сохраняются в генофондах популяций в
скрытом от естественного отбора состоянии. Накапливаясь, они образуют
резерв наследственной изменчивости. Благодаря комбинативной изменчивости
этот резерв используется для создания в каждом поколении новых комбинаций
аллелей. Объем такого резерва огромен. Так, при скрещивании организмов,
различающихся по 1000 локусов, каждый из которых представлен десятью
аллелями, количество вариантов генотипов достигает 101000.
Генетическое единство популяции обуславливается достаточным уровнем
панмиксии. В условиях случайного подбора скрещивающихся особей источником
аллелей для генотипов организмов последовательных поколений является весь
генофонд популяций. Генетическое единство проявляется также в общей
генетической реализации популяции при изменении условий существования, что
обуславливает как выживание вида, так и образование новых видов.
ЗАКОН ХАРДИ – ВАЙНБЕРГА
Любой физический признак определяется одним или несколькими генами.
Каждый ген может существовать в нескольких различных формах – аллелях.
Число организмов в данной популяции, несущих определенный аллель,
определяет частоту данного аллеля (которую иногда называют частотой гена,
что менее точно). Например, у человека частота доминантного аллеля,
определяющего нормальную пигментацию кожи, волос и глаз, равна 99%.
Рецессивный аллель, детерминирующий отсутствие пигментации – так называемый
альбинизм, - встречается с частотой 1%. В популяционной генетике частоту
аллелей или генов выражают не в процентах, а в десятичных дробях. Таким
образом, в данном случае частота доминантного аллеля равна 0,99 , а частота
рецессивного аллеля альбинизма – 0,01. Общая частота аллелей в популяции
составляет 100%, или 1,0.
Как это принято в классической генетике, аллели обозначаются буквами,
например A – доминантный аллель, a – рецессивный и рецессивный) в генофонде
популяции, используются символы p и q. Таким образом,
где p – частота доминантного, q – частота рецессивного аллеля. (В примере с
пигментацией у человека p = 0,99 , а q =0,01). Значение этого уравнения
состоит в том, что, зная частоту одного из аллелей, можно определить
Частоты отдельных аллелей в генофонде позволяют вычислять
генетические изменения в данной популяции и определять частоту генотипов.
Математическая зависимость между частотами аллелей и генотипов в
популяциях была установлена в 1908 г. независимо друг от друга английским
математиком Дж. Харди и немецким врачом В. Вайнбергом. Эта зависимость
получила название закон Харди – Вайнберга (равновесие Харди – Вайнберга).
Закон этот гласит: «В бесконечно большой популяции из свободно
скрещивающихся особей в отсутствие мутаций, избирательной миграции
организмов с различными генотипами и давления естественного отбора
первоначальные частоты доминантного и рецессивного аллелей сохраняются
Поэтому любые изменения частоты аллелей должны быть обусловлены
нарушением одного или нескольких перечисленных выше условий. Все эти
нарушения способны вызвать эволюционное изменение. Эти изменения и их
скорость можно изучить и измерить с помощью уравнения Харди – Вайнберга.
Если имеется два организма, один гомозиготный по доминантному аллелю
A, а другой – по рецессивному аллелю a, то все их потомки будут
Если наличие доминантного аллеля A обозначить символом p, а
рецессивного аллеля a – символом q, то картину скрещивания между особями
F1, возникающие при этом генотипы и их частоты можно представить следующим
Используя символы p и q результаты приведенного выше скрещивания можно
представить следующим образом:
p2 – доминантные гомозиготы
q2 – рецессивные гомозиготы.
Такое распределение возможных генотипов носит статистический характер и
основано на вероятностях. Три возможных генотипа, образующихся при таком
скрещивании, представлены со следующими частотами:
Сумма частот трех генотипов, представленных в рассматриваемой популяции,
равна единице; пользуясь символами p и q, можно сказать, что вероятности
На математическом языке [pic] представляет собой уравнение вероятности,
тогда как [pic] является квадратом этого уравнения
p – частота доминантного аллеля;
q – частота рецессивного аллеля;
p2 – гомозиготный доминантный тип;
2pq – гетерозиготный генотип;
q2 – гомозиготный рецессивный генотип,
можно вычислить частоты всех аллелей и генотипов, пользуясь выражениями
для частот аллелей: [pic]
для частот генотипов: [pic]
Однако, для большинства популяций частоту обоих аллелей можно
вычислить только по доле особей, гомозиготных по рецессивному аллелю, так
как это единственный генотип, который можно распознать по его
Например, один человек из 10000 – альбинос, т.е. частота
альбинотического генотипа составляет 1 на 10000. Поскольку аллель
альбинизма рецессивен, альбинос должен быть гомозиготным по рецессивному
гену, т.е. на языке теории вероятности
Зная, что q2 =0,0001, можно определить частоты аллеля альбинизма (q),
доминантного аллеля нормальной пигментации (p), гомозиготного доминантного
генотипа (p2) и гетерозиготного генотипа (2pq). Так как q2=0,0001,
т.е. частота аллеля альбинизма в популяции равна 0,01 или 1%. Поскольку
частота доминантного аллеля в популяции равна 0,99 или 99%. А если
т.е. частота гетерозиготного генотипа составляет 0,0198; иными словами,
примерно 2% индивидуумов в данной популяции несут аллель альбинизма либо в
гетерозиготном либо в гомозиготном состоянии.
Гетерозиготных индивидуумов, нормальных по фенотипу, но обладающих
рецессивным геном, который в гомозиготном состоянии может вызвать нарушение
метаболизма, называют носителями. Существование в популяции неблагоприятных
аллелей в составе гетерозиготных генотипов называют генетическим грузом.
Как показывают вычисления с использованием уравнения Харди – Вайнберга,
частота носителей в популяции всегда выше, чем можно было бы ожидать на
основании фенотипического проявления данного дефекта.
СЛЕДСТВИЯ УРАВНЕНИЯ ХАРДИ – ВАЙНБЕРГА
Из уравнения Харди – Вайнберга следует, что значительная доля
имеющихся в популяции рецессивных аллелей находится у гетерозиготных
носителей. Фактически гетерозиготные генотипы служат важным потенциальным
источником генетической изменчивости. Это приводит к тому, что в каждом
поколении из популяции может элиминироваться лишь очень малая доля
рецессивных аллелей. Только те рецессивные аллели, которые находятся в
гомозиготном состоянии, проявятся в фенотипе и тем самым подвергнутся
селективному воздействию факторов среды и могут быть элиминированы. Многие
рецессивные аллели элиминируются потому, что они неблагоприятны для
фенотипа – обуславливают либо гибель организма еще до того как он успеет
Однако не все рецессивные аллели неблагоприятны для популяции.
Например, у человека из всех групп крови чаще всего встречается группа 0,
соответствующая гомозиготности по рецессивному аллелю.
Другим примером служит серновидноклеточная анемия. наследственное
заболевание, широко распространенное в ряде областей Африки, индии, у
негритянского населения Северной Америки. Индивидуумы, гомозиготные по
соответствующему рецессивному аллелю, обычно умирают не достигнув половой
зрелости и элиминируя таким образом из популяции по два рецессивных аллеля.
Что касается гетерозигот, то они не гибнут. Установлено, что во многих
частях земного шара частота аллеля серновидноклеточности остается
относительно стабильной. У некоторых африканских племен частота
гетерозиготного фенотипа достигает 40%. Раньше думали, что этот уровень
поддерживается за счет появления новых мутантов. Однако в результате
дальнейших исследований выяснилось, что во многих частях Африки, где среди
факторов, угрожающих здоровью и жизни, важное место занимает малярия, люди,
несущие аллель серновидноклеточности, обладают повышенной резистентностью к
этой болезни. В малярийных районах Центральной Африки это селективное
преимущество гетерозиготного генотипа поддерживает частоту аллеля
серновидноклеточности на уровне 10 – 20%. У североамериканских негров, уже
200 – 300 лет не испытывающих на себе селективного эффекта малярии, частота
аллеля серновидноклеточности упала до 5%.
Этот пример эволюции в действии ясно демонстрирует селективное
влияние среды на частоту аллелей – механизм нарушающий равновесие Харди –
Вайнберга. Именно такого рода механизмы вызывают в популяциях сдвиги,
ведущие к эволюционному изменению.
ФАКТОРЫ, ВЫЗЫВАЮЩИЕ ИЗМЕНЕНИЯ
Принцип Харди – Вайнберга носит чисто теоретический характер. Очень
немногие популяции находятся в условиях, при которых сохраняется
Половая рекомбинация (кроссинговер во время мейоза, независимое
распределение хромосом при мейозе, случайное оплодотворение) обуславливает
перетасовку генов, лежащую в основе происходящих непрерывных изменений. Но
хотя эти процессы и приводят к образованию новых генотипов и изменяют их
частоты, они не вызывают никакого изменения имеющихся аллелей, так что
частоты аллелей остаются постоянными.
А эволюционные изменения происходят вслед за появлением новых
аллелей, а главным источником последних служат мутации.
Большой вклад в популяционную генетику внес русский ученый С. С.
Четвериков, обративший внимание на насыщенность природных популяций
рецессивными мутациями, а также на колебания частоты генов в популяциях в
зависимости от действия факторов внешней среды.
Мутационный процесс, изменяя частоту одного аллеля по отношению к
другому, оказывает на генофонд популяции прямое действие. За счет мутантных
аллелей происходит формирование резерва наследственной изменчивости.
Благодаря мутационному процессу поддерживается высокий уровень
наследственного разнообразия природных популяций. Совокупность аллелей,
возникающих в результате мутаций, составляет элементарный эволюционный
Большинство мутаций первоначально оказывает на фенотип особей
неблагоприятное действие. В силу рецессивности мутантные аллели обычно
присутствуют в генофондах популяций в гетерозиготных по соответствующему
Благодаря этому достигается тройственный положительный результат: 1)
исключается непосредственное отрицательное влияние мутантного аллеля на
фенотипическое выражение признака, контролируемого данным геном; 2)
сохраняются нейтральные мутации, не имеющие приспособительной ценности в
настоящих условиях существования, но которые смогут приобрести такую
ценность в будущем; 3) накапливаются некоторые неблагоприятные мутации,
которые в гетерозиготном состоянии нередко повышают относительную
жизнеспособность организмов (эффект гетерозиса). Таким образом, создается
резерв наследственной изменчивости.
В большинстве природных популяций спаривание происходит неслучайным
образом. Во всех тех случаях, когда наличие одного или нескольких
наследуемых признаков повышает вероятность успешного оплодотворения гамет,
имеет место половой отбор. У растений и животных существует много
структурных и поведенческих механизмов, исключающих чисто случайный подбор
родительских особей. Например, цветки, у которых лепестки крупнее и нектара
больше, чем обычно, вероятно будут привлекать больше насекомых, что повысит
вероятность опыления и оплодотворения. Характер окраски насекомых, рыб и
птиц и особенности их поведения, связанные с постройкой гнезда, охраной
территории и брачными церемониями, повышают избирательность при
Таким образом, половой отбор как механизм избирательного скрещивания
обеспечивает некоторым особям более высокий репродуктивный потенциал, в
результате чего вероятность передачи генов этих особей следующему поколению
повышается. Репродуктивный потенциал особей с менее благоприятными
признаками понижен, и передача их аллелей последующим поколениям происходит
О дрейфе генов говорят в тех случаях, когда изменение частоты генов в
популяциях бывают случайными и не зависят от естественного отбора.
Случайный дрейф генов или генетико-автоматические процессы, или эффект
Сьюэлла Райта, может служить важным механизмом эволюционных изменений в
небольших и изолированных популяциях. В небольшой популяции могут быть
представлены не все аллели, типичные для данного вида.
Генетический дрейф может возникнуть при отделении новой популяции от
родительской из-за того, что процент определенных аллелей среди
отделившихся особей будет иным, чем в исходной популяции. Например, некий
аллель встречается в популяции в 1% случаев. Допустим, что 10 животных из
этой популяции были занесены на изолированный остров и там образовали новую
популяцию. Если среди этих десяти одно было носителем данного аллеля, то
его частота в новой популяции будет составлять уже 10%. Если же не было ни
одного, то в образовавшейся популяции этот аллель будет отсутствовать.
Или же, например, если данный аллель встречался в популяции из
1000000 особей с частотой, допустим 1% (т.е. q = 0,01), то им будет
обладать 10000 особей; а в популяции состоящей из 100 особей, этот аллель
будет иметься только у одной особи, так что вероятность его случайной
утраты в малой популяции (например, в результате преждевременной гибели
особи – единственного носителя данного аллеля в популяции) будет гораздо
С дрейфом генов связано явление, известное под названием принципа
основателя. Оно состоит в том, что при отделении от родительской популяции
небольшой ее части последняя может случайно оказаться не вполне типичной по
своему аллельному составу. Некоторые аллели в ней могут отсутствовать, а
другие будут представлены с непропорционально высокой частотой. Постоянное
скрещивание внутри такой пионерной популяции приведет к созданию генофонда,
отличающегося по частотам аллелей от генофонда исходной родительской
популяции. Дрейф генов обычно снижает генетическую изменчивость в
популяции, главным образом в результате утраты тех аллелей, которые
встречаются редко. Суть дрейфа генов в том, что из-за случайных колебаний
один из аллелей может вообще выпасть из популяции и она окажется по данному
гену гомозиготной. В результате происходит гомозиготизация особей и
затухание изменчивости. Это открывает путь случайной дифференцировке
популяций вплоть до видового обособления.
Поток генов – это перемещение аллелей из одной популяции в другую в
результате скрещивания между членами этих двух популяций. Случайное
внесение новых аллелей в популяцию – реципиента и удаление их из популяции
– донора изменяет частоту аллелей в обеих популяциях и ведет к повышению
генетической изменчивости. Несмотря на то, что поток генов вносит в
популяции генетическую изменчивость, в смысле эволюционного изменения его
действие оказывается консервативным. Распространяя мутантные аллели по всем
Читайте также:
- Уровень инструментальной готовности дошкольников 6 7 лет к школьному обучению
- Раскройте содержание политики президентов дж кеннеди и л джонсона в 1960 е гг кратко
- Примерная программа дополнительного образования в школе по фгос
- Вечные темы в романе отцы и дети природа любовь искусство кратко
- Принцип работы вертикального отстойника кратко